basement membrane proteins influence brain capillary endothelial barrier function in vitro
TRANSCRIPT

Journal of NeurochemistryLippincott—Raven Publishers, Philadelphia© 1998 International Society for Neurochemistry
Basement Membrane Proteins Influence Brain CapillaryEndothelial Barrier Function In Vitro
Thomas Tilling, Dorothea Korte, Dirk Hoheisel, and Hans-Joachim Galla
Institut für Biochemie, Westfalische Wilhelms-UniversitatMllnster, Münster, Germany
Abstract:The influence of basement membrane proteinson cellular barrier properties of primary cultures of por-cine brain capillary endothelial cells grown on permeablefilter inserts has been investigated. Measurements oftranscellular electrical resistance (TEA) by impedancespectroscopy were performed with cells cultured on typeIV collagen, fibronectin, laminin, and one-to-one mixturesof these proteins. Moreover, a one-to-one combinationof type IV collagen and SPARC (secreted protein acidicand rich in cysteine) has been studied. Rat tail collagenhas been used as a reference substratum. If TERs of cellsfrom a given preparation were low (—~350Q x cm2) onthe reference substratum, type IV collagen, fibronectin,and laminin as well as one-to-one combinations of theseproteins elevated transcellular resistances significantly(2.3- to 2.9-fold) compared with rat tail collagen. TER ofcells exhibiting a high reference level (~-~1000 ~l x cm2)could, by contrast, be increased only 1.1- to 1.2-fold. Thetype IV collagen/SPARC mixturedid not elevate TER. Ourfindings suggest that type IV collagen, fibronectin, andlaminin are involved in tight junction formation betweencerebral capillaryendothelial cells. The differential effectsobserved for individual preparations probably reflectmore or less dedifferentiated states of the endothelium,in which basement membrane proteins can influence cel-lular differentiation more or less strongly. However, ourresults indicate that type IV collagen, fibronectin, andlaminin enhance the reliability and suitability of primarymicrovascular endothelial cell cultures as an in vitromodel of the blood—brain barrier. Key Words: Blood—brain barrier—Primary cell culture—Transcellular electri-cal resistance—Basement membrane proteins—Extra-cellular matrix.J. Neurochem. 71, 1151—1157 (1998).
Brain capillary endothelial cells (BCECs) are thestructural basis of the blood—brain barrier (Reese andKamovsky, 1967; Brightman and Reese, 1969), whichtightly regulates transport of hydrophilic moleculesand ions from blood to brain tissue and vice versaand thereby ensures brain homeostasis (Abbott et al.,1986). Blood—brain barrier function is due mainly totwo structural elements: First, BCECs form a continu-ous, tight junction-connected lining of the capillarywall that prevents paracellular flux of solutes (Dermiet-
zel, 1975). Second, transport proteins localized inBCECs’ apical and basolateral membranes are respon-sible for nutrient transport from blood to brain andtransport of catabolites from brain to blood (Goldsteinand Betz, 1986). Similar to tight epithelia, BCECsshow a polar distribution of enzymes and transportproteins between the apical (luminal) and the basolat-eral (abluminal) membrane (Betz and Goldstein, 1978;Betz et a!., 1980).
Various approaches have been made to create a cellculture model of the blood—brain barrier (for review,see Joó, 1992; Laterra and Goldstein, 1993). Most ofthem are based on primary culture of BCECs, althoughimmortalized cell lines of BCECs have also been used(Roux et al., 1994; Muruganandam et al., 1997). Oneof the major problems of in vitro studies is the loss ofblood—brain barrier properties such as low ion perme-ability and the activity of certain enzymes in culture(Mischeck et al., 1989; Laterra and Goldstein, 1993).Based on the observation that astrocytes can inducetight junction formation in endothelial cells (Janzerand Raff, 1987; Tao-Cheng et al., 1987), both co-cul-tures withastrocytes (e.g., Dehouck et a!., 1990; Meyeret al., 1991) and astroglia-conditioned media (e.g., Ru-bin et al., 1991; Wolburg et a!., 1994) have been ap-plied to induce barrier properties in cultured BCECs.One aspect rarely recognized until now is the questionof whether the basement membrane located betweenBCECs and astrocytes in vivo may be involved in thedifferentiation of brain capillary endothelium leadingto the formation of a functional barrier. Basementmembranes are thin (~—~30—60nm thick) sheets con-sisting of specialized extracellular matrix proteins andproteoglycans, which can be found wherever cells(other than connective tissue cells) meet connective
Received December 15, 1997; revised manuscript received March31, 1998; accepted March 31, 1998.
Address correspondence and reprint requests to Dr. H.-J. Galla atInstitut für Biochemie, Westfblische Wilhelms-Universitdt Munster,Wilhelm-Klemm-Str. 2, D-48 149 MUnster, Germany.
Abbreviations used. BCEC, brain capillary endothelial cell;PBCEC, porcine brain capillary endothelial cell; SPARC, secretedprotein acidic and rich in cysteine; TER, transcellular electrical resis-tance.
1151

1152 T. TILLING ET AL.
tissue (Farquhar, 1991). They also surround capillaryendothelium in the brain (Goldstein and Betz, 1986).A participation of basement membrane proteins in dif-ferentiation of adjacent cells has been observed inmany tissues (for review, see Timpl, 1989; Paulsson,1992). Arthur et al. (1987) have reported that tightjunction formation in cultured BCECs induced byastrocyte-conditioned medium required an endothe-hum-derived extracellularmatrix as a growth substrate.However, they did notanalyze the influence of individ-ual extracellular matrix components of basement mem-brane on barrier properties.
In our study we have focused on the influence oftype IV co!!agen, fibronectin, laminin, and SPARC (se-creted protein acidic and rich in cysteine), four majorconstituents of basement membranes (Timpl, 1989;Yamada, 1991), on barrier function of porcine braincapillary endothelial cells (PBCECs) in primary cul-ture. Type IV collagen provides the scaffold of mostbasement membranes but also possesses cell-bindingsites (KUhn, 1994). Fibronectin is a major adhesiveglycoprotein involved incell interaction withcollagensand plays an important role in embryonic developmentand wound healing (Yamada, 1991). Laminin, whichalso possesses adhesive properties, is required for epi-thelial cell polarization (Klein et a!., 1988). In contrastto the three proteins mentioned before, SPARC, alsotermed BM-40 or osteonectin, is predominantly antiad-hesive (for review, see Lane and Sage, 1994). As it isexpressed by astrocytes both in vivo (Mendis andBrown, 1994; Mendis et a!., 1995) and in vitro (We-bersinke et al., 1992), a participation in cerebral endo-the!ial cell differentiation has been suggested (We-bersinke et al., 1992). Transce!!ular electrical resis-tance (TER), a generally accepted parameter toquantify the barrier function of a given endothelia! orepithelia! layer (Diamond, 1977; Claude, 1978), hasbeen used to determine barrier properties of PBCECmonolayers grown on extracellular matrix-coated per-meable filter inserts.
MATERIALS AND METHODS
MaterialsAll cellculture media were obtained from Biochrom (Ber-
un, Germany) except for Dulbecco’s modified Eagle’s me-diumfHam’s F-12, which was purchased from Sigma (Dei-senhofen, Germany). Biochrom was also the supplier of L-
glutamine, antibiotics, trypsin, and collagen G. Ox serumwas obtainedfrom PAA (Linz, Austria); collagenase/dispaseII (Achromobacter iophagus/Bacillus polymyxa) and humanplasma fibronectin from Boehringer Mannheim (Mannheim,Germany), human placental type IV collagen from Fluka(Neu-Ulm, Germany), and EHS laminin from Sigma (Deisen-hofen, Germany). Recombinant human SPARC (BM-40)was generously providedby Dr. Rupert Timpl (Max-Planck-Institut für Biochemie, Martinsried, Germany) and by Dr.Patrik Maurer (Institut für Biochemie, Universität zu Köln,Cologne, Germany). All other chemicals were from Sigma.
Cell culture flasks were obtained from Nunc (Roskilde,
J. Neurochem., Vol. 71, No. 3, 1998
Denmark). Twelve-well plates and Transwell filter insertswere from Costar (Cambridge, MA, U.S.A.).
Cell culturePBCECs were prepared and cultured according to Bow-
man et al. (1983) as modified by Tewes et a!. (1997). Inbrief, the cerebra of freshly slaughtered adult pigs were freedfrom the meninges and homogenized mechanically. Thebrain homogenatewas digested enzymatically using 1% (wtlvol) dispase II from B. polymyxa in PBCEC preparationmedium (Medium 199 Earle supplemented with 0.7 mM L-
glutamine, 100 ~sg/mlgentamicin, 100 U/ml penicillin, and100 ~eg/mlstreptomycin). After an incubation of 2 h at 37°C,brain capillaries were separated from myelin and cell debrisby dextran density centrifugation. The capillary pellet wastriturated using a glass pipet. A second enzymatic digestionwith 0.1% (wt/vo!) collagenase/dispase II in PBCEC platingmedium [PBCEC preparation medium with 10% (vol/vol)ox serum] for 45 mm at 37°Cserved to remove the capillarybasement membrane. Released endothelial cells were furtherpurified using a discontinuous Percoll density gradient andwere sown on collagen G-coatedculture flasks. Twenty-fourhours after initial plating, cells were washed with phosphate-buffered saline containing 1 mM Ca2~and 0.5 mM Mg2~and supplied with fresh PBCEC culture medium (PBCECplating medium without gentamicin).
To reduce contaminations by pericytes andsmooth musclecells, primary cultures of PBCECs were subcu!tivated onday 3 in vitro by gentle trypsination at room temperature.Cells were sown at 2 X !0~cells/cm2 on Transwel! cellculture inserts (12-mm diameter, 0.4-~.empore size) coatedwith various extracellular matrix proteins (see below).PBCECs were characterized by their expression of factorVIII-re!ated antigen (Barnes and Scott, 1983) as describedby Beuckmann et a!. (1995).
Coating of filter insertsTranswell cell culture inserts (12-mm diameter, 0.4-/2m
pore size) were coated by adding 100 ,u! of the respectiveextracellular matrix protein solution and allowing it to dryinside a sterile bench for —3 h. Type IV collagen was dis-solved in 50 mM acetic acid and fibronectin and lamininwere dissolved in redistilled H
20, each of them at aconcen-tration of 50 ~.tg/ml.Rat tail collagen was prepared accordingto Bornstein (1958) as a 900-~.tg/mlsolution in 0.1% (volIvo!) acetic acid, which was diluted to 50 ~sg/m! proteinshortly before use. SPARC (BM-40) stock solution con-tained 800 ~ig/ml protein in 50 mM Tris-HC1 (pH 7.4). Forone-to-one mixtures of extracellular matrix proteins, equalvolumes of 50 ~sg/mlsolutions of both proteins were mingledgently but thoroughly.
Electrical resistance measurementsPBCECs subcultured on permeable cell culture inserts in
12-well plates as described above were grown in PBCECplating medium first. Thirty-six hours after passage, mediumwas removed and replaced by measuring medium (Dulbec-co’s modified Eagle’s mediumlHam’ s F- 12 supplementedwith 6.5 mM L-glutamine, 100 /2g/ml gentamicin, 100 U/mlpenicillin, 100 ,ag/ml streptomycin, 550 nM hydrocortiso!,865 nM insulin, 30 nM sodium selenite, and 2 nM epidermalgrowth factor). The upper chamber was filled with 600 ~ilof measuring medium, the lower chamber with 1 ml.
TER was determined using a self-developed continuous-wave impedance spectrometer (Wegener et a!., 1996). For
a
55
a
a

BASEMENT MEMBRANE AND BLOOD-BRAIN BARRIER 1153
TER measurements, PBCECs on cell culture inserts weretransferred to a measuring chamber (2.5-cm diameter) filledwith 2.4 ml of measuring medium. The chamber’s groundwas covered with a thin gold film (50-nm thickness) servingas an electrode. A second ring-shaped electrode made ofplatinum dipped into the fluid above the filter insert. Imped-ance analysis wascarried out at 37°Cin the frequency rangefrom 10 to io~s~1.In general, it took 2 mm to measure theimpedance spectrum of one filter insert. Measurements wereperformed ~—~36,60, 84, and 108 h after medium exchange.Data were evaluated by fitting (nonlinear least-squares fit)the parameters of an appropriate equiva!ent circuit to theexperimental data, as described by Wegener et al. (1996).This method allows determination of the cell monolayer’selectrical resistance directly. Therefore, it was not necessaryto correct the measured TER values for the resistance of theempty filter insert and the bulk electrolyte’s conductivity.
For comparison of TER measurements obtained from dif-ferent preparations or from cells on different substrata, onlythe maximal TER value of each filter insert was used. It wasgenerally reached 60—84 h after switching the medium tomeasuring medium, independent of the coating (data notshown).
RESULTS
TER of PBCEC primary cultures varies frompreparation to preparation
We have investigated the possible involvement ofextracellular matrix proteins in PBCEC tight junctionformation. In particular, the influence of basementmembrane proteins on the TER of confluent PBCECmonolayers—as a measure of the ionic permeabilityof the intercellular tight junctions — should be clari-fied. Rat tail collagen had been previously shown tobe a suitable substratum for PBCEC cultivation onfilter inserts (Tewes et al., 1997). As it consists mainlyof type I collagen, which does not occur in basementmembranes, it could be used as a reference substratum.
Comparison of TER values obtained from differentPBCEC primary cultures demonstrated that the celllayer’s electrical resistance on a rat tail collagen sub-stratum varied from ‘—~200to 1,100 ~1 X cm2 (see Fig.1). This variability of TER is correlated to a given cellpreparation. From the large number of experiments,two groups of cell monolayers became apparent: onecontaining five preparations (26 filters) exhibiting arelatively low TER (—~350Il x cm2) and a secondgroup of four preparations (19 filters) peaking at-—1,000 ~I x cm2.
Type IV collagen, fibronectin, and laminin causedifferential elevation of TER
Type IV collagen, !aminin, and fibronectin havebeen localized immunohistochemically in the adultporcine brain, where they surround microvessels (T.Tilling and S. Decker, unpublished results). For ourexperiments, we cultured PBCECs on filter insertscoated with type IV collagen, laminin, fibronectin, andone-to-one (mass ratio) mixtures of these proteins.TERs of cells on basement membrane protein substrata
FIG. 1. TER of PBCEC monolayers on rat tail collagen-coatedfilter inserts derived from nine individual preparations. N is thenumber of filter inserts per preparation. Values are given asmeans ±SD (bars).
were compared with TERs on rat tail collagen. Asmentioned above, PBCEC monolayers on rat tail colla-gen displayed resistances of either ‘—~350or ~-—1,0001~x cm2, depending on the preparation. Consequently,we decided to quantify basement membrane proteininfluence on TER in two separate evaluations: one forPBCEC cultures exhibiting low TER on rat tail colla-gen and another for PBCECs, whose TERs on rat tailcollagen were already high.
Type IV collagen, fibronectin, and !aminin as we!! asone-to-one mixtures of them elevated TER of PBCECscompared with the reference substratum rat tail colla-gen (Fig. 2). This effect was, however, correlated tothe TER level on rat tail collagen. If the resistance onrat tail collagen was low, i.e., —~350Il x cm2, a 2.3-fold (laminin) to 2.9-fold (fibronectinllaminin) increasewas observed (Fig. 2A). In contrast, TER of PBCECreaching 1,000 ~l X cm2 on rat tail co!!agen could beonly slightly elevated by basement membrane proteins(Fig. 2B). The increase in TER (——1.1-fold) observedon fibronectin, laminin, and type IV col!agenl!amininwas not significant. A small but significant elevationof electrica! resistance could be detected on type IVcollagen, type IV collagen/fibronectin (1.1-fold), andfibronectin/laminin (1.2-fold).
SPARC exerts a negative effect on TERThe basement membrane protein SPARC has been
investigated as a possible modulator of TER inPBCECcultures. Previous experiments had shown that SPARCexerts an antiadhesive effect on PBCECs (data notshown). Thus, a one-to-one mixture of SPARC withtype IV collagen, the major component of basementmembranes, was used instead of pure SPARC for coat-ing of filter inserts. Again, cell preparations that exhib-ited low (—~350~ x cm2) or high (~—~1,000~ x cm2)resistances on rat tail collagen were used. Low-resis-tance cells exhibited only a slight increase to 550±280 1~x cm2 on type IV collagen/SPARC one-to-one mixtures. Preparations with already high TER on
J. Neurochem., Vol. 71, No. 3, 1998
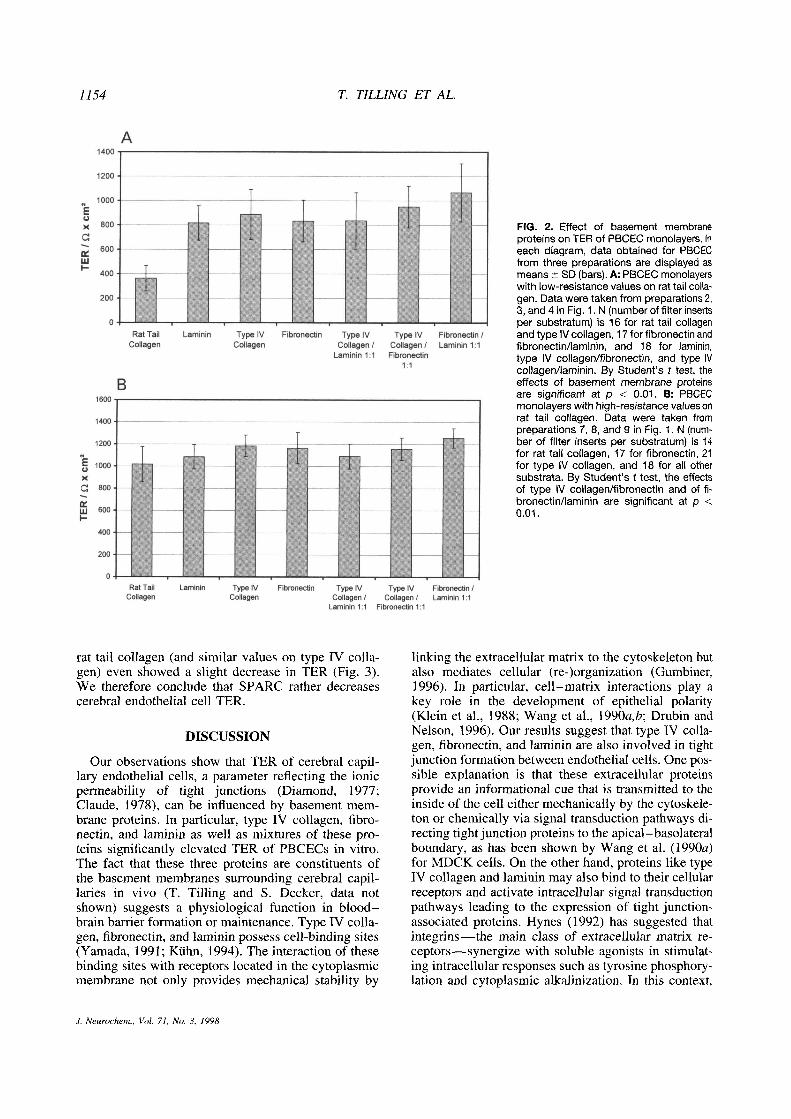
1154 T. TILLING ET AL.
FIG. 2. Effect of basement membraneproteins on TER of PBCEC monolayers. Ineach diagram, data obtained for PBCECfrom three preparations are displayed asmeans ±SD (bars). A: PBCEC monolayerswith low-resistance values on rat tail colla-gen. Data were taken from preparations 2,3, and 4 in Fig. 1. N (number offilter insertsper substratum) is 16 for rat tail collagenand type IV collagen, 17 for fibronectin andfibronectin/laminin, and 18 for laminin,type IV collagen/fibronectin, and type IVcollagen/laminin. By Student’s t test, theeffects of basement membrane proteinsare significant at p < 0.01. B: PBCECmonolayers with high-resistance values onrat tail collagen. Data were taken frompreparations 7, 8, and 9 in Fig. 1. N (num-ber of filter inserts per substratum) is 14for rat tail collagen, 17 for fibronectin, 21for type IV collagen, and 18 for all othersubstrata. By Student’s t test, the effectsof type IV collagen/fibronectin and of fi-bronectin/laminin are significant at p <
0.01.
rat tail collagen (and similar values on type IV colla-gen) even showed a slight decrease in TER (Fig. 3).We therefore conclude that SPARC rather decreasescerebral endothelial cell TER.
DISCUSSION
Our observations show that TER of cerebra! capi!-lary endothelial cells, a parameter reflecting the ionicpermeability of tight junctions (Diamond, 1977;Claude, 1978), can be influenced by basement mem-brane proteins. In particular, type IV collagen, fibro-nectin, and laminin as well as mixtures of these pro-teins significantly elevated TER of PBCECs in vitro.The fact that these three proteins are constituents ofthe basement membranes surrounding cerebral capil-laries in vivo (T. Tilling and S. Decker, data notshown) suggests a physiological function in blood—brain barrier formation or maintenance. Type IV colla-gen, fibronectin, and laminin possess cell-binding sites(Yamada, 1991; KUhn, 1994). The interaction of thesebinding sites with receptors located in the cytoplasmicmembrane not only provides mechanical stability by
linking the extracellular matrix to the cytoskeleton butalso mediates cellular (re-)organization (Gumbiner,1996). In particular, cell—matrix interactions play akey role in the development of epithelia! polarity(Klein et a!., 1988; Wang et a!., 1990a,b; Drubin andNelson, 1996). Our results suggest that type IV colla-gen, fibronectin, and laminin are also involved in tightjunction formation between endothelial cells. One pos-sible explanation is that these extracellular proteinsprovide an informational cue that is transmitted to theinside of the cell either mechanically by the cytoske!e-ton or chemically via signal transduction pathways di-recting tight junction proteins to the apical—basolateralboundary, as has been shown by Wang et al. (1990a)for MDCK cells. On the other hand, proteins like typeIV collagen and laminin may also bind to their cellularreceptors and activate intracellular signal transductionpathways leading to the expression of tight junction-associated proteins. Hynes (1992) has suggested thatintegrins —the main class of extracellular matrix re-ceptors—synergize with soluble agonists in stimulat-ing intracellular responses such as tyrosine phosphory-lation and cytop!asmic alkalinization. In this context,
a
a
as
as
as
.1. Neurochem., Vol. 71, No. 3, 1998

BASEMENT MEMBRANE AND BLOOD-BRAIN BARRIER 1155
FIG. 3. TER ofPBCECs on type IV collagen/SPARC (1:1)-coatedfilter inserts (hatched columns) compared with cells on a rattail collagen substratum (white columns) or a type IV collagensubstratum (gray columns). Cells exhibiting low resistances onrat tail collagen (‘low reference level”) are compared with thoseexhibiting high resistances (“high reference level”). On eachlevel, data acquired from three preparations are given as means±SD (bars). Low reference level: N (number of filter inserts persubstratum) is 16 for rat tail collagen and type IV collagen and18 for type IV collagen/SPARC. High reference level: N is 14 forrat tail collagen, 21 for type IV collagen, and 16 for SPARC.
it should be mentioned that hydrocortisol has beenshown to be essential for gaining PBCEC mono!ayerswith TERs up to 900 ~ X cm2 (Hoheisel et a!., 1998).Although the effects of hydrocortiso! are mediatedmainly by intracellular receptors (Evans, 1988), a syn-ergistic action withextracellu!ar matrix proteins cannotbe excluded.
In contrast to type IV collagen, fibronectin, and lam-mm, SPARC did not cause an increase in TER ofPBCECs. The TER of PBCEC on type IV collagen!SPARC-coated filters was comparable with that ofcells on the reference substrate (rat tail collagen), sug-gesting that SPARC counteracts the advantageous ef-fect of type IV collagen on endothelia! TER. AsSPARC can bind to type IV collagen (Mayer et a!.,1991), our observations may be explained by a pro-tein—protein interaction disturbing the binding of typeIV collagen to cellular receptors. Moreover, SPARChas been shown to elevate the permeability of bovinepulmonary artery endothe!ia! cells, bovine aortic endo-thelia! cells, and bovine retina! microvascular endothe-ha through F-actin-dependent changes in cell shape(Goldblum et a!., 1994), which may be due to a reduc-tion of focal contacts (Murphy-U!lrich et a!., 1995).Since during postnatal development of the mammalianbrain, SPARC can be detected in developing bloodvessels of the cerebral cortex (Mendis and Brown,1994), whereas in the adult it is located mainly inastrocytes of the hippocampus and the cerebellum(Mendis and Brown, 1994; Mendis et a!., 1995), weconclude that SPARC participates in angiogenesisrather than in blood—brain barrier formation.
Another striking result of our study was that theextent of TER elevation caused by type IV collagen,fibronectin, !aminin, and one-to-one mixtures of theseproteins was dependent on the individual PBCEC prep-aration, although we always followed the same proto-col. A pronounced increase in electrical resistance (2.3-to 2.9-fold) compared with the reference (rat tail colla-gen) could be observed only for PBCECs derived frompreparations yielding cells with low TER values on rattail collagen (—‘350 1~X cm2). If, by contrast, thereference level of TER represented by rat tail collagen-coated filters was high (‘—~1,000Il >< cm2), an elevationof TER could be achieved only by type IV collagen!fibronectin and fibronectin/laminin mixtures, both ef-fecting a rather minor but significant increase (1.1- to1.2-fold). The formation of intercellular tightjunctions,which leads to a high TER, is a main characteristicof brain microvessel endothelia! cells’ differentiatedphenotype (Crone and Olesen, 1982; Butt et a!., 1990).Differential TER levels thus reflect different degreesof PBCEC specialization. Butt et al. (1990), who inves-tigated TER in anesthetized rats from 17 days’ gesta-tion to 33 days after birth, observed values of 310±23 1~X cm2 for fetal blood vessels (17—20 days’gestation) compared with 1,215 ±110 1~X cm2 inprenatal (21 days’ gestation) and older animals. Thesharp increase in TER shortly before birth marks theonset of blood—brain barrier maturation (Butt et a!.,1990). Crone and Olesen (1982) have determined aTER of 1,870 ~ X cm2 in mature brain microvesselsof the frog. Taking these in vivo data into consider-ation, we conclude that depending on the individualcell preparation, we receive more or less dedifferenti-ated PBCECs. A cell layer with a resistance of 350 ~x cm2 is rather leaky and can be compared with fetalcerebra! endothelium. Basement membrane proteinssuch as type IV collagen, fibronectin, and lamininmight influence the barrier formation during the fo!-lowing stage of development.
If, however, cultured PBCECs form layers with analready high TER of 1,000 Il X cm2 even on rat tailcollagen, they are obviously at a more mature stage ofdifferentiation that cannot be influenced by extracellu-!ar matrix proteins to the same extent. Moreover, asTER values obtained for PBCECs cultured on fibro-nectin./!aminin in our study (1,253 ±87 1~x cm2)come relatively close to in vivo data for mature cere-bra! endothe!ia (Butt et a!., 1990), the possibility offurther elevating the electrical resistance may be gener-ally restricted.
Our observations on differential TER elevation areparticularly important for primary cultures of cerebralendothelial cells, where significant variations of cellquality can occur. It has to be pointed out that the useof basement membrane proteins as a growth substratedecreases the differences between individual prepara-tions.
Since TER is, of course, only one measure for junc-tional tightness, we are going to investigate other prop-
.1. Neurochem., Vol. 71, No. 3, 1998

1156 T. TILLING ET AL.
erties like localization of F-actin, ZO- 1, or occludin.Moreover, it will be important for future studies tocheck whether the basement membrane secreted byPBCECs in culture is dependent on the culture sur-face’s coating and whether it participates in the effectswe observed. At the present state, we are, however,able to conclude that basement membrane proteins in-crease the reliability of in vitro models of the blood—brain barrier.
Acknowledgment: We thank Bernd Tewes and JoachimWegener for helpful discussions and Christiane Engelbertzfor critically reading the manuscript. The generous gift ofrecombinant SPARC by Dr. Rupert Timpl (Max-Planck-In-stitut für Biochemie) and Dr. Patrik Maurer (Universität zuKöln) is gratefully acknowledged. This study has been sup-ported by the Deutsche Forschungsgemeinschaft, SFB 310/B 10. T.T. was a recipient of a Cusanuswerk scholarship.D.K. is a recipient of a scholarship for graduate studentsgranted by ouruniversity and the land Nordrhein-Westfalen.
REFERENCES
Abbott N. J., Bundgaard M., and Csen H. F. (1986) Comparativephysiology of the blood-brain barrier, in The Blood-Brain Bar-rier in Health and Disease (Suckling A. J., Rumsby M. G., andBradbury M. W. B., eds), pp. 52—72. Ellis Horwood, Chiches-ter.
Arthur F. E., Shivers R. R., and Bowman P. D. (1987) Induction oftight junction formation in cultured brain microvessel endothe-hal cells: local control of cell specialization. Dev. Brain Res.36, 155—159.
Barnes M. J. and Scott D. M. (1983) Glycoproteins secreted by theendothelium, in Biochemical Interactions at the Endothelium(Cryer A., ed), pp. 111—165. Elsevier, Amsterdam.
Betz A. L. and Goldstein G. W. (1978) Polarity of the blood-brainbarrier: neutral amino acid transport into isolatedbrain capillar-ies. Science 202, 225—227.
Betz A. L., Firth J. A., and Goldstein G. W. (1980) Polarity of theblood-brain barrier: distribution of enzymes between the lumi-nal and antiluminal membranes of brain capillary endnthelialcells. ~rain Res. 192, 17—28.
Beuckmann C., Hellwig S., and Galla H.-J. (1995) Induction of theblood/brain-barrier-associated enzyme alkaline phosphatase inendothelial cells from cerebral capillaries is mediated viacAMP. Eur. J. Biochem. 229, 641—644.
Bornstein M. B. (1958) Reconstituted rat-tail collagen used as sub-strate for tissue cultures on coverslips in maximow slides androller tubes. Lab. Invest. 7, 134—137.
Bowman P. D., Ennis S. R., Rarey K. E., Betz A. L., and GoldsteinG. W. (1983) Brain microvessel endothelial cells in culture: amodel for study of blood-brain barrierpermeability. Ann. Neu-rol. 14, 396—402.
Brightman M. W. and Reese T. S. (1969) Junctions between inti-mately apposed cell membranes in the vertebrate brain. J. CellBiol. 40, 648—677.
Brightman M. W. and Tao-Cheng J. H. (1993) Tight junctions ofbrain endothelium and epithelium, in The Blood-Brain Barrier(Pardridge W. M., ed), pp. 108—125. Raven Press, New York.
Butt A. M., JonesH. C., andAbbott N. J. (1990)Electrical resistanceacross the blood-brain barrier in anaesthetized rats: a develop-mental study. J. Physiol. (Lond.) 429, 47—62.
Claude P. (1978) Morphological factors influencing transepithelialpermeability: a model for the resistance of the zonulaoccludens.J. Membr. Biol. 39, 219—232.
Crone C. and Olesen S. P. (1982) Electrical resistance of brain mi-crovascular endothelium. Brain Res. 241, 49—55.
Dehouck M. P., Mdresse S., Delorme P., Fruchart J. C., and Cec-
chelli R. (1990) An easier, reproducible, and mass-productionmethod to study the blood—brain barrier in vitro. J. Neurochem.54, 1798—1801.
Dermietzel R. (1975) Junctions in the central nervous system of thecat. Cell Tissue Res. 164, 45—62.
Diamond J. M. (1977) The epithelial junction: bridge, gate and fence.Physiologist 20, 10—18.
Drubin 0. G. and Nelson W. J. (1996) Origins of cell polarity. Cell84, 335—344.
Evans R. M. (1988) The steroid and thyroid hormone receptor super-family. Science 240, 889—895.
Farquhar M. G. (1991) Theglomerular basement membrane, in CellBiology of the Extracellular Matrix (Hay E. D., ed), pp. 365-418. Plenum Press, New York.
Goldblum S. E., Ding X., Funk S. E., and Sage E. H. (1994) SPARC(secreted protein acidic and rich in cysteine) regulates endothe-hal cell shape andbarrier function. Proc. Natl. Acad. Sd. USA
91, 3448—3452.Goldstein G. W. and Betz A. L. (1986)The blood-brain barrier. Sd.
Am. 255, 70—79.Gumbiner B. (1996) Cell adhesion: the molecular basis of tissue
architecture and morphogenesis. Cell 84, 345—357.Hoheisel D., Nitz T., Franke H., Wegener J., Hakvoort A., Tilling
T., and Galla H.-J. (1998) Hydrocortisonereinforces the blood-brain barrier properties in a serum-free cell culture system.Biochem. Biophys. Res. Commun. (in press).
Hynes R. 0. (1992) Integrins: versatility, modulation, and signalingin cell adhesion. Cell 69, 11—25.
Janzer R. C. and Raff M. C. (1987) Astrocytes induce blood-brainbarrier properties in endothelial cells. Nature 325, 253—257.
Joó F. (1992) The cerebral microvessels in culture, an update. J.Neurochem. 58, 1—17.
Klein G., Langegger M., Timpl R., and Ekblom P. (1988) Role oflaminin A chain in the development of epithelial cell polarity.Cell 55, 331—341.
Kuhn K. (1994) Basement membrane (type IV) collagen. MatrixBiol. 14, 439—445.
Lane T. F. and Sage E. H. (1994) Thebiology of SPARC, a proteinthat modulates cell-matrix interactions. FASEB J. 8, 163—173.
Laterra J. and Goldstein G. W. (1993) Brainmicrovessels and micro-vascular cells in vitro, in The Blood-Brain Barrier (PardridgeW. M., ed), pp. 1—24. Raven Press, New York.
Mayer U., Aumailley M., Mann K., Timpl R., and Engel J. (1991)Calcium-dependent binding of basement membrane proteinBM-40 (osteonectin. SPARC) to basement membrane collagentype IV. Eur. J. Biochem. 198, 141—150.
Mendis D. B. and Brown I. R. (1994) Expression of the gene encod-ing the extracellular matrix glycoprotein SPARC in the devel-oping and adult mouse brain. Mol. Brain Res. 24, 11—19.
Mendis D. B., MalavalL., and Brown I. R. (1995) SPARC, an extra-cellular matrix glycoprotein containing the follistatin module,is expressed by astrocytes in synaptic enriched regions of theadult brain. Brain Res. 676, 69—79.
Meyer J., Rauh J., and Galla H.-J. (1991) The susceptibility ofcerebral endothelial cells to astrogliai induction of blood—brainbarrier enzymes depends on their proliferative state. J. Neuro-chem. 57, 1971—1977.
Mischeck U., Meyer J., and Galla H.-J. (1989) Characterization ofgamma-glutamyl transpeptidase activity of cultured endothehialcells from porcine brain capillaries. Cell Tissue Res. 256, 221—226.
Murphy-Ullrich J. E., Lane T. F., Pailero M. A., and Sage E. H.(1995) SPARC mediates focal adhesion disassembly in endo-thelial cells through a folhistatin-like region and the Ca
2~-bind-ing EF-hand. J. Cell. Biochem. 57, 341—350.
Muruganandam A., HerxL. M., Monette R., Durkin J. P., and Stani-mirovic D. B. (1997) Development of immortalized humancerebromicrovascular endothehial cell line as an in vitro modelof the human blood-brain barrier. FASEB J. 11, 1187—1197.
Paulsson M. (1992) Basement membraneproteins: structure, assem-bly, and cellular interactions. Crit. Rev. Biochem. Mol. Biol.27, 93—127.
J. Neurochem., Vol. 71, No. 3, 1998

BASEMENT MEMBRANE AND BLOOD-BRAIN BARRIER 1157
Reese T. S. and Karnovsky M. J. (1967) Fine structural localizationof a blood-brain barrier to exogenous peroxidase. J. Cell Biol.34, 207—217.
Roux F., Durieu-Trautmann 0., Chaverot N., Claire M., Mailly P.,Bourre J. M., Strosberg A. D., and Couraud P. 0. (1994) Regu-lation of gamma-glutamyl transpeptidase and alkaline phospha-tase activities in immortalized rat brain microvessel endothelialcells. J. Cell. Physiol. 159, 101—113.
Rabin L. L., Hall D. E., Porter S., Barbu K., Cannon C., HomerH. C., Janatpour M., Liaw C. W., Manning K., Morales J., Tan-ner L. I., Tomaselli K. J., and Bard F. (1991) A cell culturemodel of the blood-brain barrier. J. Cell Biol. 115, 1725—1735.
Tao-Cheng J. H., Nagy Z., and Brightman M. W. (1987) Tightjunc-tions of brain endothelium in vitro are enhanced by astroglia.J. Neurosci. 7, 3293—3299.
Tewes B., Franke H., Hellwig S., Hoheisel D., Decker S., GriescheD., Tilling T., Wegener J., and Galla H.-J. (1997) Preparationofendothehial cells in primary cultures obtained from 6-monthold pigs, in TransportAcross the Blood-Brain Barrier: In Vitroand In Viva Techniques (de Boer A. G. and Sutanto W., eds),pp. 91—97. Harwood Academic Publishers, Amsterdam.
Timpl R. (1989) Structure and biological activity of basement mem-brane proteins. Eur. J. Biochem. 180, 487—502.
Wang A. Z., Ojakian G. K., and Nelson W. J. (1990a) Steps in themorphogenesis of a polarized epithehium. I. Uncoupling theroles of cell-cell and cell-substratum contact in establishingplasma membrane polarity in multicellular epithelial (MOCK)cysts. J. Cell Sci. 95, 137—151.
Wang A. Z., Ojakian G. K., and Nelson W. J. (1990b) Steps in themorphogenesis of a polarized epithelium. II. Disassembly andassembly of plasma membrane domains during reversal of epi-thehial cell polarity in multicellular epithehial (MDCK) cysts. J.Cell Sci. 95, 153—165.
Webersinke G., Bauer H., Amberger A., Zach 0., and Bauer H. C.(1992) Comparison of gene expression of extracellular matrixmolecules in brain microvascular endothehial cells and astro-cytes. Biochem. Biophys. Res. Commun. 189, 877—884.
Wegener J., Sieber M., and Galla H.-J. (1996) Impedance analysisof epithehial and endothelial cell monolayers cultured on goldsurfaces. J. Biochem. Biophys. Methods 32, 151—170.
Wolburg H., Neuhaus J., Kniesel U., KrauB B., Schmid E.-M., Oca-lan M., Farrell C., and Risau W. (1994) Modulation of tightjunction structure in blood-brain barrier endothehial cells. J. CellSci. 107, 1347—1357.
Yamada K. M. (1991) Fibronectin and other cell interactive glyco-proteins, in Cell Biology ofExtracellular Matrix, 2nd edit. (HayE. D., ed), pp. 111—148. Plenum Press, New York.
J. Neurochem., Vol. 71, No. 3, /998