bab ii kajian pustaka 2.1 stroke iskemikiskemik serebri fokal dibagi menjadi dua yaitu iskemik...
TRANSCRIPT

12
BAB II
KAJIAN PUSTAKA
2.1 Stroke Iskemik
Penyakit serebrovaskuler termasuk stroke, menempati urutan ke dua sebagai
penyebab kematian di seluruh dunia. Di negara maju secara hitungan kasar 1 dari 20
orang dewasa menderita stroke, dan kematian setelah melewati tahun pertama setelah
menderita stroke pertama kali adalah sebanyak 20%. Beban ekonomi tidak
berpengaruh terhadap angka kematian, tetapi pada penderita yang masih hidup
sebagian besar mengalami masalah mental dan fisik. Perbandingan insiden stroke di
negara dengan sosial ekonomi baik dan di negara dengan sosial ekonomi rendah
sampai sedang menujukkan gejala yang membahayakan (Mukherjee et al., 2012).
Insiden stroke di negara maju mengalami penurunan sebanyak 42%, sedangkan
di negara dengan sosial ekonomi rendah dan sedang terjadi peningkatan sebanyak
100%. Sebanyak 80-85 % penyebab stroke yang paling sering adalah oklusi arteri,
sedangkan perdarahan intra serebral primer, perdarahan subarahnoid dan tromobisi
sinus serebri sebanyak 20-25% (Mukherjee and Patil, 2011). Di Asia stroke
hemoragik kejadiannya lebih tinggi yaitu sebanyak 50% jika dibandingkan dengan di
Eropa. Stroke terjadi bila tekanan darah terhenti karena oklusi oleh trombus atau
emboli (stroke iskemik) atau perdarahan karena pecahnya pembuluh darah (stroke
hemoragik) (Bacigaluppi et al., 2010; Mergenthaler et al., 2013)

13
2.2 Binatang Coba dengan Stroke Iskemik
Sejak tahun 1970-an model binatang telah digunakn secara luas untuk mengetahui
mekanisme terjadinya kerusakan sel otak dan percobaan terapi pre-klinik stroke.
Binatang yang umum dipakai untuk model stroke adalah primata, babi, domba, anjing,
tikus, kelinci dan tikus Mongolia (Bacigaluppi et al., 2010). Tikus sering dipakai
sebagai model stroke, karena patofisiologinya mirip dengan manusia, bisa dibuat lesi
yang sesuai dengan yang dikehendaki, prosedur relatif sederhana dan tidak infasif, biaya
lebih murah, bisa memonitor parameter fisiologi dan menganlisis jaringan otak untuk
mengetahui hasil yang diharapkan, secara genetik strainnya homogen, dan secara etik
telah diterima secara luas (Sicard et al., 2009).
Iskemik serebri fokal dibagi menjadi dua yaitu iskemik menetap dan iskemik
sementara. Iskemik menetap terjadi sebagai akibat dari iskemik yang berat, yang intinya
dikelilingi oleh daerah yang kerusakannya lebih ringan. Reperfusi dalam 3 jam tidak
memperbaiki daerah yang rusak. Iskemik sementara, mengakibatkan banyak variasi
kerusakan tergantung dari lamanya iskemik. Iskemik selama 8 menit pada tikus
meyebabkan nekrosis yang selektif, sedangkan bila iskemik lebih dari 30 menit selalu
disertai infark. Setelah iskemik, kerusakan otak yang terjadi disebabkan oleh iskemik
dan reperfusi (Bacigalupi et al., 2010, Mergenthaler et al, 2013).
Model iskemik fokal pada tikus yang sering dipakai adalah oklusi atau emboli arteri
serebri media. Metode ini sudah sering dipakai untuk menilai perkembangan
pengobatan stroke (Sicard et al; 2009; Bacigaluppi et al., 2010).
2.3 Patofisiologi Stroke
Patofisiologi terjadinya stroke sangat kompleks mulai dari mekanisme kegagalan
pembentukan energi, kehilangan homeostasis seluler, asidosis, peningkatan kadar

14
kalsium intra seluler, eksitotoksik, reactive-oxygen species mediated toxycity, aktivasi
sel neuron dan sel glia, aktifasi komplemen, terputusnya sawar darah otak dan infiltrasi
leukosit (Woodruff et al., 2011).
Otak adalah organ yang tidak bisa menyimpan cadangan energi, sehingga otak
adalah organ yang sangat rentan terhadap terhadap iskemik. Penurunan aliran darah otak
akibat suatu iskemik menyebabkan penurunan glukose dan pembentukan ATP, yang
kemudian diikuti dengan kegagalan trasnport elektron, sehingga menyebabkan kadar ion
K lebih banyak mengalir ke ekstra seluler. Akibat lebih lanjut dari kekurangan oksigen
dan glukose adalah terganggunya oksidasi di mitokondria, sehingga jumlah energi
(ATP) yang dihasilkan menurun. Penurunan jumlah ATP mengakibatkan terjadi
peningkatan metabolisme glikogen dan glukose cadangan, sehingga menyebabkan
asidosis intra seluler. Penurunan jumlah ATP mengakibatkan kegagalan pompa Na-K-
ATP ase, sehingga Na masuk ke dalam sel dan K keluar sel. Akibat selanjutnya adalah
terjadi aktivasi kanal Ca2+ dan depolarisasi neuron dan astrosit. Akibat aktivasi kanal
Ca2+, terjadi peningkatan pelepasan kadar glutamat yang berlebihan di ekstraseluler
yang kemudian akan meningkatkan jumlah Ca2+, peningkatan pembentukan radikal
bebas, mitochondria permiability trasnsisition activation, dan eksitotoksik. Mekanisme
terjadinya kematian sel pada iskemik serebri adalah karena stres oksidatif atau nitrosatif,
eksitotoksik, ketidak seimbangan ion dan apoptosis. Stres oksidatif oleh karena
eksitotoksik telah terbukti merupakan mekanisme penting mengakibatkan kematian sel
pada iskemik serebri (Niizuma K et al., 2009; Kaur et al., 2011; Breton et al., 2012)
Stroke iskemik ditandai dengan dua area iskemia yaitu; pusat inti iskemik, dan
daerah sekelilingnya yang disebut iskemik penumbra (daerah peri infark) yang terjadi
karena hipoperfusi. Luasnya daerah inti iskemik dan penumbra tergantung pada lama

15
dan beratnya okluasi pembuluh darah dan kerentanan daerah otak yang mengalami
iskemik. Yang paling sensitif terhadap iskemik adalah nukleus CA 1 neuron piramid di
hipokampus dan korteks serebri (Kumar et al., 2010).
Jaringan otak dalam keadaan fisiologis memerlukan paling sedikit 50ml darah/100
gram untuk memenuhi kebutuhan glukose dan oksigen, supaya keperluan energi (ATP)
terjamin. Bila aliran darah otak menurun kurang dari 10 ml/100 gram jaringan otak,
maka akan terbentuk inti iskemik. Daerah inti iskemik ini akan segera menjadi nekrotik
yang ireversibel dan selnya mengalami kematian (infark serebri). Bila aliran darah otak
antara 10-20 ml/100 gram jaringan otak, maka di daerah tersebut akan terbentuk daerah
iskemik yang disebut iskemik penumbra. Bila kemampuan menyediakan glukosa dan
aliran darah dari pembuluh darah kolateral tidak mencukupi maka daerah iskemik
penumbra ini akan menjadi infark, sehingga daerah infark akan bertambah (Mehta et al.,
2007; Moskowitz et al., 2010).
2.4 Mekanisme Kematian Sel pada Stroke Iskemik
2.4.1 Eksitotoksik dan iskemik
Terhentinya aliran darah ke otak selama serangan stroke, mengakibatkan terjadinya
berkurangnya oksigen dan glukose. Kekurangan oksigen dan glukose menyebabkan
otak mengalami kekurangan energi yang diperlukan untuk memelihara fungsi
homeostasis sel neuron, sehingga akan menyebabkan terjadinya depolarisasi neuron,
pelepasan neurotransmiter eksitotoksik, pengurangan ambilan kembali neurotranmiter
dari ruangan ekstraseluler. Semua proses ini mengakibatkan peningkatan Ca
ekstraseluler, yang merangsang aktivasi yang berlebihan beberapa enzim dan proses
signaling mengakibatkan kegagalan fungsi neuron yang akhirnya menyebabkan
kematian (Breton et al., 2012).

16
2.4.2 Stres oksidatif pada iskemik serebri
Stres oksidatif merupakan kunci terjadinya kelainan pada penyakit neurodegenratif
seperti penyakit Parkinson, Alzheimer, Amyitropic lateral sclerosis, multiple sclerosis
dan stroke (Allen et al., 2009). Stres oksidatif ditandai dengan terbentuknya radikal
bebas berlebihan, melebihi kapasitas antioksidan untuk mentralisasi. Antioksidan
berfungsi melindungi otak dari kerusakan yang disebabkan oleh radikal bebas. Iskemik
serebri akut menginduksi kaskade metabolik yang kompleks termasuk terbentuknya
radikal bebas seperti ROS dan reactive nitrogen species (RNS). Dalam keadaan
fisiologis ROS dan RNS terbentuk dalam jumlah kecil dan berfungsi untuk penting
untuk proses signaling dan metabolisme (Broughton et al., 2009).
Beberapa studi memperlihatkan sumber radikal bebas pada stroke iskemik adalah
stimulasi glutamat melalui mekanisme aktivasi reseptor nicotinic adenin dincleotide
phosphat dehydrogenase (NADPH), disfungsi mitokondria, aktivasi neuronal nitrix
oxide (nNOS), induksi nitrix oxid synthetase (NOS), atau cyclooxigenase (COX) auto-
oksidasi katekolamin, metabolisme asam lemak bebas, terutama yang dilepas oleh asam
arachidonat, migrasi leukosit dan neutropil yang membentuk anion superoksid dan
konversi xantin dehidrogenase ke xantin oksidase (Circu and AW, 2010, Cojocaru et al.,
2013).
Penurunan jumlah oksigen selama iskemik serebri juga menyebabkan penumpukan
asam laktat melalui metabolisme anaerob, dan mengakibatkan keadaan asidosis.
Asidosis adalah salah satu sumber ROS yang penting, karena selama asidosis H+ akan
merubah O2- menjadi H2O2 atau radikal hidroksil (0H) yang lebih reaktif (Woodruff et
al., 2011).

17
Penurunan aliran darah ke otak selama iskemik, mengakibatkan penurunan
produksi ATP. ATP diperlukan untuk semua kegiatan di dalam sel. Bila terjadi
penuruan ATP maka salah satu akibatnya adalah terjadi depolarisasi membran sel dan
pelepasan glutamat yang berlebihan ke ruang ekstra seluler yang akan mengakibatkan
eksisitotoksik. Glutamat akan berinteraksi dengan respetor N-methyl-D-aspartat
(NMDA) dan α-amino-3-hydroxy-5 methylisoxazole-4-propionic acid (AMPA) akan
mengakibatkan peningkatan Ca2+ dalam sel neuron. Selama iskemik serebri Ca2+ akan
mengaktifkan enzim proteolitik yang bertanggung jawab terhadap degradasi struktur
intra dan ekstra seluler, dan juga enszim phosfolipase A2 (PLA2) dan COX yang juga
memproduksi radikal bebas. Radikal bebas akan menyebabkan kematian sel melalui
mekanisme nekrosis atau apoptosis seperti terlihat pada Gambar 2.1 (Woodruff et al.,
2011, Jordan et al., 2012).
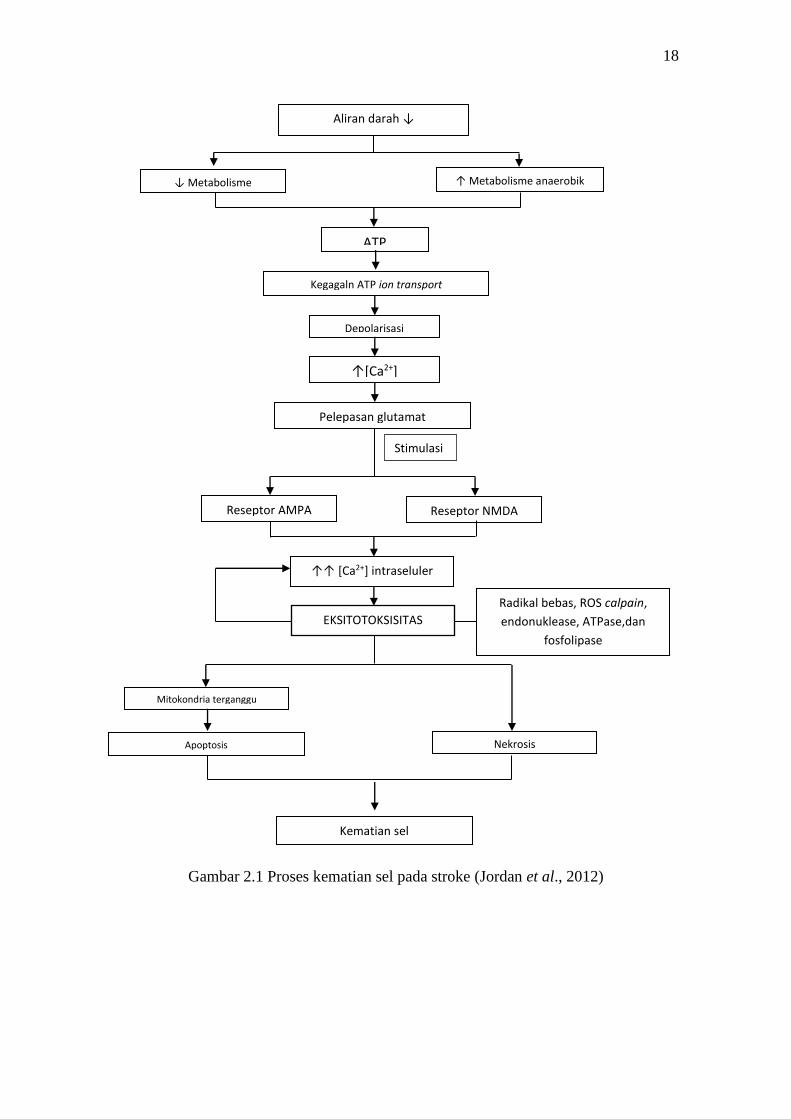
18
Gambar 2.1 Proses kematian sel pada stroke (Jordan et al., 2012)
Kematian sel
Radikal bebas, ROS calpain,
endonuklease, ATPase,dan
fosfolipase
Mitokondria terganggu
Apoptosis Nekrosis
Aliran darah ↓
↓ Metabolisme
aerobik
↑ Metabolisme anaerobik
ATP
Kegagaln ATP ion transport
Depolarisasi
↑[Ca2+]
intraseluler
Pelepasan glutamat
Stimulasi
Reseptor AMPA Reseptor NMDA
↑↑ [Ca2+] intraseluler
EKSITOTOKSISITAS

19
2.4.3 Pembentukan ROS di mitokondria
Mitokondria adalah sumber pembentukan energi bagi sel. Fungsi utama dari
mitokondria adalah pembentukan energi tinggi (ATP) melalui reaksi oksidasi-forforilasi
dengan mekanisme transport elektron, pada rantai respirasi I-IV. ROS terbentuk pada
rantai respirasi I dan III selama proses respirasi mitokondria (Niizuma et al., 2010).
ROS adalah istilah yang digunakan untuk menerangkan O2 yang membentuk radikal
bebas seperti anion superoksid (O2-), hidroksil (OH-), peroksil (RO2
-) dan alkoksil (RO-
). O2- juga membentuk spesies non radikal seperti hidrogen peroksidase (H2O2).
Mitokondria menggunakan O2 sebanyak 1-2% dan akan digunakan untuk pembentukan
ROS terutama pada kompleks I dan III. Anion superoksid yang terbentuk di
mitokondria akan dikonversi menjadi H2O2 oleh ensim superoksid dismutase yang
selanjutnya akan dikonversi menjadi H2O oleh enzim katalase. ROS terbentuk dari
molekul oksigen sebagai hasil metabolisme sel, dalam keadaan fisiologis jumlahnya
sangat sedikit (Allen and Bayraktutan, 2009, Olmez and Ozurt, 2012).
ROS dibagi menjadi 2 grup yaitu radikal bebas dan non radikal. Molekul yang
mengandung satu atau lebih elektron yang tidak berpasangan dan memberikan reaksi
terhadap molekul itu disebut radikal bebas. Bila dua radikal bebas berbagi dengan
elektron yang tidak berpasangan maka akan terbentuk non radikal.Ada tiga grup ROS
yaitu anion superoksid (O2-), radikal hidroksil (OH-) dan hidrogen peroksida (H2O2).
Anion superoksid dibentuk dengan menambahkan satu elektron ke molekul oksigen.
Proses ini diperantarai oleh nicotine adenine dincleotide phosphate oxidase (NOX),
xantin oksidase atau sistem transport elektron mitokondria. Ensim pro-oksidan seperti
NADPH Oxidase (NOX) dan xanthin oxidase mengakatalise terbentuknya O2-, yang
kemudian bereaksi dengan nitrix oxide (NO) membentuk peroxynitrite (ONOO-) yang

20
merupakan suatu radikal okisidatif sangat kuat dan akan mengakibatkan nitratisasi
(Chen et al., 2011; Breton and Rodriguez 2012).
ROS terbentuk di mitokondria oleh karena pengurangan satu elektron dari O2,
sehingga terbentuk anion superoksid (O2-). Di dalam tubuh O2
- juga dibentuk dari proses
enzimatik yaitu NADPH oksidase, cytochrome p 450 dependent oxigenase, xantin
oksidase dan peroses non ensim yaitu ketika satu elektron ditransfer langsung ke O2.
Kemudian O2- bereaksi dengan radikal bebas lainnya yaitu NO, mebentuk RNS. Anion
superoksid (O2-) kemudian dikatalisis oleh superoxid dismutase (SODs), membentuk
H2O2, yang relatif stabil yang kemudian akan dieliminasi oleh ensim antioksidan yang
ada di mitkondria maupun di sitosol. Bila H2O2 tidak dimetabolisme maka jika tersedia
ion metal dengan reaksi Fenton akan terbentuk radikal hidroksil (OH-), yan merupakan
salah satu oksidan yang bisa merusak sel (Niizuma et al., 2009, Olmez and Ozyurt,
2012) (Gambar 2.2).
ROS di mitokondria bisa terbentuk di kompleks IV (sitokrome c oksidase), tetapi
secara historis tempat pembentukan ROS pertama kali adalah kompleks I yang terletak
di lapisan dalam membran mitokondria (kompleks ubiquinone-sitokrom c reduktase.
Kompleks II yang disebut suksinat dehidrogenase (SDH) secara teoritis juga mebentuk
ROS, tetapi sampai sat ini jumlah ROS yang dibentuk tidak dapat diukur. Kompleks III
mitokondria yang disebut NADH dehidrogenase juga merupakan sumber utama ROS.
Lewat kompleks ini merupakan pintu masuk transport elektron dalam rantai respirasi
(Niizuma et al., 2010, Olmez and Ozyurt 2012).
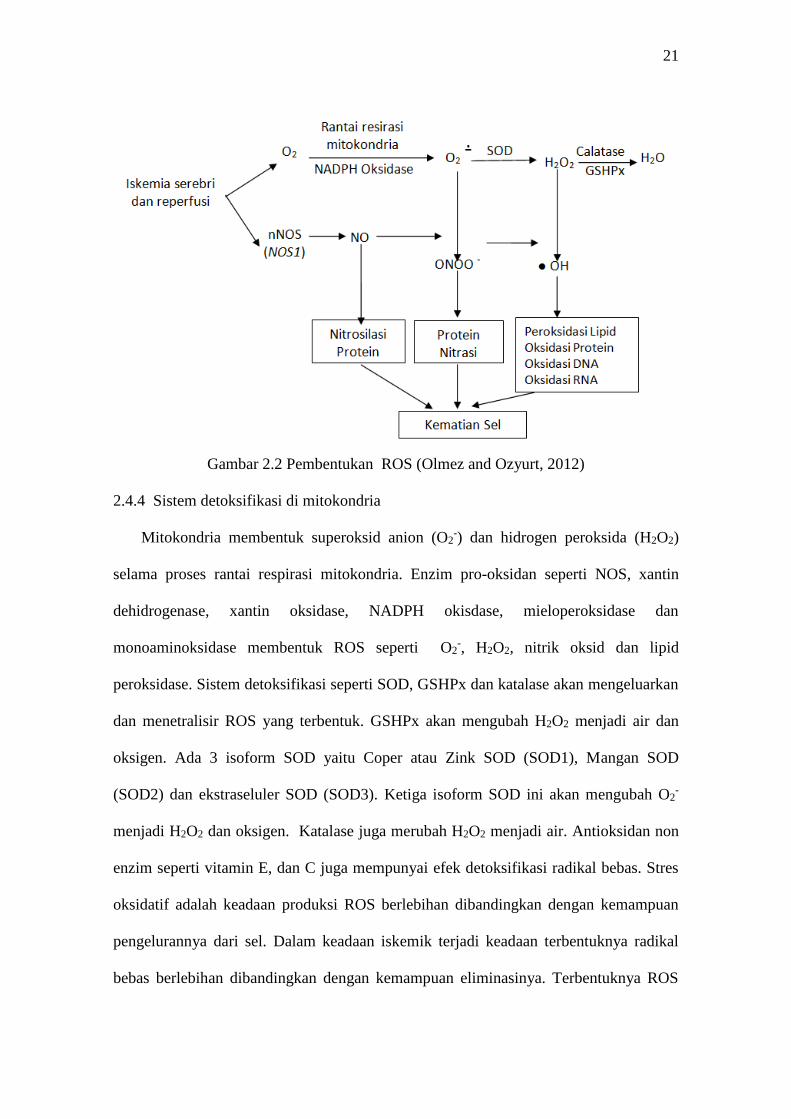
21
Gambar 2.2 Pembentukan ROS (Olmez and Ozyurt, 2012)
2.4.4 Sistem detoksifikasi di mitokondria
Mitokondria membentuk superoksid anion (O2-) dan hidrogen peroksida (H2O2)
selama proses rantai respirasi mitokondria. Enzim pro-oksidan seperti NOS, xantin
dehidrogenase, xantin oksidase, NADPH okisdase, mieloperoksidase dan
monoaminoksidase membentuk ROS seperti O2-, H2O2, nitrik oksid dan lipid
peroksidase. Sistem detoksifikasi seperti SOD, GSHPx dan katalase akan mengeluarkan
dan menetralisir ROS yang terbentuk. GSHPx akan mengubah H2O2 menjadi air dan
oksigen. Ada 3 isoform SOD yaitu Coper atau Zink SOD (SOD1), Mangan SOD
(SOD2) dan ekstraseluler SOD (SOD3). Ketiga isoform SOD ini akan mengubah O2-
menjadi H2O2 dan oksigen. Katalase juga merubah H2O2 menjadi air. Antioksidan non
enzim seperti vitamin E, dan C juga mempunyai efek detoksifikasi radikal bebas. Stres
oksidatif adalah keadaan produksi ROS berlebihan dibandingkan dengan kemampuan
pengelurannya dari sel. Dalam keadaan iskemik terjadi keadaan terbentuknya radikal
bebas berlebihan dibandingkan dengan kemampuan eliminasinya. Terbentuknya ROS

22
yang berlebihan akan mengakibatkan stres oksidatif dan merangsang sel signaling
kerusakan sel (Olmez and Ozyurt, 2012).
2.4.5 Reperfusion injury dan ROS
Reperfusion injury adalah kerusakan otak karena kembalinya aliran darah dari
pembuluh darah kolateral yang mengakibatan odem vasogenik, trasformasi hemoragik
dan meluasnya volume infark. Pada keadaan iskemik otak ROS terbentuk karena
peningkatan aktivitas ensim pro-oksidan mitokondria, dan tidak aktifnya sistem
detoksifikasi, dan kegagalan netralisasi ROS oleh antioksidan. Produkasi ROS yang
berlebihan menyebabkan kerusakan makro molekul dan aktivasi beberapa jalur kaskade
kematian sel. (Circu and Aw, 2010).
Mitokondria menjadi target utama ROS selama iskemia yang akan memulai
terjadinya kematian sel neuron, dengan mengeluarkan sitokrom c dari mitokondria.
Pelepasan sitokrom c ini dikontrol oleh protein Bcl-2 family yang terdiri atas protein
anti-apoptosis dan pro-apoptosis. ROS juga menginisiasi apotosis melalui jalur nuclear
factor kB (NF-kB) yang selanjutnya mengatifkan jalur mitogen activity protein kinase
(MAPK)/Jun kinase (JNK). Aktifasi NF-kB dapat dihambat oleh antioksidan.
Perubahan gen NF-kB, NOS, COX-2, Matrix metalloproteinase-9 (MMP-9), dan intra
celuler adhesion molecules (ICAM) yang juga terlibat dalam jalur kematian sel
(apoptosis), rusaknya sawar darah otak dan proses inflamasi (Pautz et al., 2010, (Hsieh
et al., 2010). Setelah berada di nukleus aktivasi JNK juga mengaktifkan faktor
transkripsi c-jun dan appoptosis transcription factor-2 (ATF-2) membentuk Jun-ATF-2
kompleks activator protein-1 (AP-1) yang mengatur regulasi gen proapoptosis seperti
TNF-α, Fas-L dan Bak (Fan et al., 2001).

23
2.4.6 NOS mitokondria
NOS terdiri atas neuronal NOS (nNOS), inducible NOS (iNOS) dan endotelial
NOS (eNOS). Saat ini mitokondria diketahui mempunyai NOS sendiri, yaitu
mitochondria NOS (mtNOS) yang terletak di lapisan dalam (inner membrane)
mitokondria. NOS mitokondria diproduksi di sitosol dan kemudian dipindahkan ke
mitokondria. Proses pemindahan ini belum diketahui dengan pasti. mtNOS ini selalu
aktif karena mengandung Ca+ dan terus mengkontrol respirasi mitokondria dan
merupakan molekul kunci pada reperfusion injury. Enzim yang mengaktifkan mtNOS
lebih aktif saat binatang dalam keadan hipoksia dibandingkan saat normal. mtNOS juga
dipakai sebagai penanda penuaan otak. Pada tikus yang tua mtNOS berkorelasi dengan
kemampuan untuk hidup. Selama mtNOs mengkontrol respirasi mitokondria dan
pembentukan nitriks oksida, mungkin berkorelasi dengan apoptosis setelah stroke
(Boveris et al., 2008; Finocchietto et al., 2009).
2.4.7 Peranan ROS dalam reperfusion injury.
Endotil pembuluh darah otak mempunyai fungsi unik yaitu sebagai barier yang
melindungi aliran ion secara bebas ke jaringan otak. Terganggunya barier ini saat stroke
mengakibatkan meningkatnya permiabilitas vaskuler yang akhirnya menyebakan odem
(Bektas et al,. 2010). Peningkatan reperfusi menyebabkan efek yang lebih buruk dari
pada saat iskemik oleh karena terjadi peningkatan pelepasan ROS dan akumulasi
neutropil. Keadaan ini disebut reperfusion injury. Kerusakan yang terjadi berkorelasi
dengan kadar ROS, dan peningkatan permiabilitas vaskuler juga berkorelasi dengan fase
awal dari reperfusi. Pada binatang percobaan keadaan ini dapat dikurangi dengan
menghambat produksi ROS. ROS juga menginduksi faktor lain yang bersifat sitotoksik,
yang juga berperan dalam ischemic/reperfusion injury seperti peningkatan Ca

24
intraseluler oleh karena disfungsi sarkolema retikulum dan asidosis, penurunan kadar
ATP, dan perubahan permiabiltas mitokondria (Chen et al., 2010).
NO mempunyai efek ganda yaitu bisa bermanfaat maupun merugikan. Efek
merugikan yaitu saat iskemik serebri akan menginduksi respon inflamasi dan terjadi
peningkatan pelepasan sitokin dan pembentukan iNOS, yang akan bereaksi dengan
anion superoksid membentuk peroksinitrit. Peroksinitrit dengan ROS lainnya akan
menyebabkan sel mengalami nekrosis melalui mekanisme peroksidasi lipid, kerusakan
DNA, memutus rantai respirasi mitokondria dan produksi ATP. Peroksinitrit dan ROS
lainnya juga menginduksi apoptosis melalaui peningkatan regulasi p53, yang akan
menurunkan regulasi protein Bcl-2 anti apoptopsis (Chen et al., 2010; Ola et al., 2011).
NO juga mempunyai peran menguntungkan yang diperankan oleh eNOS yaitu
melindungi sel neuron dalam keadaan iskemik sehingga tidak terjadi kerusakan sel
neuron. Efek menguntungkan ini adalah melalui mekanisme yaitu sebagai ko-faktor dari
ATP yang akan mengaktifkan kanal kalium yang mempunyai efek proteksi dan juga
melaui aktivasi posphoinisitide -3 kinae (PI-3 kinase) atau jalur Akt (Hausenloy et al.,
2006).
2.5 Apoptosis
ROS memegang peranan dalam apoptosis ketika sel mengalami stres oksidatif.
Signal apoptosis tidak secara langsung menyebabkan apoptosis, tetapi melalui regulasi
beberapa protein yang mengawali apotosis. Signal kematian sel menyebabkan apotosis
melalui jalur intrinsik maupuan ektrinsik (Azad and Iyer, 2014).
2.5.1 Jalur ekstrinsik
Apoptosis jalur ekstrinsik dimediasi oleh death receptor yang akan berikatan
dengan reseptor ligand dan akan mengaktifkan kaspase. Death receptor terdiri dari Fas

25
receptor (CD 95 atau APO-1), TNFR-1 dan TRAIL-R1 juga disebut DR4 dan TRAIL
receptor-2 (TRAIL-R2) juga disebut DR5. Death receptor kemudian berikatan dengan
ligands seperti FAS ligand, TNFα dan TRAIL. Ketiga reseptor ini berperan dalam
menginduksi apotosis dengan memberikan signal kematian sel dengan pembentukan
DISC. DISC merupakan kompleks reseptor dan ligand yang akan menginduksi pro-
kaspase 8 dan pro-kaspase 10 dan regulator lainnya sebagai ko- faktor, dan kemudian
akan mengaktifkan kaspase 3 dan 7 sebagai kaspase eksekutor (Schutze et al., 2008).
2.5.2 Jalur intrinsik
Apoptosis jalur intrinsik melibatkan mitokondria setelah menerima stimulus dari
anggota protein BH3 only seperti Bid, Bad dan Bim. Keadaan ini akan menyebabkan
oligomerisasi anggota protein pro-apptosis seperti Bax dan Bak yang terdapat
dipermukaan membran mitokondria dan akan mengakibatkan permiabilitas outer
membran mitochondria (OMM), yang akhirnya melepaskan mediator apoptosis seperti
HtrA2/Omi (mammalian serine protease) second nitochondrial activator of kaspase /
direct IAP binding protein with low pl) (Smac/diablo) sitokrom c, endonuclease G
(endo G) dan apoptosis induction factor (AIF). Sitokrom c dan APAF dan ATP/dioksi-
ATP membentuk apoptosome yang akan mengaktifkan prokaspase 9 menjadi kaspase 9,
yang selanjutnya mengaktifkan kaspase 3 sebagai eksekutor apoptosis. Peran Endo G
dan AIF adalah tidak tergantung pada kaspase, tetapi langsung menyebabkan kondensi
dan fragmentasi sel (Niizuma et al; 2009, Circu and Aw, 2010).
Inhibitor of apoptosis protein (IAP) berperan menghambat mediator apoptosis
seperti HtrA2/Omi/ Smac/diablo yang akan berikatan dengan kaspase3/7/9. Anggota
BH3 only yaitu Bid diaktifkan oleh kaspase 8 yang akan membentuk truncal Bid (tBid),
yang selanjutnya mengaktifkan Bax dan Bak. Hal ini memperlihatkan adanya hubungan
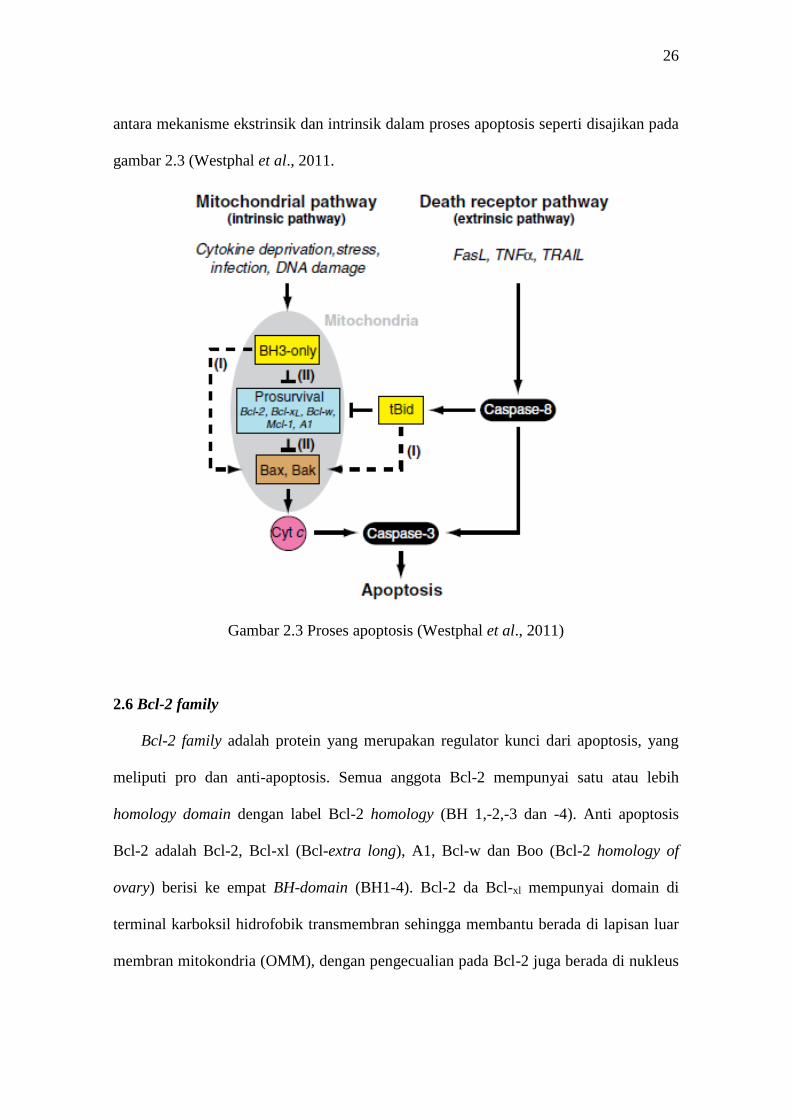
26
antara mekanisme ekstrinsik dan intrinsik dalam proses apoptosis seperti disajikan pada
gambar 2.3 (Westphal et al., 2011.
Gambar 2.3 Proses apoptosis (Westphal et al., 2011)
2.6 Bcl-2 family
Bcl-2 family adalah protein yang merupakan regulator kunci dari apoptosis, yang
meliputi pro dan anti-apoptosis. Semua anggota Bcl-2 mempunyai satu atau lebih
homology domain dengan label Bcl-2 homology (BH 1,-2,-3 dan -4). Anti apoptosis
Bcl-2 adalah Bcl-2, Bcl-xl (Bcl-extra long), A1, Bcl-w dan Boo (Bcl-2 homology of
ovary) berisi ke empat BH-domain (BH1-4). Bcl-2 da Bcl-xl mempunyai domain di
terminal karboksil hidrofobik transmembran sehingga membantu berada di lapisan luar
membran mitokondria (OMM), dengan pengecualian pada Bcl-2 juga berada di nukleus

27
dan retikulum endoplasmik dan akan bergeser ke OMM saat terjadi signal apoptosis.
Myeloid Cell leukemia factor-1 (Mcl-1) adalah anti-apoptosis yang mempunyai 3
domaian BH (BH 1-3) (Petros et al., 2004), Youle and Stresser, 2008). Bcl-2 pro-
apoptosis terdiri atas dua sub grup tergantung pada domain BH seperti Bax, Bak dan
Box, yang memiliki BH 1-3, yang hanya memiliki domain BH-3 adalah Bid, Bim dan
Bad, dengan perkecualian Bcl-xs mempunyai BH-3 dan BH-4. Ada 8 anggota BH3 only
protein yaitu harakiri (HrK), BH3 interacting domain death agonist (Bid), Bcl-2
interacting mediator cell (Bim), Bcl-2 modifying factor (Bmf), p53, promoter
upregulated modulator of apoptosis (Puma), Noxa (named for "damage"), Bcl-2
antagonist cell death (Bad) dan Bcl-2 interacting killer (Bik). Bid, Bad, dan Bim
berlokasi di sitosol dekat mitokondria. BH3 domain only juga disebut minimal death
domain yang dapat menetralisir atau menekan efek protein anti apoptosis sehingga
memungkinkan protein apoptosis seperti Bax/Bak merangsang terjadinya apoptosis
(Youle and Strsser, 2008, Ola et al.,2011) (Gambar 2,4).
Gambar 2.4 Famili Bcl-2 (Ola et al.,2011)

28
2.6.1 Peran Bcl-2 family dalam Apotosis
2.6.1.1 Bcl-2 family anti apoptosis
Setiap sel yang mempunyai nukleus paling tidak mempunyai satu anggota Bcl-2
homolog yang mengatur homeostasis pada sel dan jaringan mamalia. Ekspresi dari Bcl-
2 di organ hemopoitik mengakibatkan ekspresi sel T dan B dan sel mieloid, yang
memberi kesempatan sel progenitor untuk menghindari efek sitotoksik. Keadaan ini
terlihat jika ekspresi gen Bcl-2 pro-apoptosis pada sel yang spesifik meningkat,
disebabkan oleh rendahnya konsentrasi BCl-2 homolog anti apoptosis sehingga tidak
bisa mengimbangi Bcl-2 family proapoptosis. Bcl-2 adalah membran protein integral,
sedangkan Bcl-xl dan Bcl-w hanya berikatan secara kuat dengan membran setelah ada
signal apoptosis yang merangsang perubahan struktur untuk melindungi sel. Famili anti-
apoptosis Bcl-2 seperti Bcl-2 sendiri mencegah/menurunkan apoptosis dengan
mencegah peningkatan permiabilitas OMM, yaitu dengan menetralisir aktifitas famili
Bcl-2 pro-apotosis (BH3 only protein dan yang lainnya). Jika Bcl-2 dan Bcl-xl tidak
bisa mengaktifkan efek anti apoptosis, maka Bax mengalami oligomerisasi dan
mengambil tempat tersebut sehingga terjadi proses apoptosis (Youle and Strasser, 2008,
Brunelle et al., 2009).
2.6.1.2 Pro-apoptotik family Bcl-2
Protein anti-apoptotik seperti Bcl-2 berinterkasi dengan Bak dan Bax akan
menghambat oligomersisasnya atau berikatan dengan BH3 only untuk memblok
apoptosis. Perubahan Bak menjadi bentuk ganda Bak atau Bax mengakibatkan
fragmentasi mitokondria. Bak berhubungan Bax yang menyebabkan perubahan
permiabilitas membran luar mitokondria mengakibatkan kaskade apoptosis. Bak

29
menyebakan fragmentasi awal mitokondria, sedangkan Bax berperan dalam degenerasi
membran luar mitokondria (Brooks et al., 2007).
Dalam keadaan sehat Bak ada dalam bentuk tidak aktif di OMM, sedangkan Bax
dalam keadaan dorman di sitosol. Bila terdapat signal apoptosis maka akan merangsang
BH3 only-protein-dependent translocation dari Bax, diikuti oleh insersi ke OMM dan
terbentuknya Bak atau Bax homo-oligomer. Setelah terbentuk Bak/Bax homo-
oligomer maka akan terbentuk lubang di OMM, yang menyebabkan terjadi
permiabilitas OMM dan pelepasan isi dari intermembran space (IMS) seperti Smac
diablo, endo G dan sitokrome c ke dalam sitosol. Pelepasan protein ini akan
mengaktifkan kaskade kaspase (Wang, 2001).
2.7 Kaspase
Kaspase adalah Cystein aspartic proteases yang berkontribusi terhadap terjadinya
proses inflamasi, apoptosis dan proses vaskuler yang dominan pada stroke. Kaspase
dibedakan menurut struktur, mekanisme aktifasinya, fungsi seluler dan jalur yang
mengaktifkan apoptosis. Menurut strukturnya kaspase dibagi menjadi rantai pangjang
(casps-1, casps-2, casps-8, casps-9 dan casps-11) dan pendek (casps-3, casps-6, caps-7).
Ukuran dan komposisinya ditentukan oleh perlu tidaknya pemecahan untuk aktifitasnya
(Pop and Salvesen, 2009; Akpan and Troy 2013).
2.7.1 Mekanisme aksi kaspase
Kaspase dengan pro-domain yang panjang terdapat dalam bentuk zimogen
monomer. Aktifasi terjadi saat dimerisasi, karena perubahan struktur di inter subunit
yang lebih panjang memungkinkan bersentuhan dengan sisi katalitik yang aktif.
Pemecahan ikatan proteolitik di inter subunit diperlukan secara absolut untuk terjadinya
aktifasi (Broughton et al; 2009; Pop and Salvesen, 2009).

30
2.7.2 Fungsi seluler kaspase
Secara umum fungsi kaspase adalah: inflamasi, inisiator dan eksekutor apoptosis.
Kaspase -1 berperan dalam inflamasi pada stoke dengan mengaktifkan sitokin
proinflamasi menjadi IL-1β (Green, 2011). Struktur kaspase menentukan fungsinya
sebagai inisiator atau eksekutor. Kaspase dengan domain pendek berfungsi sebagai
eksekutor dan domain panjang sebagai inisiotor, kecuali kaspase-2 bisa beperan sebagai
eksekutor atau inisiator. Kaspase-2 sedikit berbeda meskipun mempunyai domain
panjang tetapi bentuknya adalah dimer zimogen (Parrish et al., 2013).
2.7.3 Jalur aktifasi kaspase
Kaspase berperan dalam apoptosis melalui kaskade signal kematian yaitu secara
ekstrinsik dan intrinsik. Apoptosis jalur ekstrinsik tergantung pada terbentuknya death-
inducing signaling complex (DISC), yang akan berikatan dengan ligand seperti Fas-
associated protein death dominant (FADD) yang akan mengaktifkan kaspase 8, yang
kemudian akan mengaktifkan kaspase 3 dan 7 yang bertindak sebagai eksekutor
terjadinya apoptosis. Lewat jalur ekstrinsik kaspase 8 akan memecah Bid, menjadi tBid
yang akan menyebabkan mitokondria melepaskan sitokrome c. Sitokrome c bersama
dengan apoptosis protein activator-1 (Apaf-1) dan prokaspase 9 membentuk
apoptosome. Apoptosome akan mengaktifkan pro-kaspase 3, mejandi kaspase-3, yang
akan beritndak sebagai eksekutor terjadinya apoptosis (Akpan et al., 2013, Parrish et al.,
2013).
Apoptosis jalur intrinsik di dirangsangan oleh stres oksidatif, sehingga terjadi
permiabilisasi membran luar mitokondria yang dipicu oleh famili Bcl-2 pro-apoptosis
yaitu Bax dan Bak. Dalam keadan sehat Bak berada di membran luar mitokondria
sedangkan Bax berada di sitosol. Bila terjadi signal kematian maka Bax akan bergerak

31
ke membran luar mitokondria dan akan terjadi perubahan bentuk Bax dan Bak menjadi
homo-oligomer, yang akan merubah permiabilitas membran luar mitkondria, sehingga
terjadi pelepasan sitokrome c, yang selanjutnya akan membentuk apoptosom dengan
APAF dan prokaspase 9. Prokaspase 9 berubah menjadi bentuk kaspase 9 yang akan
menstimulasi kaspase 3 sebagai eksekutor apoptosis. Mitokondria juga melepaskan
protein inhibisi yang menghambat kaspase -9, kaspase-7, kaspase 3 yaitu inhibitor of
apoptosis proteins (IAPs), terutama XIAPs. Protein lain yang dilepaskan oleh
mitokondria yaitu Smac dan Diablo yang akan memblok fungsi penghambatan dari
IAPs (Westphal et al., 2011; Niizuma et al., 2011).
2.8. Sitokrom c
Sitokrom c berperan aktif dalam biogenesis mitokondria yaitu dalam pembentukan
energi (ATP) dalam reaksi oksidasi porpolisasi. Kompleks I dari rantai respirasi yaitu
(NADH; ubiquinon oksidasereduktase) dan kompleks II (suksinat dehidrogenase) yang
berisi NADH dan FADH2, yang terbentuk dari siklus asam trikarbosilik di matrik
mitokondria. Donasi elektron dari ikatan ini melewati kompleks III (sitokrom bc1)
melalui ubiquinon. Sitokrom c menerima elektron tunggal dari kompleks III dan
membawa ke komokes IV (sitokrom c oksidase), yang merupakan batas akhir transport
elektron. Dengan menggunakan 4 elektron kompleks IV merubah oksigen menjadi air
dan merubah elektron melalui kompleks I,III dan IV ke inner membrane mitokondrial
(IMM). Pembentukan ATP (kompleks V) berasal dari masuknya proton ke matrik
mitokondria dan memerlukan membran potensial yang normal. Sitokrom c diperlukan
pada transport elektron di kompleks I dan IV. (Hosler et al., 2006; Vempati et al.,
2009)

32
Bila terjadi keadaan mitochondrial outer membrane permiabilzation (MOMP)
maka sitokrom c akan dilepaskan dari ruangan intermembran dan berikatan dengan
Apaf-1 dan dATP membentuk apoptosom, yang akan mengaktifasi prokaspase 9 dan
memicu kaskade kaspase, selanjutnya akan mengaktifkan kaspase 3 sebagai eksekutor
(Gamdo et al., 2006).
2.9. Terapi Stroke
Mortalitas dan morbiditas pasien dengan stroke masih sangat tinggi. Kesembuhan
pasien dalam 30 hari setelah stroke masih dalam tingkat sedang, hal ini terjadi sejak
tahun 1960. Sampai sat ini belum ada solusi untuk menangani kerusakan otak yang
progresif pada saat fase akut serangan stroke (Boysen et al., 2009).
2.9.1 Terapi stroke iskemik
Terapi stroke iskemik yang direkomendasikan FDA adalah rekanalisasi dengan
trombolisis yaitu dengan rTPA. Tetapi tidak banyak pasien yang mendapatkan terapi ini
karena waktu yang baik untuk pemberian rTPA adalah 3 jam setelah serangan, dan
harganya sangat mahal dan efek sampingnya terjadi perdarahan. Di Amerika Serikat
pasien yang mendapat terapi dengan rTPA adalah sebanyak 3% (Simith et al., 2008.,
Treier et al., 2011). Penelitian oleh Hacke et al., (2008), yang memberikan rtPA pada
penderita stroke dengan waktu 3-4,5 jam setelah serangan ternyata memberikan
perbaikan yang moderat yaitu perbaikan sebanyak 52,4% pada rTPA dan 45,2% pada
plasebo, tetapi insiden perdarahan meningkat. Pengobatan stroke iskemik lebin baik
dengan reperfusi seperti penggunaan rTPA ditambah dengan neuroprotektor untuk
mencegah kaskade iskemik yang terjadi selama fase akut stroke (Green et al., 2005).
Kaskade molekuler dan seluler pada ischemic-reperfusion inury pada otak adalah
sangat kompleks dan melalui bebrapa jalur. Glutamat adalah neurotransmiter utama di

33
susunan saraf pusat dan memainkan peranan utama pada sususnan saraf pusat (SSP).
Konsentasi glutamat yang berlebihan menyebabkan kerusakan neuron. Keadaan ini bisa
didapat pada penyakit neurodegeneratif, termasuk hipoksia atau iskemik serebri (Metha
et al., 2007).
Gambaran utama pada ischemia-reperfusion injury adalah terbentuknya ROS
berlebihan seperti anion superoksid (O2-) radikal hidroksil (OH-) dan hidrogen peroksida
(H2O2). Akumulasi ROS intra seluler menyebabkan oksidasi lipid, protein dan DNA
yang menyebabkan perubahan jalur signal dan menyebabkan kerusakan seluler yang
akhirnya berlanjut dengan kematian sel pada iskemik dan reperfusi serebri (Abramov et
al., 2007). Produksi ROS sangat berlebihan segera setelah iskemik, sehingga kemampun
antioksidan endogen untuk menetralisir berkurang, dan ROS yang sangat berlebihan,
akan menyebabkan kerusakan neuron. Berdasarkan mekanisme ini direkomendasikan
terapi dengan antioksidan eksogen (Margaill et al., 2005).
Mitokondria menjadi target dari ROS, yang mengalami disfungsi selama iskemik
atau reperfusi, sehingga selanjutnya akan mengalami lagi kerusakan neuron. Mekanisme
yang pasti dari kerusakan neuron sangat kompleks dan belum sepenuhnya dimengerti,
sehingga memberi peluang untuk mengembangkan terapi dan diet melawan efek
kerusakan neuron akibat iskemik. Bukti yang ada bahwa diet pitofarmaka seperti
vitamin dan bahan yang mengandung fenolik mempunyai efek proteksi terhadap neuron
saat melawan stres oksidatif, dengan menekan reaksi neuroinflamasi dan memberi
perbaikan fungsi koginitf (Cherubini et al., 2008).
Studi oleh Hariri et al., (2013) memperlihatkan penurunan kejadian stroke dan
perbaikan fungsi pada pasien yang mengkomsumsi makanan yang kaya dengan vitamin
B. Asam folat, vitamin B6 dan vitamin B12 adalah ko-faktor dalam metabolisme

34
homosistin. Hiperhomosistein diketahui sebagai salah satu faktor risiko stroke. Studi
ekspremental memperlihatkan stres oksidatif memegang peranan penting terjadinya lesi
iskemik serebral, dan mengkonsumsi anti-oksidan dengan dosis yang tinggi
dihubungkan dengan risiko yang lebih rendah terjadinya kejadian stroke pada populasi.
2.9.2. Ubiungu
Ubiungu (Ipomoea batalas L), memiliki umbi dengan warna yang berbeda yaitu ada
yang berwarana ungu, putih, merah, atau kuning. Batangnya tidak berkayu dan
mengandung air, daunnya bulat seperti jantung (Anonim, 2012). Akarnya yang disebut
dengan ubiungu banyak digunakan sebagai obat tradisional. Akar dan kulitnya banyak
mengandung polivenol seperti antosianin dan asam penolik merupakan sumber dari
vitamin A,B,C, Fe, Ca dan fosfor (Suprapta et al., 2004 ; Panda and Sonkambe, 2012).
Ubiungu ungu saat ini banyak digunakan untuk kesehatan karena di dalamnya
banyak mengandung substrat yang berguna untuk kesehatan seperti vitamin C dan
antosianin yang bersifat sebagai antioksidan. Kandungan vitamin C yang tinggi didapat
pada umbi ketela rambat, kemudian talas, ubiaung dan ketala pohon. Umbi yang
berwarna unggu seperti ketela rambat, ubiungu dan tales mengandung antosianin lebih
tinggi dibandingkan dengan umbi berwarna lainnya. Warna ungu pada ubiungu
disebabkan karena ubi tersebut banyak mengandung antosianin. Antosinin yang tinggi
pada ubiungu ini menyebabkan stabilitasnya yang lebih tinggi dibandingkan dengan
antosianin dari sumber yang lain (Suprapta et al., 2004, Motila et al., 2011). Di
Indonesia dikenal beberapa varietas ubiungu ungu yaitu MSU 01022-12, MSU 03028-
10, dan RIS03063-05. Varietas MSU 0328-10 mempunyai kadar antosianin lebih tinggi
dari ubiungu ungu dari Jepang yaitu 560 mg/100g (Jusuf et al., 2011)

35
Di Bali ditemukan varietas ubiungu unggu, diantara jenis umbi yang paling banyak
mengandung antosianin adalah ketala rambat ungu kecil kulit putih (110,51
mg/100grm) dan ketela ungu kulit putih (209,8 mg/100gram) (Suprapta et al., 2004).
Gambar 2.5 Ekstrak dan Ubiungu Varietas Bali
2.9.3. Antosianin
Antosianin adalah pigmen alami yang banyak terdapat pada buah-buahan dan
sayuran. Kandungan antosianin berbeda pada beberapa tanaman per 100 gramnya
seperti blueberries mengandung 330 mg, chokeberries 660 mg, buah kemiri 500 mg,
buncis 790 mg, coklat 1630 mg, gandum 1.900 mg, bunga anggur 3.500 mg dan kayu
manis 8.100mg (Gu et al., 2004). Ubiunggi kultivar Bali mengandung 157 mg/100 gr
(Suprapta et al., 2004). Pigmen warna merah, orange dan ungu yang menyebabkan
industri farmasi tertarik untuk menggunakan ubi ungu sebagai bahan baku industri
makanan. Antosianin yang terdapat pada tumbuhan adalah pelargonidin, sianidin,
delphinidin, petunidin dan malvidin. Antosianin juga terdapat pada kacang, sayuran dan
buah-buahan. Antosianin yang terdapat pada kedelai hitam banyak mengandung

36
cyanidine-3-glucoside (C3G) bisa melewati sawar darah otak dan mempunyai efek anti
oksidan, sehingga bisa melindungi otak dari stres oksidatif (Kang et al., 2006; Lee et
al., 2009).
2.9.4. Struktur Antosianin
Antosianin termasuk dalam kelompok plavonoid, yang biosintesisnya dari 1
molekul 4-coumaryl-CoA dan 3 molekul malonyl-CoA, yang dengan bantuan enzim
chaclone synthase (CHS), membentuk chaclone yang kemudian mengalami isomerisasi
menjadi flavanones. Flavanone akan mengalami perubahan pada 2,3-dihydro-2-
phenylchrome-4 one skleton structure dan hidrosilasi dari C-3 pada cicin C akan
berdifrensiasi menjadi bebrapa jenis flavnoid. Enzim flavanone 3-hydroxilase (F3H)
akan mengkatalase stereospesipic 3β-hydroxilation of (2S) flavanones menjadi
dihydroplavanols. Pembentukan antosianin dilanjutkan dengan mengkalase
dihydroplavanols oleh dihydroplavanols reductase (DFR) menjadi falvan-3,4-diol
(leucoanyhocyanin) yang kemudian dikonversi menjadi antosianidin oleh enzim
anthocyanidine synthetase (ANS), yang oleh enzim UDP glucoseflavonoid 3-0-glucosyl
transferase (UFGT) dikonversi menjadi antosianin (Ghmasemzadeh et al., 2012).
Seperti terlihat pada Gambar 2.5

37
Gambar 2.6 Biosintesis Antosianin (sumber Ghasemzadeh et al., 2012, dengan
modifikasi)
Antosianin diklasifikasikan dalam bermacam tipe berdasarkan substitusi pada R'3,
R'4 dan R'5 cincin B, (seperti terlihat pada tabel 2.1) dan jumlah gula yang mengalami
konyugasi, ada/tidak grup asil. Ada 6 tipe antosianin yaitu: pelargonidins, delphinidins,
peonidins, petunidins dan malvidins. Warna antosianin tergantung dari substitusi yang
terjadi pada cincin B dan saturasi warna akan meningkat dengan jumlah grup hidroksil
dan terjadi pemurunan bila terjadi penambahan grup metoksi. Bila terjadi kondisi asam
yang tinggi maka warna antosianin akan merah dan realatif stabil. Apabila keasaman
yang lemah atau pH netral maka warna antosianain akan berkurang dan menjadi tidak
stabil. Antosianin yang terdapat pada ubiungu mengandung cyanidin dan peonidin yang
lebih banyak dibandingkan dari tanaman lainnya dalam penyerapan mengalami asetilasi
sehingga menjadi lebih stabil. Kandungan cyanidin yang lebih banyak akan

38
menyebabkan warna lebih dominan berwarna ungu atau disebut tipe cyanidin
(perbandingan peo/cy < 1) dan bila kandungan peonidin yang lebih banyak maka akan
dominan berwarna merah atau disebut tipe peonidin (perbandingan peo/cy > 1) .
Perbedaan struktur kimia antosiani/antosianidin terletak pada posisi cincin 3 dan 5
seperti tampak pada Gambar 2.7 (Montilla et al; 2011; de Pascual-Tresa, 2014).
Gambar 2.7 Struktur Antosianin (Sonia de Pascual-Teresa, 2014)
Tabel 2.1 Jenis Antosianin
Anthocyanins R'3 R'4 R'5
Pelargonidin-3-glucoside
Cyanidin-3-glucoside
Peonidin-3-glucoside
Delphinin-3-glucoside
Petunidin-3-glucoside
Malvidin-3-glucoside
H
OH
OCH3
OH
OCH3
OCH3
OH
OH
OH
OH
OH
OH
H
H
H
OH
OH
OCH3
(Sumber Sonia de Pascual-Teresa, 2014)
2.9.5. Farmakokinetik Antosianin
Antosianin diabsorpsi di gaster dan intestinal dan berikatan dengan protein,
sehingga konsenstrasinya lebih tinggi dari pada plasma. Antosianin mengalami
metabolisme di hati dengan mengalami metilasi glukoronidasi, selanjutnya akan di
distribusikan ke seluruh tubuh, termasuk ke otak. Ekskresinya lewat urine (McGhie and

39
Stevenson, 2014). Antosianin ditemukan pada sel endotel otak dan parenkim otak
setelah pemberian dosis tunggal per oral pada rodensia, sehingga diduga antosianin bisa
melewati sawar darah otak (Youdin et al., 2003). Tranpotasinya ke otak sangat cepat
setelah makan dan dan dapat dijumpai dalam beberapa menit di jaringan. Antosianin
sampai ke otak lewat mekanisme mirip dengan absorsi di gaster dan saluran pencernaan
lainnya, yaitu ikatan dengan bilirubin yang memungkinkan antosianin sampai ke otak.
Ikatan antara antosianin dan bilirubun terjadi karena ikatan hidrogen yang tidak berubah
yang memungkinkan terjadinya mekanisme tranport ini (Karawjczyk et al., 2007).
Kadar antosianin yang dapat ditrasnport lewat membran basolateral berbanding terbalik
jumlah gugusan hidroksil pada cincin B, ini mengindikasikan perbedaan kadar
lipopilisiti yang membedakan selektifitas antosianin lewat sawar darah otak (Yi et al.,
2006).
Antosianin adalah suatu polivenol yang menarik perhatian untuk dikembangkan
sebagai terapi neurodegeneratif. Antosianin relatif hidrofilik, dibandingkan dengan
antioksidan lainnya tetapi antosianin bisa melewati sawar darah otak dan masuk ke
SSP. Distribusi antosianin di jaringan otak berbeda karena faktor perbedaan
pengantarannya, metabolisme dan retensi jaringan otak. Area otak yang banyak
mengandung antosianin (3-glukosid) adalah korteks, serebelum, batang otak dan
diensefalon (Milbury and Kalt, 2010)
Konsentrasi yang utuh di otak setelah pemberian per oral cukup rendah di bawah
konsentrasi yang mempunyai efek farmakologi dan neurprotktif. Secara in vivo
metabolitnya dipercaya mempunyai efek farmakologi, dan mempunyai efek lain
disamping sebagai antioksidan terhadap jaringan otak yaitu (Spencer, 2010):

40
1. Melalui interaksi dengan kaskade neuronal dan sel glia yang sangat diperlukan
untuk kehidupan sel otak terutama posfotidilinositol-3-kinase (P3IK)/Akt dan
mitogen activated-protein kinase (MAPK) famili ERK1/2, dan regulasi
prosurvival transcription factor dan ekspresi gen.
2. Kemampuannya memperbaiki aliran darah perifir dan serebral untuk
merangsang terjadinya angiogenessi dan neurogenesis di hipokampus.
3. Kemampuannya bereaksi secara langsung dengan dan agen neurorotoksis dan
pro-inflamasi yang diproduksi oleh otak yang normal maupun abnormal
2.9.6. Efek Antosianin Sebagai Antioksidan.
Efek antosianin sebagai antioksidan tergantung pada struktur dan substitusi cincin
heterocyclic dan cincin B, khususnya struktur O-di-OH pada cincin B, 2-3 double bond
dalam konyugasi dengan fungsi 4-oxo dan tambahan grup 3 dan 5-OH pada cincin
heterocyclic. Efek antioksidan sangat tergantung pada gugus hidroksil pada cincin B.
Efek antioksidan antosianin adalah melalui mekanisme: 1) Mencegah pembentukan
radikal bebas dengan enzim xanthine oksidase dan khelasi metal; 2) Mendonorkan
elektron dan menangkap radikal bebas, 3) Menghambat proses propogasi reaksi
oksidatif, 4) Menginduksi ekspresi antioksidan endogen (Montilla et al; 2010; de
Pascual-Teresa, 2014).
Antosianin menginduksi antioksidan endogen melalui aktifasi P13K/Akt, MAPK
dan Nrf2. Nrf-2 adalah faktor transkirpsi yang sangat sensitif terhadap ROS, dan
inflamsi. Antosianin akan mengaktifkan jalur Nrf2 dan dan meningkatkan ikatan dengan
antioxidant respon element (ARE ) di nukleus, sehingga menigkatkan ekspresi enszm
heme oxygenase 1 (HO-1), NADPH: quinone oxidoreductase-1 (NQO 1), glutamate

41
cystein ligase (GCL), peroxiredoxine 1 dan glutathione-S-transferase (GST) peroxidase
seperti terlihat pada Gambar 2.8 (Hwang et al., 2011; de Pascual-Teresa, 2014).
Gambar 2.8 aktivasi Nrf2 oleh antosianin. (Sumber : de Pascual-Teresa, 2014)
Penelitian Cyanindin-3-O-glucoside (CG) pada tikus dengan iskemik serebral fokal
memperoleh hasil CG dengan dosis 2mg/kg BB menurunkan volume infark sebanyak
27% yang diterapi sebelum dilakukan oklusi arteri serebri media permanen dan
menurunkan ukuran infark sebanyak 25% pada tikus yang diberikan CG dengan dosis 2
mg/kg BB 3 jam setelah oklusi arteri serebri media permanen. CG juga memblok
pengeluaran AIF dari mitokondria, tetapi tidak memblok penegeluaran sitokorm c (Min
et al., 2011).
Penelitian oleh Kang et al. (2006) mendapatkan hasil pada tikus yang dilakukan
oklusi arteri serebri media, kadar mieloperoksidase yang rendah pada otak tikus yang
diberikan CG dengan dosis 10 mg/kg BB jika dibandingkan dengan kontrol. Penelitian
oleh Giacomo et al. (2010) mendapatkan cyanidine-3-0-D-glucoside pada iskemik
/reperfusion injury pada tikus yaitu terjadi penurunan kadar lipid peroksidase, ekspresi

42
neuronal dan inducible NOS dan peningkatan kadar heme oxygenase (H0-1) dan γ -
glutamyl cystein synthetase secara bermakna antara kasus dan kontrol pada otak tikus
yang mengalami iskemik/reperfusion injury. Penelitian oleh Lu et al. ( 2010)
mendapatkan hasil PSPC menigkatkan kadar CuSOD, ZnSOD dan CAT pada oak tikus
secara bermakna (p<0,001) pada yang diberikan PSPC dibandingkan dengan kontrol
yang diinduksi dengan D-galaktose.
Percobaan pada binatang oleh Kesley et al. (2011) mendapatkan hasil bahwa
ekstrak antosianin (kalistepin dan kuromanin) mempunyai efek proteksi dari
mitochondrial oxidative stress pada culture cerebeler granule neurons (CGNs) tikus,
yaitu dengan cara menjaga ketersediaan GSH, kadar Bcl-2 pada mitokondria,
melindungi oksidasi kardiolipin dari stres oksidatif dan fragmentasi mitokondria. Studi
lain menunjukkan antosianin bisa melindungi hilangnya neuron dopaminergik yang
diinduksi dengan 6-hidroksi dopamin, melalui efek anti-oksidan dengan menekan
produksi ROS dan menjaga keseimbangan protein anti-apotosis (Bcl-2) dan protein
apoptosis (Bax), menstabilkan membran potensial mitokondria dan menghambat
aktivasi kaspase-3. Antosianin juga bisa menghambat efek 1-methyl-4-phenyl-1,2,3,6-
tetrahydroxypiridine (MPTP) yang bersifat toksik pada sel neuron dopaminergik (Kim
et al., 2010).
Hasil yang mirip diltunjukkan oleh Roghani et al., (2010), bahwa pemberian
pelargonidin yang diberikan per oral pada tikus bisa melindungi neuron dopamine dari
efek toksik, akibat 6-hidroksi dopamin, dengan menekan produksi MDA sehingga
disimpulkan antosianin merupakan efek terapi pada penyakit neurodegeneratif, seperti
penyakit Parkinson.

43
2.9.7. Efek Antosianin Terhadap Homeostasis Ion Ca2+
Kalsium memegang peranan penting untuk terjadinya eksitotoksik di sel neuron.
Antosianin memperlihatkan efek mencegah gangguan Ca2+ di intra seluler pada
penyakit neurodegenratif dan peroses penuaan. Pada percobaan oleh Shih et al. ( 2011)
dengan memberikan Aβ peptida pada beberapa jenis sel neuroblastoma terjadi
peningkatan kadar Ca2+, ketika diberikan bersamaan dengan malvidin-3-0-glucoside
pada sel neuroblastoma, maka terjadi penurunan kadar Ca2+, dibandingkan dengan
hanya pemberian Aβ peptida. Antosianin dari ekstrak ubiungu memperbaiki kerusakan
membran potensial sel yang mengakibatkan terjadinya apoptosis pada sel PCL2 yang
diinduksi dengan Aβ peptida (Ye et al., 2010)
2.9.8 Efek antosianin terhadap apoptosis.
Efek sebagai anti apoptosis, karena antosianin mempengaruhi signal protein yang
merangsang terjadinya pertumbuhan, dan mengatur jalur apoptosis yang tergantung dan
tidak tergantung dari kaspase (Reddivari et al., 2007). Peneltian oleh Lu et al., (2010)
mendapatkan jumlah sel hipokampus dan korteks serebri yang mengalami apoptosis
meningkat pada tikus yang diberikan D-galaktose, tetapi pada tikus yang diberikan D-
galaktose dan antosianin dari ekstrak ubi ungu ternyata jumlah sel hipokampus dan
korteks yang mengalami apoptosis lebih rendah secara bermakna (p<0,001). Juga terjadi
penurunan aktifasi kaspse-3 yang diperiksa dengan Western blotting pada otak tikus
yang diberikan ekstrak ubi ungu (p<0,001 Penelitian oleh Shin et al 2006
membuktikan antosianin bisa menghambat jalur apaoptosis melalui signal P53 dan
JNK pada tikus dengan iskemil serebri. Antosisnin mencegah pelepasan Aipoptosis-
inducing factor (AIF) dari mitokondria yang merupakan jalur apoptosis tidak
tergantung pada kaspase (Min et al., 2011). Penelitian oleh Kim et al. ( 2010)

44
antosianin mempunyai efek meningkatkan ekspresi protein anti-apoptosis seperti Bcl-2
dan menekan ekspresi protein pro-apoptis Bax, pada model penyakit Parkinson yang
diinduksi dengan sat toksik 6-S
2.9.9. Antosianin dan iskemik serebri
Antosianin mempunyai efek mengurangi lesi pada iskemik baik in vitro maupun in
vivo. Secara in vitro antosianin mempunyaai efek neuroprotektif. Penelitian oleh Kang
et al. (2006) membuktikan antosianin dari ekstrak buah mulbery dan cyanidine-3-0 -
glucoside mempunyai efek menurunkan lesi iskemik pada sel PC12, terutama pada sel
yang diinduksi dengan stres oksidatif dengan oxygen glucose devripation (OGD). Hasil
peneltian yang hampir sama diperoleh oleh Bhuiyan et al., (2011), yaitu pemberian
cyanidin-3-0-glucoside bisa melindungi sel neuron korteks yang diinduksi dengan
OGD dan reperfusi. Percobaan in vivo terhadap iskemik selama oklusi maupun reperfusi
pada tikus yang dilakukan penyumbatan arteri serebri media dan reperfusi, antosianin
mempunyai efek neuroprotektif karena dengan pemberian antosianin maka infark yeng
terjadi lebih ringan dibandingkan denga kontrol (Shin et al., 2006; Ritz et al., 2008a).
Studi oleh Ritz et al. (2008a) (2008b) menunnjukkan antosianin dari anggur merah
memperlihatkan efek neuroprotektif selama oklusi pada tikus karena terjadi penurunan
kadar asam amino eksitotoksik seperti glutamat, yang terjadi selama iskemik serebri.
Studi in vivo oleh Nade et al. (2010) menunjukkan bahwa antosianin memiliki efek
proteksi terhadap kerusakan karena iskemik dan berefek terhadap perbaikan fungsi
kognisi akibat iskemik pada pemberian jangka panjang. Tikus dengan oklusi arteri
serebri media dan reperfusi yang diberikan antosainin dari ekstrak akar Hibiscus rosa
sinesis mempunya kapasitas kemampuan belajar dan memori lebih baik.

45
Antosianin mempunyai beberapa efek lain seperti penurunan lipid di sel hepar,
lewat interkasi dengan peroxisome proliferator-activated receptor gamma (PPARY).
Cyanidin mempunyai efek seperti obat yang menurunkan lipid seperti lovastatin atau
fenofibrate. Mekanisme ini dijelaskan pada studi epidemiologi klinik bahwa dengan diet
tinggi antosianin mempunyai efek terhadap kekakuan arteri dan homeostasis kolesterol
(Jennings et al., 2012).
2.9.10. Efek terhadap inflamasi
Studi in vivo menunjukkan bahwa antosianin mempunyai efek anti inflamasi yang
bermakna. Seperti contoh pada tikus yang diberikan lipopolisakarida, ternyata
antosianin dari ubi ungu manis secara bermakna menekan efek lipopalisakarida untuk
mengekpresikan COX, iNOS, TNF-alpha, interleukin 1 betha (IL-1β), interleukin-6
(IL-6) pada otak tikus (Wang et al., 2010). Antosianin yang diekstrak dari ubi jalar
manis berwarna ungu mempunyai efek menekan ekspresi iNOS dan COX-2 pada liver
tikus yang diinduksi dengan dimetilnitrosamin (Hwang et al., 2011).
2.9.11 Efek neuroprotektif
Mekanisme yang telah diketahui sebelumnya yaitu efek neuroprotektif antosianin
terhadap lesi oksidatif adalah melalui induksi dari NF-E2-related factor-2(NRF-
2)/antioxidant response element (ARE). NRF-2 aktif selama stress oksidatif dan
meningkatkan transkripsi antioksidan. NRF-2 mempunayi efek neuroprotektif pada
bermacam model dari lesi neuronal (Johnson et al., 2008; Greco et al., 2010).
Mekanisme lain dari efek neuroprotektif dari antosianin adalah melalui pengaturan
beberapa jalur signaling seperti kaskade jalur exstracelluler signaling -regulated kinase
(ERK1/2), c AMP respon elemen binding protein (CREB), brain-drived neurotrophic

46
factor (BDNF), sama seperti jalur P13K/AKT. Sehingga antosianin mempunyai efek
pleotropik baik in vitro dan in vivo (Wlliam et al., 2008; Lu et al., 2010).
Penelitian antosianin yang diekstrak dari ubiungu kultivar Bali, pada mencit yang
mengalami stres oksidatif, dapat menurunkan kadar malondialdehyde (MDA), yang
merupakan suatu pertanda oksidasi lipid pada membran atau organ (Jawi et al., 2008).
Penelitian oleh Jawi et al. (2012a) memperlihatkan ekstrak ubiungu yang mengandung
antosianin pada tikus yang diberikan streptozotocin untuk menginduksi terjadinya
diabetes melitus, terdapat peningkatan total kadar antioksidan yang bermakna (p<0,05)
pada tikus yang diberikan ekstrak ubujalar ungu, serta terjadi perbedaan yang bermakna
(p<0,05) peningkatan glukose darah dan kadar MDA pada tikus yang tidak diberikan
ekstrak ubiungu. Ekstrak ubiungu kultivar Bali juga mempunyai efek antihipertensi
dan peningkatan ekspresi eNOS pada tikus yang diinduksi hipertensi (Jawi et al.,
2012b). Efek ekstrak ubiungu terhadap proteksi stres oksidatif pada endotil aorta kelinci
diteliti oleh Jawi et al. (2014). Hasil dari penelitian ini adalah terjadi peningkatan yang
bermakna (p<0,05) kadar SOD-2, NRF2, dan penurunan yang bermakna (p<0,05) kadar
vascular celluler adhesion molecule-1 (VCAM-1).
Berdasarkan efek antosianin seperti disebutkan oleh peneliti di atas yang
mempunyai efek anti inflamasi, proteksi terhadap stres oksidatif sehingga memberi
perlindungan terjadinya apoptosis, maka penulis ingin meneliti ekstrak ubiungu
terhadap apoptosis dengan melihat ekspresi protein pro atau anti-apoptosis.