atp mediates calcium signaling between astrocytes … · atp mediates calcium signaling between...
TRANSCRIPT

of August 27, 2018.This information is current as
γby IFN-Astrocytes and Microglial Cells: Modulation ATP Mediates Calcium Signaling Between
Claudia Verderio and Michela Matteoli
http://www.jimmunol.org/content/166/10/6383doi: 10.4049/jimmunol.166.10.6383
2001; 166:6383-6391; ;J Immunol
Referenceshttp://www.jimmunol.org/content/166/10/6383.full#ref-list-1
, 22 of which you can access for free at: cites 54 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

ATP Mediates Calcium Signaling Between Astrocytes andMicroglial Cells: Modulation by IFN- g1
Claudia Verderio2 and Michela Matteoli
Calcium-mediated intercellular communication is a mechanism by which astrocytes communicate with each other and modulatethe activity of adjacent cells, including neurons and oligodendrocytes. We have investigated whether microglia, the immuneeffector cells involved in several diseases of the CNS, are actively involved in this communication network. To address this issue,we analyzed calcium dynamics in fura-2-loaded cocultures of astrocytes and microglia under physiological conditions and in thepresence of the inflammatory cytokine IFN-g. The intracellular calcium increases in astrocytes, occurring spontaneously or as aresult of mechanical or bradykinin stimulation, induced the release of ATP, which, in turn, was responsible for triggering adelayed calcium response in microglial cells. Repeated stimulations of microglial cells by astrocyte-released ATP activated P2X7
purinergic receptor on microglial cells and greatly increased membrane permeability, eventually leading to microglial apoptosis.IFN-g increased ATP release and potentiated the P2X7-mediated cytolytic effect. This is the first study showing that ATP mediatesa form of calcium signaling between astrocytes and microglia. This mechanism of intercellular communication may be involvedin controlling the number and function of microglial cells under pathophysiologic CNS conditions. The Journal of Immunology,2001, 166: 6383–6391.
Resident microglia are CNS immune effector cells that par-ticipate in many different diseases (1, 2). Under patho-logical conditions they change their morphology, up-reg-
ulate a number of surface molecules, and acquire the features ofcytotoxic, phagocytic cells. Although the phenomenology of theiractivation has been well documented, the signals by which theyinteract with their environment are still elusive (2). Microenviron-ment plays a key role in CNS immunopathogenetic events, and theinteractions between microglia and lymphocytes or other immu-nocompetent cells probably play a pivotal role in the developmentof CNS inflammation (3). Identifying the key signals governing theinteractions between microglial cells and their surroundings is ofparamount importance for understanding both normal and patho-logic functioning of the nervous system.
ATP is a major factor mediating intercellular communication inthe immune and nervous systems and triggers a variety of strik-ingly different biological effects (4–7). In the brain it is consideredto be the dominant extracellular messenger for astrocyte-to-astro-cyte calcium-mediated communication. Astrocytes release ATPupon mechanical stimulation (8) or glutamatergic receptor activa-tion (9) and respond to ATP with a propagating wave of intracel-lular calcium ([Ca21]i)
3 increases (8), a process that is thought toserve as a long-range signaling system in the CNS (10, 11).
Functionally active purinergic receptors have been detected incultured and in situ microglial cells (12–17), thus suggesting thepossibility that ATP may also act in astrocyte-to-microglia com-munication. In this study we demonstrate the existence of an ATP-mediated calcium signaling mechanism between astrocytes andmicroglial cells that is increased in the presence of the inflamma-tory cytokine IFN-g and eventually leads to microglial apoptosis.
Materials and MethodsCell cultures
Hippocampal mixed glia cultures from embryonic rat pups (embryonic day18) were obtained using previously described methods (18). The cultureswere grown in MEM supplemented with 20% FCS and 5.5 g/L glucose.The astrocytic and microglial components of the cultures were determinedby means of immunostaining for glial fibrillar acidic protein (GFAP) andCSF-1R. The purified microglia were harvested by shaking 3-wk-old cul-tures, seeded on glass coverslips, and cultured in the same medium.
Fura-2 videomicroscopy
The cultures were loaded with 5mM fura-2 pentacetoxy-methylester inKrebs-Ringer solution buffered with HEPES (KRH; 150 mM NaCl, 5 mMKCl, 1.2 mM MgSO4, 2 mM CaCl2, 10 mM glucose, and 10 mM HEPES/NaOH, pH 7.4) for 1 h at 37°C, washed in the same solution to allowde-esterification of the dye, and transferred to the recording chamber of aninverted microscope (Axiovert 100; Zeiss, New York, NY) equipped witha calcium-imaging unit. A modified CAM-230 dual wavelength microflu-orometer (Jasco, Tokyo, Japan) was used as a light source for the assays.The experiments were performed at room temperature (24–25°C) using anAxon Imaging Workbench 2.2 equipped with a PCO Super VGA Sensi-Cam (Axon Instruments, Foster City, CA). A single astrocyte in the fieldwas gently mechanical stimulated with a glass microelectrode to evokeintercellular calcium waves.
ATP measurements
Bioluminescence assay.ATP levels in the extracellular saline incubated30 min with dishes containing a pure hippocampal astrocytic monolayerwere measured using a luciferin/luciferase assay (Molecular Probes, Lei-den, The Netherlands) and a luminometer (Lumat LB9501; Berthold,Nashua, NH). The experimental samples were compared with a standardATP curve created on the basis of saline samples containing known con-centrations of ATP. Each sample was run in duplicate. Most of the sampleswere assayed within 5–10 min of collection; the others were frozen forsubsequent ATP determination.
Consiglio Nazionale delle Ricerche Cellular and Molecular Pharmacology and “B.Ceccarelli” Centers, Department of Medical Pharmacology, Milan, Italy
Received for publication October 16, 2000. Accepted for publication March 7, 2001.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from Istituto Superiore di Sanita’ (Project Mul-tiple Sclerosis 61), Cofin 2000, and Azienda Spaziale Italiana (I/R/149/00).2 Address correspondence and reprint requests to Dr. Claudia Verderio, ConsiglioNazionale delle Ricerche, Cellular and Molecular Pharmacology and “B. Ceccarelli”Centers, Department of Medical Pharmacology, University of Milan, Via Vanvitelli32, 20129 Milan, Italy. E-mail address: [email protected] Abbreviations used in this paper: [Ca21]i, intracellular calcium; KRH, Krebs-Ringersolution with HEPES; CSF-1R, CSF receptor; PPADS, pyridoxalphosphate-6-azo-phenyl-2,4-disulfonic acid, tetrasodium; oATP, oxidized ATP; MS, multiple sclero-sis; GFAP, glial fibrillar acidic protein.
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Extracellular ATP bioassay.One milliliter of extracellular saline incu-bated for 30 min with dishes containing a pure hippocampal astrocyticmonolayer in the presence or in the absence of IFN-g was split into twoaliquots before testing on fura-2-loaded astrocytes. One aliquot was pre-treated with apyrase (30 U/ml) for 15 min before testing. Five hundredmicroliters of test solution or saline containing known concentrations ofATP were applied to astrocytes as ATP sensor cells during image acqui-sition, and the amplitude of the [Ca21]i response was measured.
Immunocytochemistry
At the end of the recording session, cultures were fixed at room tempera-ture with 4% paraformaldehyde in 0.1 M phosphate buffer containing 0.12M sucrose for 25 min. Fixed cells were permeabilized with detergent andlabeled with anti-GFAP monoclonal Abs (Sigma, Milan, Italy), and anti-CSF-R1 polyclonal Abs (Santa Cruz Biotechnology, Santa Cruz, CA), fol-lowed by fluorescein-anti-mouse Abs (Jackson ImmunoResearch, WestGrove, PA) and rhodamine-conjugated anti-rabbit Abs (Jackson Immu-noResearch). The coverslips were mounted in 70% glycerol in phosphatebuffer containing 1 mg/ml phenylenediamine. The images were acquiredusing a Bio-Rad MRC-1024 confocal microscope (Bio-Rad, Hercules, CA)equipped with LaserSharp 3.2 software. The morphological features ofapoptosis were monitored by means of phase contrast microscopy and la-beling with propidium iodide in PBS on cultures fixed in methanol pre-cooled at220°C.
Chemicals
Most of the chemicals (pyridoxalphosphate-6-azophenyl-2,4-disulfonicacid, tetrasodium (PPADS), ATP, bradykinin, apyrase, and 18-a2glycyr-rhetinic acid) were obtained from Sigma. Fura-2-acetoxymethyl ester waspurchased from Calbiochem (La Jolla, CA), and periodate-oxidized ATPwas a gift from Prof. F. Di Virgilio (University of Ferrara, Ferrara, Italy).
Results[Ca21] i increases in astrocytes induce delayed [Ca21] i
responses in microglial cells
Primary cultures of embryonic hippocampal glial cells predomi-nantly contain astrocytes, with a variable percentage of microglialcells. Microglial cells are characterized by their amoeboid shapeand can be unambiguously distinguished from astrocytes becausethey are stained by Abs against the CSF receptor (CSF-1R; Fig. 1,A andB, red), but not by Abs against the GFAP cytoskeletal glialprotein (green). The mixed cultures were loaded with the cell-permeant fura-2-acetoxymethyl ester fluorescent calcium dye andimaged using digital imaging techniques in a static bath. Gentlemechanical contact between a patch pipette and the surface of asingle astrocyte (pseudocolor images in Fig. 1C, arrow) induced apropagating calcium wave in neighboring cells (Fig. 1C) that wereidentified as astrocytes or microglia on the basis of morphologicalcriteria or CSF-1R and GFAP immunostaining at the end of therecording session (Fig. 1C,inset). Analysis of the kinetics of[Ca21]i changes in the stimulated astrocytes and in two adjacentmicroglia cells revealed a transient, delayed [Ca21]i response inthe microglial cells (Fig. 1D).
To confirm that the delayed [Ca21]i response in microglia wasa consequence of increased [Ca21]i in astrocytes, cocultures werestimulated with 1mM bradykinin, a widely used stimulus for se-lectively evoking increased [Ca21]i in astrocytes (19). Bradykinindid not cause any [Ca21]i changes in the purified microglial cellsshaken from mixed cultures and plated onto glass coverslips (Fig.1E, bottom panel), but a large increase in [Ca21]i was induced inthe majority of cocultured astrocytes and microglial cells (Fig. 1E,top panel). Whereas bradykinin application induced a prompt[Ca21]i response in astrocytes, [Ca21]i increases occurred severalseconds later in microglia (506 20 s; mean6 SE; n 5 12).Efficient propagation of calcium signaling between astrocytes andmicroglia was also observed in cultures characterized by sponta-neous [Ca21]i oscillations (Fig. 1F), a phenomenon that may de-pend on cycles of calcium release and uptake from intracellularstores (10, 11, 20, 21). Analysis of several fields containing both
astrocytes and microglial cells (n5 31) revealed that spontaneousastrocytic [Ca21]i oscillations were coupled to a delayed responsein adjacent microglial cells (Fig. 1F).
Delayed [Ca21] i responses in microglial cells are mediated byATP
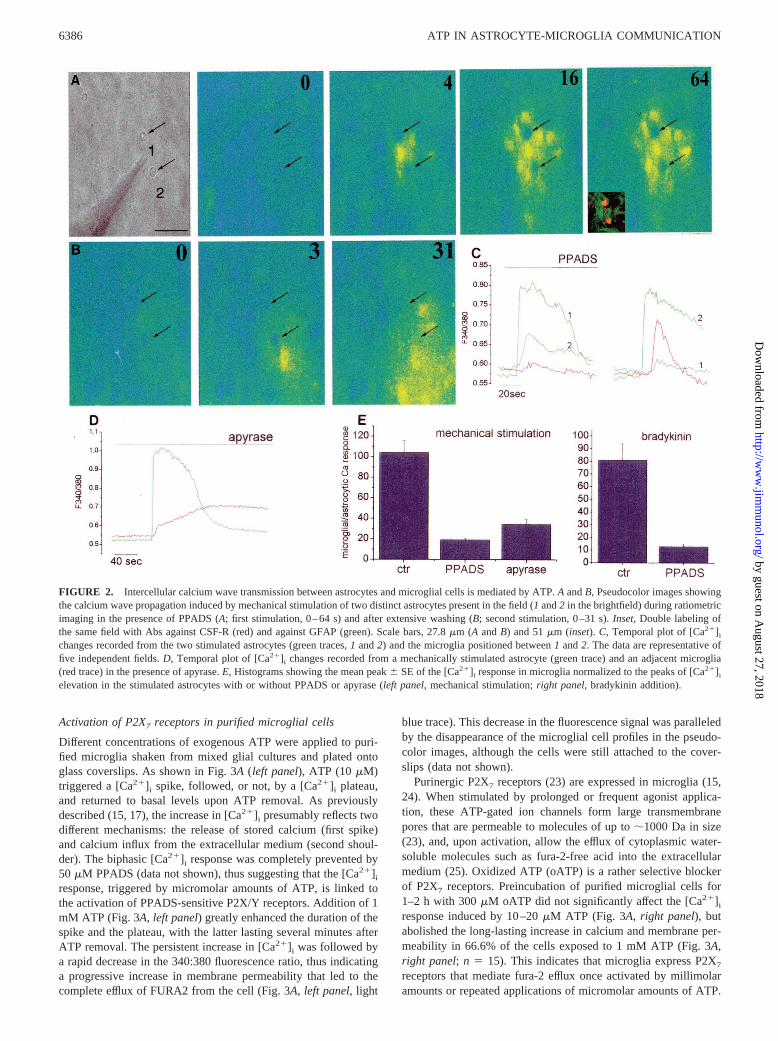
The delayed response of microglia triggered by spontaneous orevoked [Ca21]i increases in astrocytes is consistent with a second-ary response to a bioactive compound released by astrocytes upon[Ca21]i elevations. Glutamate and ATP have recently been iden-tified as diffusable factors, capable of inducing calcium wavesamong astrocytes (8, 22). As microglial cells from hippocampuslack calcium-permeable glutamate receptors (our unpublished ob-servation) and express purinergic receptors permeable to calciumions (15), we investigated whether ATP might be involved in in-tercellular calcium signaling between astrocytes and microglialcells. We found different lines of evidence indicating that this wasthe case. First, ATP was detected in the extracellular medium ofbradykinin-stimulated and spontaneously oscillating astrocytes bymeans of the sensitive luciferase bioluminescence assay (notshown) and was also detected in the medium collected from me-chanically stimulated astrocytes (8). Second, samples of extracel-lular medium taken from spontaneously oscillating astrocytes andgently added to fura-2-loaded astrocytes as ATP-sensor cells in-duced a [Ca21]i response that was prevented by pretreatment of theconditioned medium with the ATP-degrading enzyme apyrase(Fig. 5C, right panel). Third, when the astrocytes in astrocyte-microglia cocultures were stimulated mechanically or with brady-kinin, a potent inhibition of the propagation of the [Ca21]i signalto microglial cells was recorded in the presence of either the P2X/P2Y purinergic receptor antagonist PPADS (50mM) or apyrase(Fig. 2). The series of pseudocolor images in Fig. 2A shows anexample of a mechanically induced calcium wave (Fig. 2A, patchpipette in position 1) that did not propagate to microglia in thepresence of PPADS (Fig. 2A, arrows, seeinset in image 64 forimmunocytochemical identification) despite the efficient transmis-sion of the calcium signal to adjacent astrocytes (Fig. 2A, imagesfrom 0–64 s). After extensive washing of PPADS, the calciumwave induced by mechanical stimulation of another astrocyte inthe field (Fig. 2A, patch pipette shifted to position 2) did propagateto microglia according to the diffusion of the wave front (Fig. 2B,images from 0 to 31 s). The kinetics of [Ca21]i changes in the twostimulated astrocytes (green traces) and in one microglial cell (redtrace, positioned between 1 and 2) are shown in Fig. 2C. Thecalcium signal propagation from astrocytes to microglial cells wasalso potently inhibited in the presence of the ATP-degrading en-zyme apyrase (Fig. 2D). Fig. 2Eshows the quantitative analysis ofPPADS and apyrase inhibition of the delayed response of micro-glia (81% inhibition in the presence of PPADS,n 5 8; 66.5%inhibition in the presence of apyrase, with a 420% increase in thetime to peak response,n 5 13; controls,n 5 14); a similar reduc-tion in calcium signal propagation was produced by PPADS whenincreased astrocyte [Ca21]i levels were evoked by bradykinin (Fig.2E; 83.9% inhibition in the presence of PPADS,n 5 9; controls,n 5 12). Efficient propagation of the calcium signal between as-trocytes and microglia was detected in the presence of a cocktail ofglutamate receptor antagonists (100mM 2-amino-5-phosphonova-leric acid, 20mM 6-cyano-7-nitroquinoxanine-2,3-dione, and 1mM (S)-a-methyl-carboxyphenylglycine) or after 5- to 10-min in-cubation in the presence of the gap junction blocker 18a-glycyr-rhetinic acid (10mM; data not shown). These results indicate that[Ca21]i increases in astrocytes induce the release of ATP, which,in turn, activates purinergic receptors in microglial cells.
6384 ATP IN ASTROCYTE-MICROGLIA COMMUNICATION
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
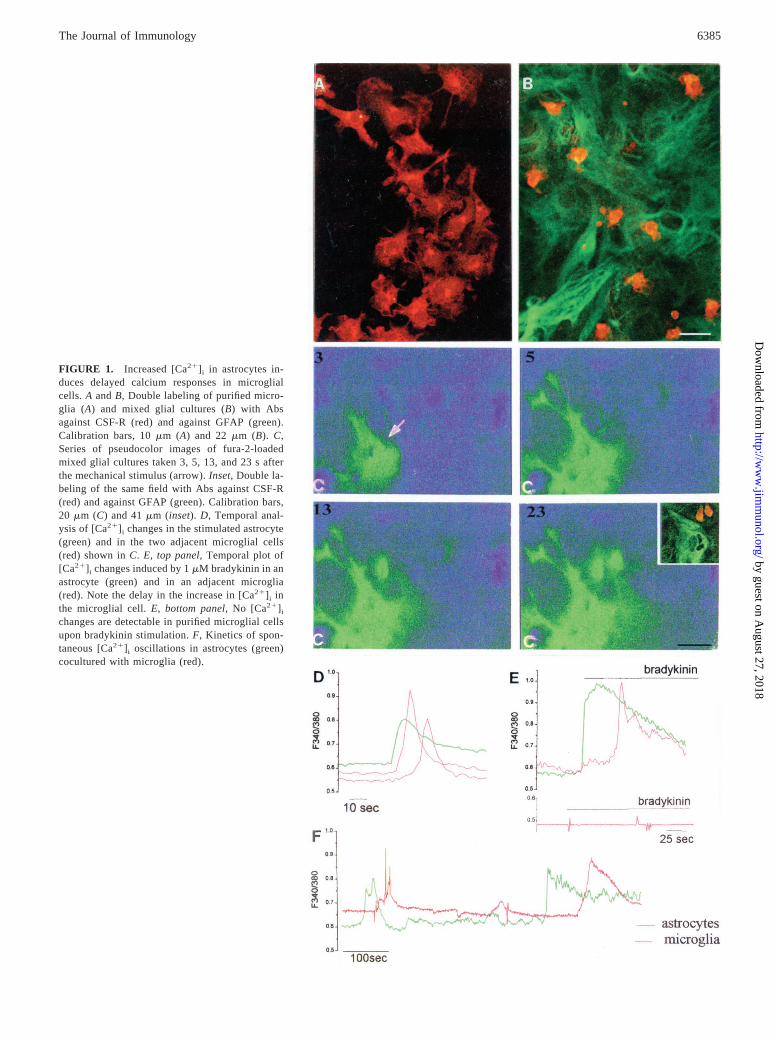
FIGURE 1. Increased [Ca21] i in astrocytes in-duces delayed calcium responses in microglialcells. A andB, Double labeling of purified micro-glia (A) and mixed glial cultures (B) with Absagainst CSF-R (red) and against GFAP (green).Calibration bars, 10mm (A) and 22mm (B). C,Series of pseudocolor images of fura-2-loadedmixed glial cultures taken 3, 5, 13, and 23 s afterthe mechanical stimulus (arrow).Inset, Double la-beling of the same field with Abs against CSF-R(red) and against GFAP (green). Calibration bars,20 mm (C) and 41mm (inset).D, Temporal anal-ysis of [Ca21] i changes in the stimulated astrocyte(green) and in the two adjacent microglial cells(red) shown inC. E, top panel, Temporal plot of[Ca21] i changes induced by 1mM bradykinin in anastrocyte (green) and in an adjacent microglia(red). Note the delay in the increase in [Ca21] i inthe microglial cell.E, bottom panel, No [Ca21] i
changes are detectable in purified microglial cellsupon bradykinin stimulation.F, Kinetics of spon-taneous [Ca21] i oscillations in astrocytes (green)cocultured with microglia (red).
6385The Journal of Immunology
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Activation of P2X7 receptors in purified microglial cells
Different concentrations of exogenous ATP were applied to puri-fied microglia shaken from mixed glial cultures and plated ontoglass coverslips. As shown in Fig. 3A(left panel), ATP (10mM)triggered a [Ca21]i spike, followed, or not, by a [Ca21]i plateau,and returned to basal levels upon ATP removal. As previouslydescribed (15, 17), the increase in [Ca21]i presumably reflects twodifferent mechanisms: the release of stored calcium (first spike)and calcium influx from the extracellular medium (second shoul-der). The biphasic [Ca21]i response was completely prevented by50 mM PPADS (data not shown), thus suggesting that the [Ca21]iresponse, triggered by micromolar amounts of ATP, is linked tothe activation of PPADS-sensitive P2X/Y receptors. Addition of 1mM ATP (Fig. 3A,left panel) greatly enhanced the duration of thespike and the plateau, with the latter lasting several minutes afterATP removal. The persistent increase in [Ca21]i was followed bya rapid decrease in the 340:380 fluorescence ratio, thus indicatinga progressive increase in membrane permeability that led to thecomplete efflux of FURA2 from the cell (Fig. 3A,left panel, light
blue trace). This decrease in the fluorescence signal was paralleledby the disappearance of the microglial cell profiles in the pseudo-color images, although the cells were still attached to the cover-slips (data not shown).
Purinergic P2X7 receptors (23) are expressed in microglia (15,24). When stimulated by prolonged or frequent agonist applica-tion, these ATP-gated ion channels form large transmembranepores that are permeable to molecules of up to;1000 Da in size(23), and, upon activation, allow the efflux of cytoplasmic water-soluble molecules such as fura-2-free acid into the extracellularmedium (25). Oxidized ATP (oATP) is a rather selective blockerof P2X7 receptors. Preincubation of purified microglial cells for1–2 h with 300mM oATP did not significantly affect the [Ca21]iresponse induced by 10–20mM ATP (Fig. 3A, right panel), butabolished the long-lasting increase in calcium and membrane per-meability in 66.6% of the cells exposed to 1 mM ATP (Fig. 3A,right panel;n 5 15). This indicates that microglia express P2X7
receptors that mediate fura-2 efflux once activated by millimolaramounts or repeated applications of micromolar amounts of ATP.
FIGURE 2. Intercellular calcium wave transmission between astrocytes and microglial cells is mediated by ATP.A andB, Pseudocolor images showingthe calcium wave propagation induced by mechanical stimulation of two distinct astrocytes present in the field (1 and2 in the brightfield) during ratiometricimaging in the presence of PPADS (A; first stimulation, 0–64 s) and after extensive washing (B; second stimulation, 0–31 s).Inset, Double labeling ofthe same field with Abs against CSF-R (red) and against GFAP (green). Scale bars, 27.8mm (A andB) and 51mm (inset).C, Temporal plot of [Ca21]i
changes recorded from the two stimulated astrocytes (green traces,1 and2) and the microglia positioned between1 and2. The data are representative offive independent fields.D, Temporal plot of [Ca21]i changes recorded from a mechanically stimulated astrocyte (green trace) and an adjacent microglia(red trace) in the presence of apyrase.E, Histograms showing the mean peak6 SE of the [Ca21]i response in microglia normalized to the peaks of [Ca21]i
elevation in the stimulated astrocytes with or without PPADS or apyrase (left panel, mechanical stimulation;right panel, bradykinin addition).
6386 ATP IN ASTROCYTE-MICROGLIA COMMUNICATION
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
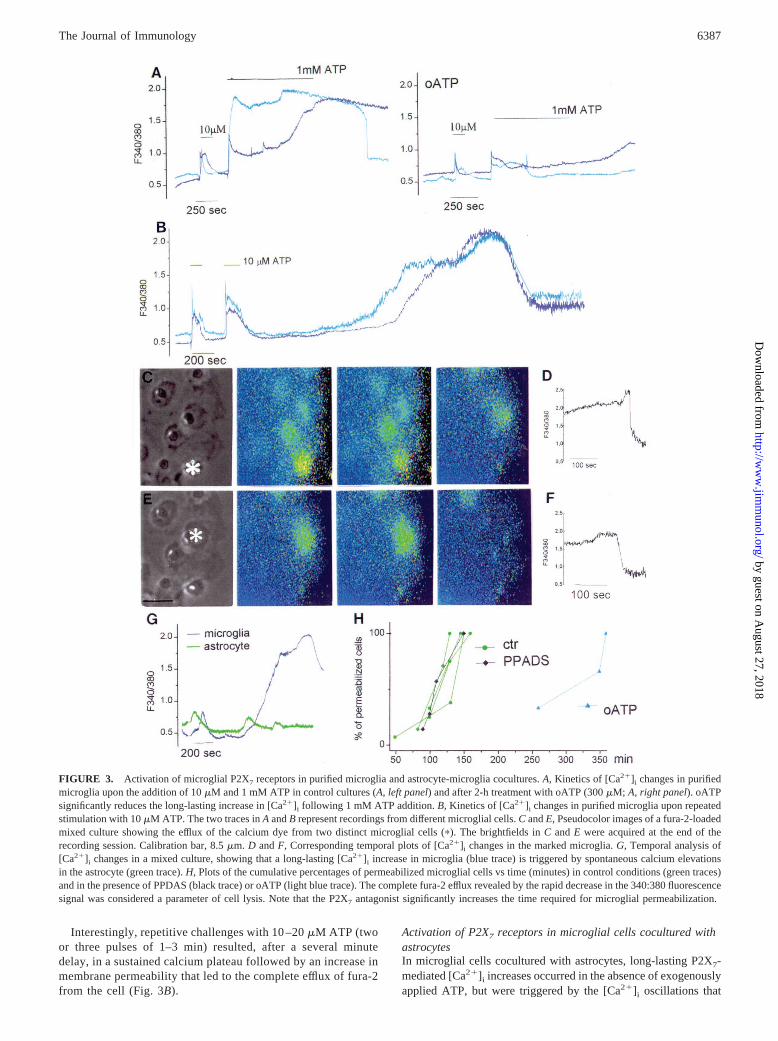
Interestingly, repetitive challenges with 10 –20mM ATP (twoor three pulses of 1–3 min) resulted, after a several minutedelay, in a sustained calcium plateau followed by an increase inmembrane permeability that led to the complete efflux of fura-2from the cell (Fig. 3B).
Activation of P2X7 receptors in microglial cells cocultured withastrocytesIn microglial cells cocultured with astrocytes, long-lasting P2X7-mediated [Ca21]i increases occurred in the absence of exogenouslyapplied ATP, but were triggered by the [Ca21]i oscillations that
FIGURE 3. Activation of microglial P2X7 receptors in purified microglia and astrocyte-microglia cocultures.A, Kinetics of [Ca21]i changes in purifiedmicroglia upon the addition of 10mM and 1 mM ATP in control cultures (A,left panel) and after 2-h treatment with oATP (300mM; A, right panel). oATPsignificantly reduces the long-lasting increase in [Ca21]i following 1 mM ATP addition.B, Kinetics of [Ca21]i changes in purified microglia upon repeatedstimulation with 10mM ATP. The two traces inA andB represent recordings from different microglial cells.C andE, Pseudocolor images of a fura-2-loadedmixed culture showing the efflux of the calcium dye from two distinct microglial cells (p). The brightfields inC andE were acquired at the end of therecording session. Calibration bar, 8.5mm. D andF, Corresponding temporal plots of [Ca21]i changes in the marked microglia.G, Temporal analysis of[Ca21]i changes in a mixed culture, showing that a long-lasting [Ca21]i increase in microglia (blue trace) is triggered by spontaneous calcium elevationsin the astrocyte (green trace).H, Plots of the cumulative percentages of permeabilized microglial cells vs time (minutes) in control conditions (green traces)and in the presence of PPDAS (black trace) or oATP (light blue trace). The complete fura-2 efflux revealed by the rapid decrease in the 340:380 fluorescencesignal was considered a parameter of cell lysis. Note that the P2X7 antagonist significantly increases the time required for microglial permeabilization.
6387The Journal of Immunology
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

spontaneously occur in astrocytes (Fig. 3G). The persistent in-creases in [Ca21]i were followed by fura-2 leakage (Fig. 3G, bluetrace, and Fig. 3,D andF). As in the case of the purified microgliastimulated with 1 mM ATP, the sharp decrease in the fluorescencesignal occurring in microglia cocultured with astrocytes was par-alleled by the disappearance of all microglial cell profiles in thepseudocolor images within 180 min (Fig. 3,C andE, pseudocolorimages; Fig. 3,D andF, temporal plots). Brightfield analysis re-vealed that the cells were still attached to the coverslips (Fig. 3,CandE, asterisks). No changes in [Ca21]i were detected in the pu-rified microglial cultures imaged for up to 6 h in control mediumand in the absence of astrocytes (data not shown), which furtherconfirms that the persistent [Ca21]i increase recorded in coculturesis specifically attributable to the presence of astrocytes.
The sustained [Ca21]i elevations followed by the complete lossof fura-2 in microglia cocultured with astrocytes were greatly de-layed when the cultures were pretreated with the P2X7 antagonistoATP (see the different kinetics of microglial cell lysis in Fig. 3H).On the contrary, the presence of the P2Y/P2X antagonist PPADS(added at the beginning of recording to the extracellular solution)was much less effective in delaying the increase in membrane per-meability (Fig. 3H). These results indicate that the large increase incalcium and membrane permeability of microglial cells may bemediated by the paracrine activation of microglial P2X7 receptorscaused by locally high ATP concentrations or lower ATP concen-trations repeatedly released from astrocytes.
Astrocytes-released ATP mediates cytotoxicity in microglial cells
It has been reported that activation of P2X7 receptors by exoge-nous ATP induces microglial cell death (24), because pore openingradically disrupts ionic homeostasis and causes the depletion ofintracellular low weight metabolites. Fig. 4 shows that microglialcell death is produced by pharmacological treatments that increaseastrocyte [Ca21]i and enhance ATP release; 24-h treatment of as-trocyte-microglia cocultures with repeated challenges of 1mMbradykinin in KRH led to massive microglial death accompaniedby chromatin condensation as revealed by the intercalation of pro-pidium iodide in the DNA (Fig. 4B). The astrocytes present in themixed glial cultures did not show any morphological changes typ-ical of apoptosis (Fig. 4B) and had intact nuclei. The percentage ofcondensed nuclei in microglia associated with astrocytes was73.5% in bradykinin-treated cultures; however, such degenerativechanges occurred to a much lesser extent (18.4%) in the microgliapresent in the same coverslip but not directly in contact with as-trocytes (Fig. 4A). The percentage of condensed nuclei in micro-glia associated with astrocytes in the absence of pharmacological
treatments was 51.5%. Thus, ATP locally released by astrocytes,either spontaneously or upon stimulation, activates P2X7 recep-tors, leading to the apoptotic death of microglial cells.
IFN-g potentiates ATP-mediated calcium signaling betweenastrocytes and microglia
Application of IFN-g to pure astrocyte cultures caused an averageincrease in the frequency of spontaneously occurring [Ca21]i os-cillations of 520% (n5 37 control;n 5 42 IFN-g-treated cells),with a preonset latency of 5–10 min (Fig. 5,A andB). The meanfrequency of [Ca21]i oscillations was derived from 30-min record-ings made before and after application of IFN-g. No significantchanges in the frequency of [Ca21]i oscillations were observedbetween the two 30-min recordings made in the absence of IFN-g(data not shown).
The increased frequency of astrocytic [Ca21]i oscillations wasassociated with a 4- to 5-fold increase in the released ATP (Fig.5C), which was detected in samples of extracellular medium in-cubated with astrocytes for 30 min before and after the addition ofIFN-g using the sensitive luciferase bioluminescence assay (Fig.5C, left panel). The amount of ATP released by astrocytes was alsodetermined using a biological assay (8) in which samples of ex-tracellular medium taken before and after treatment with IFN-gwere gently added to fura-2-loaded astrocytes as ATP sensor cells.Although both samples of extracellular medium evoked a measur-able [Ca21]i response, the [Ca21]i transients induced by the ex-tracellular saline incubated with IFN-g-treated astrocytes were sig-nificantly higher (see quantitation in Fig. 5C, right panel). Basedon a standard dose-response curve of the calcium response ampli-tude vs different ATP concentrations, the actual ATP level in theextracellular medium was estimated to be 1406 16 nM in basalconditions and 6506 45 nM after IFN-g treatment. The localconcentration of ATP at the releasing astrocyte was probablyhigher, as microglial cells do not respond to exogenous applicationof ATP ,10 mM. The [Ca21]i responses were prevented when theconditioned medium was treated with apyrase (30 U/ml for 10–15min), thus indicating that ATP is the bioactive compound releasedin the medium and is responsible for the observed effects.
IFN-g potentiates P2X7 receptor activation and microglialapoptosis
In line with the enhanced stimulation of ATP release from astro-cytes, IFN-g considerably shortened the time of activation of P2X7
receptors in microglial cells cocultured with astrocytes (Fig. 6A).The treatment of mixed glial cells cultures with IFN-g for 24 hexacerbated the apoptotic death of microglial cells, as revealed by
FIGURE 4. Apoptotic changes in micro-glial cells exposed to ATP released from ad-jacent astrocytes.A andB, Fluorescence mi-crographs showing astrocytes and microglialcells following propidium iodide staining af-ter 24-h treatment with repetitive challengesof 1 mM bradykinin in KRH. The astrocyteshave intact nuclei, but clear nuclear conden-sation is visible in the microglia stimulatedwith bradykinin (B). Degenerative changestypical of apoptosis were only occasionallyobserved in the microglial cells exposed tobradykinin in the same coverslip, but in theabsence of neighboring astrocytes (A). Theright panelsin A andB show the correspond-ing brightfields. Calibration bar, 7.2mm.
6388 ATP IN ASTROCYTE-MICROGLIA COMMUNICATION
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
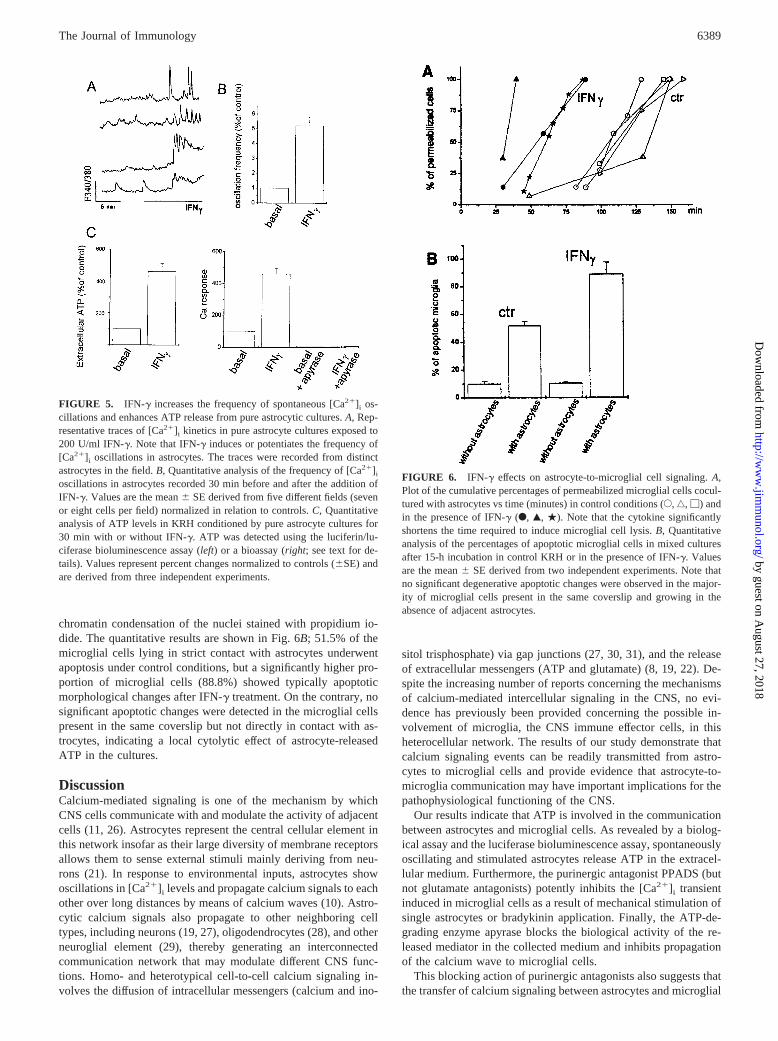
chromatin condensation of the nuclei stained with propidium io-dide. The quantitative results are shown in Fig. 6B; 51.5% of themicroglial cells lying in strict contact with astrocytes underwentapoptosis under control conditions, but a significantly higher pro-portion of microglial cells (88.8%) showed typically apoptoticmorphological changes after IFN-g treatment. On the contrary, nosignificant apoptotic changes were detected in the microglial cellspresent in the same coverslip but not directly in contact with as-trocytes, indicating a local cytolytic effect of astrocyte-releasedATP in the cultures.
DiscussionCalcium-mediated signaling is one of the mechanism by whichCNS cells communicate with and modulate the activity of adjacentcells (11, 26). Astrocytes represent the central cellular element inthis network insofar as their large diversity of membrane receptorsallows them to sense external stimuli mainly deriving from neu-rons (21). In response to environmental inputs, astrocytes showoscillations in [Ca21]i levels and propagate calcium signals to eachother over long distances by means of calcium waves (10). Astro-cytic calcium signals also propagate to other neighboring celltypes, including neurons (19, 27), oligodendrocytes (28), and otherneuroglial element (29), thereby generating an interconnectedcommunication network that may modulate different CNS func-tions. Homo- and heterotypical cell-to-cell calcium signaling in-volves the diffusion of intracellular messengers (calcium and ino-
sitol trisphosphate) via gap junctions (27, 30, 31), and the releaseof extracellular messengers (ATP and glutamate) (8, 19, 22). De-spite the increasing number of reports concerning the mechanismsof calcium-mediated intercellular signaling in the CNS, no evi-dence has previously been provided concerning the possible in-volvement of microglia, the CNS immune effector cells, in thisheterocellular network. The results of our study demonstrate thatcalcium signaling events can be readily transmitted from astro-cytes to microglial cells and provide evidence that astrocyte-to-microglia communication may have important implications for thepathophysiological functioning of the CNS.
Our results indicate that ATP is involved in the communicationbetween astrocytes and microglial cells. As revealed by a biolog-ical assay and the luciferase bioluminescence assay, spontaneouslyoscillating and stimulated astrocytes release ATP in the extracel-lular medium. Furthermore, the purinergic antagonist PPADS (butnot glutamate antagonists) potently inhibits the [Ca21]i transientinduced in microglial cells as a result of mechanical stimulation ofsingle astrocytes or bradykinin application. Finally, the ATP-de-grading enzyme apyrase blocks the biological activity of the re-leased mediator in the collected medium and inhibits propagationof the calcium wave to microglial cells.
This blocking action of purinergic antagonists also suggests thatthe transfer of calcium signaling between astrocytes and microglial
FIGURE 5. IFN-g increases the frequency of spontaneous [Ca21]i os-cillations and enhances ATP release from pure astrocytic cultures.A, Rep-resentative traces of [Ca21]i kinetics in pure astrocyte cultures exposed to200 U/ml IFN-g. Note that IFN-g induces or potentiates the frequency of[Ca21]i oscillations in astrocytes. The traces were recorded from distinctastrocytes in the field.B, Quantitative analysis of the frequency of [Ca21]i
oscillations in astrocytes recorded 30 min before and after the addition ofIFN-g. Values are the mean6 SE derived from five different fields (sevenor eight cells per field) normalized in relation to controls.C, Quantitativeanalysis of ATP levels in KRH conditioned by pure astrocyte cultures for30 min with or without IFN-g. ATP was detected using the luciferin/lu-ciferase bioluminescence assay (left) or a bioassay (right; see text for de-tails). Values represent percent changes normalized to controls (6SE) andare derived from three independent experiments.
FIGURE 6. IFN-g effects on astrocyte-to-microglial cell signaling.A,Plot of the cumulative percentages of permeabilized microglial cells cocul-tured with astrocytes vs time (minutes) in control conditions (E,‚, M) andin the presence of IFN-g (F, Œ, .). Note that the cytokine significantlyshortens the time required to induce microglial cell lysis.B, Quantitativeanalysis of the percentages of apoptotic microglial cells in mixed culturesafter 15-h incubation in control KRH or in the presence of IFN-g. Valuesare the mean6 SE derived from two independent experiments. Note thatno significant degenerative apoptotic changes were observed in the major-ity of microglial cells present in the same coverslip and growing in theabsence of adjacent astrocytes.
6389The Journal of Immunology
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

cells is entirely due to an extracellular messenger. Astrocyte-mi-croglia intercellular communication mediated by gap junctions isunlikely because microglial cells are not dye-coupled with astro-cytes (28). Furthermore, we observed that astrocytic calcium sig-naling efficiently propagates to microglial cells in the presence ofthe gap junction blocker 18a-glycyrrhetinic acid. Given that 18a-glycyrrhetinic acid as well as other gap junction blockers may alsoblock the release of ATP controlled by connexins (32), our datasupport the existence of a calcium-dependent, gap junction block-er-insensitive release of ATP from astrocytes. How ATP crossesthe membrane of astrocytes is still unknown. The existence in as-trocytes of typical secretory granules undergoing regulated secre-tion has been recently reported (33), and a number of synapticproteins for regulated secretion in neuronal cells have been iden-tified in glial cells (34–36). The association of these proteins withATP-storing organelles in astrocytes (36), along with the partialsensitivity of ATP release to clostridial toxins (S. Coco, M. Mat-teoli, and C. Verderio, unpublished observations) suggest that ATPin astrocytes is stored in intracellular vesicles that undergo regu-lated secretion.
The main finding of this study is that astrocyte-released ATPmediates a paracrine activation of microglial P2X7 receptors thattriggers a perturbation of calcium homeostasis and finally leads tomicroglial cell death. This is consistent with previously reporteddata showing that P2X7 activation by exogenously administeredATP mediates cytolysis in mouse as well as in human macro-phages and microglial cells (27, 37–40). Although other P2X re-ceptor subtypes can form a pore permeable to ethidium dyes (41,42), uniquely for the P2X7 receptors this pore dilation progressesto cell lysis (41).
The physiological source of the massive amounts of ATP needed totrigger P2X7 activation in macrophages and microglia in situ is stillnot defined, although it has been proposed to be neighboring dyingcells. Our data identify astrocytes as the major source of endogenousATP. The purine is released in the absence of cell injury as a conse-quence of increased [Ca21]i in astrocytes. Furthermore, our data, byshowing that repetitive challenges with low doses of ATP result inmicroglial cell lysis, provide clues for understanding how P2X7 re-ceptors, which are characterized by a low affinity for ATP, might beactivated in situ. ATP, released by astrocytes following [Ca21]i in-creases, may reach local concentrations sufficient to activate high af-finity purinergic receptors in microglial cells. This repetitive activa-tion could, in turn, lead to either calcium-dependent P2X7 recruitmentto the plasma membrane or induction of an autocrine/paracrine loopresulting in an even higher extracellular ATP concentration, able toactivate P2X7 receptors.
[Ca21]i changes, which occur spontaneously (20) or in responseto different kinds of stimuli (8, 9) in cultured astrocytes, have beenshown to occur in brain as a consequence of neuronal activationand glutamate release (26). Alterations of calcium homeostasis inastrocytes have been suggested to occur in specific pathophysio-logical conditions of the CNS (43). Interestingly, we have foundthat the inflammatory cytokine IFN-g increases the frequency ofspontaneous calcium oscillations in glial cells. IFN-g receptorstimulation has been previously associated with elevation ofcAMP, elevations of inositol trisphosphate that cause release ofcalcium from stores, and protein kinase C activation in cells of theimmune system and astrocytes (44). The effect of IFN-g on astro-cyte calcium homeostasis potentiates astrocyte-to-microglia com-munication, with the enhanced ATP release from astrocytes pre-sumably accounting for the increased apoptosis of microglial cells.This finding adds to the reported up-regulation of P2X7 receptorscaused by prolonged IFN-g treatment in human macrophages andmouse microglial cells (38, 25).
The activation of P2X7 receptors induces IL-1b release frommicroglia (15, 45). It has recently been reported that IL-1b signif-icantly down-regulates gap junction connectivity among astrocytesand also potentiates interastrocyte calcium signaling mediated bythe extracellular messenger ATP (46). Microglial cells activated byATP released from astrocytes upon IFN-g treatment can thereforefeed back to glial cells via IL-1b and thus activate a paracrine loop,sustaining further ATP release.
The repeated secretion of ATP from astrocytes and the apoptoticresponse in microglial cells may represent a homeostatic mecha-nism for controlling the number of microglial cells in pathophys-iological conditions of the CNS. It is interesting to note that anincreased number of microglial cells undergoing apoptosis hasbeen reported in inflammatory demyelinating diseases, includingexperimental autoimmune encephalomyelitis in rats (47, 48). Inacute multiple sclerosis (MS) plaques, in particular, microglia andmacrophages represent a relevant percentage (;60%) of theTUNEL-positive cells (49, 50). It is noteworthy that IFN-g hasbeen selectively detected in active plaques, where it is predomi-nantly found on astrocytes (51). As the interaction between mi-croglia and T cells is important in the development of CNS in-flammation, the apoptosis of microglia could be related to thedevelopment and progression of MS, as it has been shown thatAg-stimulated T cells undergo apoptosis in the presence of micro-glial cells (52–54). By acting locally to kill activated T cells, mi-croglia seem to be a component of the so-called immunologicalbrain barrier (3) and may contribute to the down-regulation ofimmunopathologic processes in the brain. Our data showing that inthe presence of the inflammatory cytokine IFN-g, astrocytes in-duce extensive microglia apoptosis due to the release of ATP sug-gest that this mechanism may operate during MS inflammation andmay thus represent a potential target for therapeutic strategies.
AcknowledgmentsWe thank Drs. Elena Pravettoni, Silvia Coco, and Barbara Viviani for helpwith some experiments, and Prof. Giulio Levi, Dr. Sergio Visentin (IstitutoSuperiore di Sanita, Rome, Italy), and Dr. Marco Righi (Consiglio Nazio-nale delle Ricerche, Milan, Italy) for stimulating discussions. We thankProf. Francesco Clementi (University of Milan, Milan, Italy) and Prof.Guido Fumagalli (University of Verona, Verona, Italy) for critically read-ing the manuscript.
References1. Gehrmann, J., Y. Matsumoto, and G. W. Kreutzberg. 1995. Microglia: intrinsic
immunoeffector cell of the brain.Brain Res. Brain Rev. 20:269.2. Kreutzberg, G. W. 1996. Microglia: a sensor for pathological events in the CNS.
Trends Neurosci. 19:312.3. Bechmann, I., G. Mor, J. Nilsen, M. Eliza, M., R. Nitsch, and F. Naftolin. 1999.
FasL (CD95L, Apo 1L) is expressed in the normal rat and human brain: evidencefor the existence of an immunological brain barrier.Glia 27:62.
4. Dubyak, G. R., and C. El-Moatassim. 1993.Signal transduction via P2-purinergicreceptors for extracellular ATP and other nucleotides.Am. J. Physiol. 265:C577.
5. Neary, J. T., M. P. Rathbone, F. Cattabeni, M. P. Abbracchio, and G. Burnstock.1996. Trophic actions of extracellular nucleotides and nucleosides on glial andneuronal cells.Trends Neurosci. 19:13.
6. Apasov, S., M. Koshiba, F. Redegaeld, and M. Sitkovsky. 1995. Role of extra-cellular ATP and P1 and P2 classes of purinergic receptors in T-cell developmentand cytotoxic T lymphocyte effector functions.Immunol. Rev. 146:5.
7. Brake, A. J., and D. Julius. 1996. Signaling by extracellular nucleotides.Annu.Rev. Cell Dev. Biol. 12:519.
8. Guthrie, P. B., J. Knappenberg, M. Segal, M. V. L. Bennett, A. C. Charles, andS .B. Kater. 1999. ATP released from astrocytes mediates glial calcium waves.J. Neurosci. 19:520.
9. Queiroz, G., P. J. Gebicke-Haerter A. Schobert, K. Starke, and I. Von Kugelgen.1997. Release of ATP from cultured rat astrocytes elicited by glutamate receptoractivation.Neuroscience 78:1203.
10. Cornell-Bell, A. H., S. M. Finkbeiner, M. S. Cooper, and S. J. Smith. 1990.Glutamate induces calcium waves in cultured astrocytes: long-range glial signal-ing. Science 247:470.
11. Pasti, L., T. Pozzan, and G. Carmignoto. 1995. Long-lasting changes of calciumoscillations in astrocytes.J. Biol. Chem. 270:15203.
12. Kettenmann, H., R. Banati, and W. Walz. 1993. Electrophysiological behaviourof microglia.Glia 7:93.
6390 ATP IN ASTROCYTE-MICROGLIA COMMUNICATION
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

13. Walz, W., S. Ilschner, C. Ohlmeyer, R. Banati, and H. Kettenmann. 1993. Ex-tracellular ATP activates a cation conductance and a K1 conductance in culturedmicroglial cells from mouse brain.J. Neurosci. 13:4403.
14. Norenberg, W., J. M. Langosch, P. J. Gebicke-Haerter, and I. P. Lles. 1994.Characterization and possible function of adenosine 59-triphosphate receptors inactivated rat microglia.Br. J. Pharmacol. 111:942.
15. Ferrari, D., M. Villalba, P. Chiozzi, S. Falzoni, P. Ricciardi-Castagnoli, andF. Di Virgilio. 1996. Mouse microglial cells express a plasma membrane poregated by extracellular ATP.J. Immunol.156:1531.
16. Visentin, S., M. Renzi, C. Frank, A. Greco, and G. Levi. 1999. Two differentionotropic receptors are activated by ATP in rat microglia.J. Physiol. 519:723.
17. Moller, T., O. Kann, A. Verkhratsky, and H. Kettenmann, H. 2000. Activation ofmouse microglial cells affects P2 receptor signaling.Brain Res. 853:49.
18. Banker, G. A., and W. M. Cowan. 1977. Rat hippocampal neurons in dispersedcell culture.Brain Res. 126:379.
19. Parpura, V., T. A. Basarsky, F. Liu, S. Jeftinijia, and P. G. Haydon. 1994. Glu-tamate-mediated astrocyte-neuron signalling.Nature 369:744.
20. Fatatis, A., and J. T. Russell. 1992. Spontaneous changes in intracellular calciumconcentration in type I astrocytes from rat cerebral cortex in primary culture.Glia5:95.
21. Giaume, C., and L. Venance. 1998. Intercellular calcium signaling and gap junc-tional communication in astrocytes.Glia 24:50.
22. Innocenti, B., V. Parpura, and P. G. Haydon. 2000. Imaging extracellular wavesof glutamate during calcium signaling in cultured astrocytes.J. Neurosci. 20:1800.
23. Surprenant, A., F. Rassendren, E. Kawashima, R. A. North, and G. Buell. 1996.Science 272:735.
24. Ferrari, D., P. Chiozzi, S. Falzoni M. Dal Susino, G. Collo, G. Buell, andF. Di Virgilio. 1997. ATP-mediated cytotoxicity in microglial cells.Neurophar-macology 36:1295.
25. Falzoni, S., M. Munerati, D. Ferrari, S. Spisani, S. Moretti, andF. Di Virgilio.1995. The purinergic P2Z receptor of human macrophage cells:characterization and possible physiological role.J. Clin. Invest. 95:1207.
26. Pasti, L., A. Volterra, T. Pozzan, and G. Carmignoto. 1997. Intracellular calciumoscillations in astrocytes: a highly plastic, bidirectional form of communicationbetween neurons and astrocytes in situ.J. Neurosci. 17:7817.
27. Nedergaard, M. 1994. Direct signalling from astrocytes to neurons in cultures ofmammalian brain cells.Science 263:1768.
28. Venance, L., J. Cordier, M. Monge, B. Zalc, J. Glowinski, and C. Giaumie. 1995.Homotypic and heterotypic coupling mediated by gap junctions during glial celldifferentiation in vitro.Eur J. Neurosci. 7:451.
29. Newman, E. A., and K. R. Zahs. 1998. Modulation of neuronal activity by glialcells in the retina.J. Neurosci. 18:4022.
30. Boitano, S., E. R. Dirksen, and M. J. Sanderson. 1992. Intercellular propagationof calcium waves mediated by inositol triphosphate.Science 258:292.
31. Froes, M. M., A. H. P. Correira, J. Garcia-Abreu, D. C. Spray,A. C. Campos de Carvalho, and V. M. Neto. 1999. Gap-junctional couplingbetween neurons and astrocytes in primary central nervous system cultures.Proc.Natl. Acad. Sci. USA 96:7541.
32. Cotrina, M. L., J. H.-C. Lin, A. Alves-Rodrigues, S. Liu, J. Li, H. Azmi-Ghadimi,C. C. G. Naus, and M. Nerdergard. 1998. Connexin regulate calcium signaling bycontrolling ATP release.Proc. Natl. Acad. Sci. USA 95:15735.
33. Calegari, F., S. Coco, E. Taverna, M. Bassetti, C. Verderio, N. Corradi,M. Matteoli, and P. Rosa. 1999. A regulated secretory pathway in cultured hip-pocampal astrocytes.J. Biol. Chem. 274:22539.
34. Parpura, V., Y. Fang,. T. Basarsky, R. Jahn, and P. G. Haydon. 1995. Expressionof synaptobrevin II, cellubrevin and syntaxin but not SNAP 25 in cultured as-trocytes.FEBS Lett. 377:744.
35. Jeftinija, S. D., K. V. Jeftinija, and G. Stefanovic. 1997. Cultured astrocytesexpress proteins involved in vesicular glutamate release.Brain Res. 750:41.
36. Maienschein, V., M. Marxen, W. Volknandt, and H. Zimmermann. 1999. Aplethora of presynaptic proteins associated with ATP-storing organelles in cul-tured astrocytes.Glia 26:233.
37. Steinberg, T. H., A. S. Newman, J. A. Swanson, and S. C. Silverstein. 1987ATP4- permeabilizes the plasma membrane of mouse macrophages to fluorescentdyes.J. Biol. Chem. 262:8884.
38. Blanchard, D. K., S. McMillen, and J. Y. Djeu. 1991. IFN-g enhances sensitivityof human macrophages to extracellular ATP-mediated lysis.J. Immunol. 147:2579.
39. Murgia, M., P. Pizzo, T. H. Steinberg, and F. Di Virgilio. 1992. Characterizationof the cytotoxic effect of extracellular ATP in J774 mouse macrophages.Bio-chem. J. 288:897.
40. Molloy, A., P. Loachumroonvorapong, and G. Kaplan. 1994. Apoptosis, but notnecrosis, of infected monocytes is coupled with killing of intracellular bacillusCalmette-Guerin.J. Exp. Med. 180:1499.
41. Virginio, C., A. Mackenzie, F. A. Rassendren, R. A. North, and A. Surprenant.1999. Pore dilation of neuronal P2X receptor channels.Nat. Neurosci. 2:315.
42. Khakh, B. S., X. R. Bao, C. Labarca, and H. A. Lester. 1999. Neuronal P2Xtransmitter-gated cation channels change their ion selectivity in seconds.Nat.Neurosci. 2:322.
43. Zanotti, S., and A. Charles. 1997. Extracellular calcium sensing by glial cells: lowextracellular calcium induces calcium release and intercellular signaling.J Neu-rochem. 69:594.
44. Holliday, J., and D. L. Gruol. 1993. Cytokine stimulation increases intracellularcalcium and alters the response to quisqualate in cultured cortical astrocytes.Brain Res. 621:233.
45. Sanz, J. M., and F. Di Virgilio. 2000. Kinetics and mechanism of ATP-dependentIL-b release from microglial cells.J. Immunol. 164:4893.
46. John, G. R., E. Scemes, S. O. Suadicani, J. S. H. Liu, P. C. Charles, S. C. Lee,D. C. Spray, and C. F. Brosnan. 1999. IL-1b differentially regulates calcium wavepropagation between primary human fetal astrocytes via pathways involving P2receptors and gap junction channels.Proc. Natl. Acad. Sci. USA 96:11613.
47. White, C. A., P. A. McCombe, and M. P. Pender. 1998. Microglia are moresusceptible than macrophages to apoptosis in the CNS in experimental autoim-mune encephalomyelitis through a mechanism not involving Fas (CD95).Int.Immunol. 10:935.
48. Bonetti, B., J. Pohl, Y. L. Gao, and C. S. Raine. 1997. Cell death during auto-immune demyelination: effector but not target cells are eliminated by apoptosis.J. Immunol. 159:5733.
49. Dowling, P., W. Husar, J. Menona, H. Donnenfeld, S. Cook, and M. Sidhu.1997.Cell death and birth in multiple sclerosis brain.J. Neurol. Sci. 149:1.
50. Bonetti, B., and C. S. Raine. 1997. Multiple sclerosis: oligodendrocytes displaycell death-related molecules in situ but do not undergo apoptosis.Ann. Neurol.42:74.
51. Traugott, U., and P. Lebon. 1988. Multiple sclerosis: involvement of interferonsin lesion pathogenesis.Ann. Neurol. 24:243.
52. Klyushnenkova, E. N., and P. Vanguri.1997. Ia expression and antigen presen-tation by glia: strain and cell type-specific differences among rat astrocytes andmicroglia.J. Neuroimmunol. 79:190.
53. Ford, A. L., E. Foulcher, F. A. Lemckertand, and J. D. Sedgwick. 1996. Microgliainduce CD4 T lymphocyte final effector function and death.J. Exp. Med. 184:1737.
54. Frigerio, S., V. Silei, E. Ciusani, G. Massa, G. M. Lauro, and A. Salmaggi. 2000.Modulation of Fas-ligand (Fas-L) on human microglial cells: an in vitro study.J. Neuroimmunol. 105:109.
6391The Journal of Immunology
by guest on August 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from