articulo geoquimica
DESCRIPTION
articulo de geoquimicaTRANSCRIPT

The American Association of Petroleum Geologists Bulletin
V 64, No. 8 (August 1980) P. 1179-1209, 18 Figs.
Anoxic Environments and Oii Source Bed Genesis'
G. J. DEMAISON^ and G. T. MOORE3
Abstract The anoxic aquatic environment is a mass of water so depleted in oxygen that virtually all aerobic biologic activity has ceased. Anoxic conditions occur where the demand for oxygen in the water column exceeds the supply. Oxygen demand relates to surface biologic productivity, whereas oxygen supply largely depends on water circulation, which is governed by global climatic patterns and the Coriolis force.
Organic matter in sediments ttelow anoxic water is commonly more abundant and more lipid-rich than under oxygenated water mainly because of the absence of benthonic scavenging. The specific cause for preferential lipid enrichment probably relates to the biochemistry of anaerobic bacterial activity. Geochemical-sedimentologic evidence suggests that potential oil source beds are and have been deposited in the geologic past in four main anoxic settings as follows.
1. Large anoxic lakes—Permanent stratification promotes development of anoxic bottom water, particularly in large lakes which are not subject to seasonal overturn, such as Lake Tanganyika. Warm equable climatic conditions favor lacustrine anoxia and nonma-rine oil source bed deposition. Conversely, lakes in temperate climates tend to be well oxygenated.
2. Anoxic silled basins—Only those landlocked silled basins with positive water balance tend to become anoxic. Typical are the Baltic and Black Seas. In arid-region seas (Red and Mediterranean Seas), evaporation exceeds river inflow, causing negative water balance and well-oxygenated bottom waters. Anoxic conditions in silled basins on oceanic shelves also depend upon overall climatic and water-circulation patterns. Silled basins should be prone to oil source bed deposition at times of woridwide transgression, both at high and low paleolatltudes. Sllled-basin geometry, however, does not automatically imply the presence of oil source beds.
3. Anoxic layers caused by upwelling—^These develop only when the oxygen supply in deep water cannot match demand owing to high surface biologic productivity. Examples are the Benguela Current and Peru coastal upwelling. No systematic correlation exists between upwelling and anoxic conditions because deep oxygen supply is often sufficient to match strongest demand. Oil source beds and phosphorites resulting from upwelling are present preferentially at low paleolatltudes and at times of woridwide transgression.
4. Open-ocean anoxic layers—^These are present in the oxygen-minimum layers of the northeastern Pacific and northern Indian Oceans, far from deep, oxygenated polar water sources. They are analogous, on a reduced scale, to woridwide "oceanic anoxic events" which occurred at ^lot}al climatic warmups and major transgressions^ as in Late Jurassic and middle Cretaceous times. Known marine oil source bed systems are not randomly distributed in time but tend to coincide with periods of worldwide transgression and oceanic anoxia.
Geochemistry, assisted by paleogeography, can greatly help petroleum exploration by identifying pa-leoanoxic events and therefore widespread oil source bed systems in the stratigraphic record. Recognition of the proposed anoxic models in ancient sedimentary biasins should help In regional stratigraphic mapping of oil shale and oil source beds.
INTRODUCnON The most significant progress made in petro
leum geology in the last 10 years has been the attainment of a satisfactory understanding of the processes of oil and gas generation and destruction in sedimentary basins. Geochemical techniques now routinely identify oil source beds by analysis of rock samples retrieved from deep wells. However, geologists often question the validity of projecting the presence of source beds identified from a few wells and a few grams of rocks, to entire sedimentary basins. Until the last few years these hesitations were legitimate; it was difficult, if not inconceivable, to map oil source beds without a clear understanding of their depo-sitional environment.
Fortunately, recent oceanographic and geochemical observations now make it possible to comprehend many formerly obscure aspects of the genesis and stratigraphic distribution of petroleum source beds. It is timely, thus, to review depositional environments of oil source beds considering these recent findings and address ourselves to the following questions.
1. What modem sediments are precursors to oil source beds?
©Copyright 1980. The Atnerican Association of Petroleum Geologists. All rights teserved.
AAPG grants permission for i single photocopy of this article for research purposes. Other photocopying not allowed by the 1978 Copyright Law is prohibited. For more than one photocopy of this article, users should send request, article identification number (see below), and $3.00 per copy to Copyright Clearance Center, Inc., 21 Congress Street, Salem, MA 01970.
'The original version of this paper was read at the 10th International Congress on Sedimentology, Jerusalem, 1978, and published in Organic Geochemistry. Manuscript received, August 24, 1979; accepted, January 17, 1980.
^Chevron Overseas Petroleum Inc., San Francisco, California 94105.
^Chevron Oil Field Research Co., La Habra, California 90631.
The writers are indebted to R. R. Hammes. R. W. Jones, R. A. Lagaay, R. S. Oremland, W. D. Redfield, S. R. Silverman, and J. A. Sutherland for critical reviews of the manuscript.
Published with permission of Chevron Overseas Petroleum Inc. and Chevron Oil Field Research Co.
Article Identification Number O149-1423/80/B0O8-0O02$03.0O/0
1179

1180 G. J. Demaison and G. T. Moore
2. What factors affect the preservation of organic matter in aquatic environments?
3. Why are anoxic conditions more favorable than oxic conditions for organic-matter preservation, both in quaUty and quantity?
4. What causes favor anoxic conditions in lakes, seas, and oceans?
5. Can natural anoxic settings be scientifically classified to help oceanographers, stratigraphers, and petroleum explorationists?
PRECURSOR SEDIMENTS TO OIL SOURCE BEDS
Potential oil source beds are organic-rich sediments containing a kerogen type that is sufficiently hydrogen-rich (type I or type II; Tissot et al, 1974) to convert mainly to oil during thermal maturation. Kerogen type and thus oil source character in ancient sediments is identified through such approaches as elemental analysis of kerogen and whole-rock pyrolysis with additional support from microscopic organic analysis (Tissot and Welte, 1978, p. 81-91; Hunt, 1979, p. 454-472).
Evaluation of organic content, which can be a gross quantitative index of oil generative potential // kerogen is oil-prone, is measured by the amount of organic carbon present in sediments. Documented oil source beds and oil shales around the world always contain hydrogen-rich kerogen and fall into a range of organic carbon content between about 1% and over 20% by weight. The boundary between very rich oil source beds and oil shales is determined by mining and processing economics.
Rich to very rich marine oil source beds and oil shales commonly contain higher than average uranium, copper, and nickel concentrations. Uranium content in most marine oil shales commonly shows a positive correlation with oil yield upon rock pyrolysis (Swanson, 1960).
The measurement of organic carbon in sediments alone is insufficient to identify potential oil source beds. Transported terrestrial organic matter, oxidized aquatic organic matter, and reworked organic matter from a previous sedimentary cycle can create levels of organic carbon in marine sediments up to about 4%. Yet this abnormally concentrated organic matter is hydrogen-poor, gas-prone, and without significant oil generating potential (Tissot et al, 1974; Demaison and Shibaoka, 1975; Dow, 1977). This is essentially the organic facies that has been described in middle Cretaceous marine black shales encountered by several Deep Sea Drilling Project holes in the northwestern Atlantic Basin (Tissot et al, 1979). An identical situation is present today on the Arctic Shelf of the USSR where high organic carbon concentrations in marine sediments result
from the influx of large amounts of terrestrial organic matter brought in by fluvial discharge (Bezrukov et al, 1977).
In summary, a high organic carbon content in sediments is not necessarily an indication of oil source rock precursor character. Additional geo-chemical evidence, such as measurement of hydrogen richness of humic substances and kerogen, pyrolysis yield of whole sediment, and overall soluble-Upid content of the sediment, is needed to ascertain a possible oil source precursor character.
FACTORS INFLUENCING ORGANIC MATTER ACCUMULATION IN AQUATIC ENVIRONMENTS
Factors capable of influencing organic-matter accumulation in sediments are both biologic and physical. Biologic factors include primary biologic productivity of the surface-water layers and of adjoining landmasses and biochemical degradation of dead organic matter by metazoan and microbial scavengers. Physical factors include modes of transit of organic matter to depositional sites, sediment particle size, and sedimentation rates. These factors interact to determine quaUta-tive and quantitative preservation of organic matter in sediments.
Primary Biologic Productivity The principal source of aquatic organic matter
is the phytoplankton (Bordovsky, 1965) composed largely of single-cell microscopic algae residing in the uppermost layers of water illuminated by sunlight, the euphotic zone. The main limiting factor to planktonic productivity, in addition to light, is the availability of mineral nutrients, particularly nitrates and phosphates, which are in short supply in the euphotic zone. Phytoplankton are intensively grazed by zoo-plankton. Both phytoplankton and zooplankton are then consumed by large invertebrates and fish.
The other source of organic matter in the aquatic environment is transported terrestrial organic matter from streams and rivers. Land-plant productivity is largely dependent on the amount of rainfall on supporting landmasses. Because terrestrial organic matter has undergone considerable degradation in subaerial soils prior to its transport, it is usually hydrogen depleted and refractory in nature.
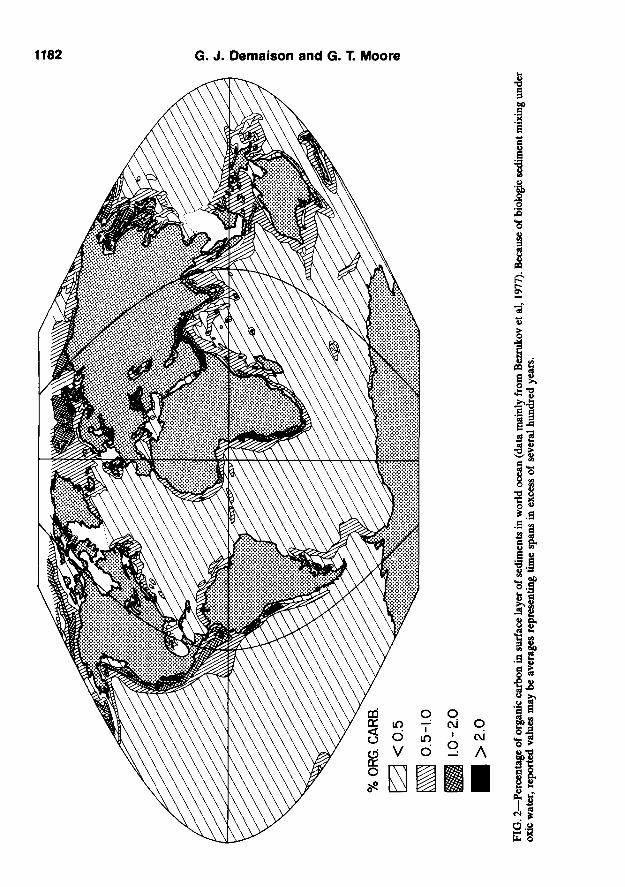
The traditional view is that fields of high surface productivity in the ocean should be associated with high organic enrichment of underlying sediments; however, after exhaustive investigation, we could not find a systematic correlation between primary biologic productivity (Fig. 1)

mg
Clm
^ da
y
^ <
10
0
^ 1
00
-25
0
^ >
25
0
o
K m
3 < 3 3 3 « 3 0)
PIG
. 1
Prim
ary
biol
ogic
pro
duct
ion
in w
orld
oce
an, e
xpre
ssed
in
mill
igra
ms
of o
rgan
ic c
arbo
n pe
r sq
uare
met
er p
er d
ay (m
odifi
ed f
rom
Kob
lent
z-M
ishk
e et
al,
1970
). U
nice
llula
r, m
icro
scop
ic, p
lank
toni
c al
gae
(phy
topl
ankt
on)
are
prin
cipa
l pr
imar
y pr
oduc
ers
of o
rgan
ic m
atte
r in
sea
.
09
% O
RG
. C
AR
B.
<0
.5
O i 0)
M' o
3 fi> o
o o
3
FIG
. 2—
Per
cent
age
of o
rgan
ic c
arbo
n in
sur
face
lay
er o
f se
dim
ents
in
wor
ld o
cean
(da
ta m
ainl
y fr
om B
ezni
kov
et a
l, 19
77).
Bec
ause
of
biol
ogic
sed
imen
t m
ixin
g im
der
oxic
wat
er,
repo
rted
val
ues
may
be
aver
ages
rep
rese
ntin
g ti
me
span
s in
exc
ess
of s
ever
al h
undr
ed y
ears
.

Anoxic Environments 1183
and organic carbon content of bottom sediments in the oceans (Fig. 2). Some areas of high productivity like the Peruvian Shelf and Southwest Shelf of Africa were enriched; others of equally high productivity like the Grand Banks of Newfoundland, the northeastern Brazilian Shelf, or the Northwest Shelf of Africa were not. Clearly, factors other than surface productivity had to be evaluated.
Biochemical Degradatioii of Dead Organic IVfatter
Dead organic matter is inherently unstable thermodynamically and seeks its lowest level of free energy in any given environment. Above all, it serves as a source of energy and nutrients for Uving organisms. Heterotrophic microorganisms, mainly bacteria, play a critical role in decomposing organic matter in the water column, in the interstitial water of sediments, and in the digestive tracts of animal scavengers. In marine basins bacterial biomass approaches that of the phy-toplankton (Bordovsky, 1965). Bacterial degradation proceeds quickly and efficiently in aerobic (oxygen-rich) water. The overall oxidation processes by aerobic bacteria are illustrated by:
(CH2O) + O2 ^ CO2 H- H2O.
When the oxygen supply becomes exhausted, oxidation of organic matter by anaerobic bacteria using nitrates as a source of oxygen (electron acceptor) takes over according to the generalized scheme:
(CH2O) -I- 4NO3 -^ 6C02-I- 6H2O -I- 2N2.
Once nitrates have been exhausted, degradation of organic matter continues by anaerobic bacteria using sulfates as the oxidant by the generalized reaction scheme:
CH2O -I- SO4 ->• CO2 -I- H2O -I- H2S.
The last and least efficient step in anaerobic metabohsm is fermentation. Here carboxyl (CO2) groups and organic acids of the organic matter itself, or from bacterial breakdown, are employed as electron acceptors. A special type of anaerobic fermentation is bacterial methanogenesis (Clay-pool and Kaplan, 1974). It is a complex process carried out in several steps by types of bacteria which degrade one another's by-products (Man-heim, 1976). The methane-generating (methano-genic) bacteria occupy the terminal niche of the anaerobic food web and produce methane by CO2 reduction by H2 or by attacking acetate, formate, or methanol (Wolfe, 1971). Higher hydro
carbons, at least up to butane, are always present together with bacterial methane in anoxic water, although in much smaller amounts (Deuser, 1975). Anaerobic methane fermentation is common below the sulfate reducing zone in recent marine sediments and prevails in anoxic waters where both nitrate and sulfate levels are low (e.g., in freshwater lakes). Methane fermentation and sulfate reduction, however, are not mutually exclusive (Oremland and Taylor, 1978). The processes mentioned are presented in a simplified form. They represent an important developing area of research in geomicrobiology.
Anaerobic degradation is thermodynamically less efficient than aerobic decomposition (Clay-pool and Kaplan, 1974) and results in a more lip-id-rich and more reduced (hydrogen-rich) organic residue than does aerobic degradation (Force and McCarty, 1970; Beliaeva and Romankevich, 1976; Gerschanovich et al, 1976; Pelet and De-byser, 1977; Didyk et al, 1978). Moreover, under such conditions, a significant fraction of the preserved organic matter consists of remains of the bacterial biomass itself (Lijmbach, 1975). The mechanisms leading to this preferential enrichment in lipids are Uttle understood and in need of fundamental research. The enhanced preservation of hydrogen-rich and lipid-rich organic matter in sediments deposited under anoxic water is critical for the genesis of oil source beds.
Explanations proposed for the higher than normal organic matter concentrations observed under anoxic water (Richards, 1970; Deuser, 1971) include the suggestions that anaerobic degradation is inherently slower (Lijmbach, 1975) or that enhanced preservation might be a consequence of high sedimentation rates (Richards, 1970). (The latter is discussed under the effects of other physical parameters.)
As far as biologic factors are concerned, Force and McCarty (1970) have shown by laboratory experiments that rates of anaerobic degradation of algae by sulfate reduction are identical to those of aerobic degradation. Observations in the natural environment by Orr and Gaines (1974) led to parallel conclusions. This suggests that most of the organic matter consumption at oxygen-rich ("oxic") sediment/water interfaces is by the meta-zoan, not by the microbial population (Degens and Mopper, 1976).
Under an oxic water column (Fig. 3) the benth-ic fauna actively scavenges and reworks the "organic rain" falling through the water column. Except in shallow waters where sunlight penetrates to the bottom, there is very Uttle primary production and mainly consumption of organic matter at the benthic boundary. Bottom muds under an oxygen-rich water column are commonly anoxic

1184 G. J. Demaison and G. T. Moore
OXIC ENVIRONMENT
TYPE OF RESPIRATION
RESIDENCE TIME
OF O.M.
POORER CM. PRESERVATION (0.2-4%T.O.C.) LOWER QUALITY O.M.
BIOLOGICAL REWORKING IS ENHANCED BY:
. PRESENCE OF ANIMAL SCAVENGERS AT INTERFACE.
. BIOTURBATION BY WORMS FACILITATES DIFFUSION OF OXIDANTS (02,S04) IN SEDIMENTS.
. LESSER ORGANIC COMPLEXATION WITH TOXIC METALS.
OXYGEN CONSUMPTION
days-mos.
SULFATE 1 REDUCTION ••
BACTERIAL CO2 REDUCTION
750 yrs
500 yrs 6
750 yrs ^
U=50cm/1000yrs
FIG. 3—Degradation of organic matter under oxygenated (oxic) water column. Organic carbon concentration under oxygen-rich water is closely related to sedimentation rate, without regard to surface primary productivity.
but nevertheless extensively disrupted by oxygen-respiring invertebrates such as polychaetes (worms), holothurians, and bivalves. The most common modes of feeding are on particulate organic matter and bacteria in water above the sediment (suspension feeders), and on organic matter and bacteria present within the sediment itself (deposit feeders). Mobile deposit feeders (burrow-ers) cause mixing and transport of particles, as well as irrigation of sediments, thereby accelerating geochemical processes at or below the sediment-water interface (Rhoads, 1974; Aller, 1978). Many who have studied deep-sea sediments believe that the sediments are mixed to a depth of 5 to 30 cm by biologic activity (Peng et al, 1977). Bioturbation is ubiquitous under oxic water where it has been observed at all water depths, including the deep-sea sediments of the abyssal realm. The range of mixing (bioturbation) rates in the deep sea is comparable to that in nearshore regions, with no apparent correlation between the mixing rate and the sediment type or sedimentation rate (Turekian et al, 1978). This suggests that mixing rate may be almost solely related to benthic biomass variabihty, and thus to productivity in surface waters and subsequent dehvery of edible organic material to the ocean floor.
Under an anoxic water column (Fig. 4), how
ever, oxygen depletion (even in the absence of hydrogen sulfide) depresses and eventually eUmi-nates benthonic metazoan Ufe (Theede et al, 1969). The benthic metazoan biomass is unaffected by oxygen concentrations as low as about 1 ml/1 (Rhoads, 1974), but between 0.7 ml/1 and 0.3 ml/l it sharply decreases by about a factor of 5 (Rosenberg, 1977). Below 0.3 ml/1, deposit feeders become rare, less active, and soft bodied only. Eventually bioturbation ceases. Below 0.1 ml/l, the suspension feeders disappear, leaving anaerobic bacteria as the only effective reworkers of organic matter.
Once organic matter is incorporated in the anoxic sediment itself, the lack of bioturbation acts as a limiting factor to diffusion of oxidants into the sediment—hence bacterial sulfate reduction is slowed down if not completely arrested. A classic observation is that the pore fluids of anoxic sediments are depleted in sulfates (Manhetm, 1976, p. 115-118). Lack of bioturbation under anoxic water results in laminated and organic-rich sediments. Reducing conditions imder anoxic water make certain toxic metals, like lead, available for chelation with organic matter (Jones, 1973). Degens and Mopper (1976) have suggested that metal complexation of organic matter makes it less susceptible to microbial at-

Anoxic Environments 1185
ANOXIC ENVIRONMENT BETTER O.M. PRESERVATION {1-25%T.0.C.)
HIGHER QUALITY O.M.
BIOLOGICAL REWORKING IS SLOWED BY:
.THE ABSENCE OF ANIMAL SCAVENGERS.
. RESTRICTED DIFFUSION OF OXIDANTS (SO4) INTO UNDISTURBED SEDIMENT.
.LESSER UTILIZATION OF LIPIDS BY ANAEROBIC BACTERIA.?
TYPE OF RESPIRATION
OXYGEN CONSUMPTION
RESIDENCE TIME
OF O.M.
BACTERIAL SULFATE
REDUCTION
days - mos.
OCEAN BOTTOM
BACTERIAL CO, REDUCTION
•100 cm •
U=50cm/lOOOyrs
FIG. 4—Degradation of organic matter under anoxic water column. Bioturbation becomes minimal at oxygen concentrations below 0.5 ml/1. This concentration, rather than total absence of oxygen, is effective "biochemical fence" between poor and good organic matter preservation.
tack. Comparison of Figures 3 and 4 shows that, giv
en identical sedimentation rates, dead organic matter in oxygenated environments is exposed to oxidants much longer than in anoxic environments, owing to bioturbation. For a given sedimentation rate, bioturbation imder oxic water prolongs the exposure of organic matter to oxidation by hundreds of years, which strongly enhances degradation. Added to this effect is the grazing of the sediment bacterial biomass by mobile deposit feeders responsible for bioturbation.
IVfo«|es of Tnmsit of Organic Matter to Depositknial Sites
E>ead zooplankton, unconsumed algal cells, fecal pellets, and fish carcasses originating in the shallow euphotic zone continuously sink to the bottom. The speed of fall of particulate organic matter is exceedingly slow and ranges from 0.10 to 5 m per day, according to their shape and size, the smallest particles being the slowest to reach bottom. The transit time for fecal pellets is much faster (Spencer et al, 1978). It is estimated that their passage through a water column 4 km deep will not exceed 15 days.
If organic debris falls through a well-oxygenat
ed water column in which animal life is abundant, it is consumed by scavenging animals, until much of the final organic "rain" which reaches the bottom consists of fecal pellets (Spencer et al, 1978). These are reworked biologically at the sediment interface, thereby losing their original morphologies.
Long residence of organic matter in the water colimm before sedimentation adversely affects its preservation (Degens and Mopper, 1976). Thus the depth of the water column as well as the size of organic particles affects the quantity and quality of the organic input to the sediments.
Influence ai Sediment-Particle Size In addition to being affected by bioturbation,
bacterial activity in recent sediments is also influenced by granulometry. Fine-grained sediments, where diffusibility of oxidizing agents is restrained, have lower levels of bacterial activity than coarse-grained sediments (Bordovsky, 1965), which helps explain the broad correlation between particle size and organic carbon content in modem and ancient sediments. Besides being deposited in high-energy environments, coarse clastic sediments, such as sands, permit easy diffusion of free oxygen and oxidizing salts dissolved in wa-

1186 G. J. Demaison and G. T. Moore
ter (Tissot and Welte, 1978, p. 81-91). They are always very low in organic matter.
Influence of Sedimentation Rate The rate of accumulation of organic carbon in
marine sediments is closely related to the bulk accumulation rate (Heath et al, 1977). This regularity is well demonstrated under oxic water in areas such as the Argentine basin (Stevenson and Cheng, 1972), the northwest African continental margin (Hartmann et al, 1976), and the deep regions of the North Pacific and North Atlantic Oceans (Heath et al, 1977). In all these areas there is a positive correlation between sedimentation rate and organic carbon content in sediments. The latter usually ranges between 0.3 and about 4%. Under anoxic water this correlation is less evident, as will be discussed later. The overall range of organic carbon under anoxic water in modem sediments is significantly wider and higher (about 1 to 20-1- %), regardless of sedimentation rate.
Furthermore, insufficient documentation exists for modem sediments with regard to possible dilution effects created by very high sedimentation rates (above 500 cm/lO' year). There is evidence that the positive correlation between organic carbon content and sedimentation rate at low and intermediate sedimentation rates may not extend indefinitely into the realm of very high rates. If it did, prodelta muds should be the richest organic sediments in the marine realm, yet organic carbon contents are systematically below 1% in modem prodelta sediments of the Louisiana Gulf Coast (Dow and Pearson, 1975), Niger delta (Klingebiel, 1976), and Amazon delta (Bezrukov et al, 1977) pointing to a dilution effect. Primary biologic productivities offshore from the Mississippi and Amazon deltas are exceptionally high (Fig. 1) but, as expected in such oxic environments, are not reflected by the sedimentary organic carbon content.
Sediments under anoxic water tend to be organically richer than those under oxic water, largely due to lack of benthonic scavenging and absence of bioturbation at the seafloor. Of even greater importance to the genesis of oil source beds, sedimentary organic matter is more reduced and lipid-rich under anoxic water than under oxic water. The cause for this preferential lipid enrichment probably relates to the biochemistry of anaerobic bacterial activity and requires further research.
Under oxygenated ("oxic") water, fluctuations in organic carbon content are clearly related to sedimentation rates (up to the point where dilution becomes significant), with little influence of surface productivity. Variations in surface pro
ductivity appear systematically, compensated by quantitative variations of the benthic biomass responsible for organic matter consumption and bioturbation at the seafloor. Under anoxic water, both sedimentation rates and perhaps surface productivity are factors which explain fluctuation in organic carbon content within the observed range. The respective inputs of these two factors, however, are yet to be satisfactorily elucidated.
DEVELOPMENT OF ANOXIC CONDITIONS IN WATER MASSES
Maximum oxygen saturation in sea water is about 6 to 8.5 ml/1, depending on water temperature and salinity. For this study we define as "anoxic" any water containing less than 0.5 milliliters of oxygen per liter of water (0.5 ml/1), which is the threshold below which the metazoan benthic biomass and, more specifically, bioturbation by deposit feeders, becomes significantly depressed. Therefore, it is proposed as the effective "biochemical fence" between potentially poor or good quaUtative and quantitative preservation of organic matter in sediments.
There are two "end member" causes to anoxia in natural waters: excessive oxygen demand and deficient oxygen supply. Anoxic conditions occur where the natural demand for oxygen in water exceeds the supply.
Oxygen Demand in Natural Water Oxygen consumption in water is essentially a
biochemical process resulting from the degradation of organic matter produced in the shallow layers of the euphotic zone. At least 80% of this oxidation occurs in these shallow layers and decreases sharply with depth. However, dead organic matter that has escaped total degradation and has sunk to the bottom continues to create a demand for oxygen which, however weak in relation to that prevailing in the euphotic zone, remains to be satisfied by a matching supply. If this continued demand is not replenished by deep-water circulation, the water colunm becomes anoxic.
Even water columns with a normal oxygen supply can become anoxic. This occurs in areas of very high primary productivity wherever the oxygen supply near sea bottom cannot cope with the load of descending dead organic matter.
Oxygen Supply in Natural Waters
Oxygen is supplied to water masses by two physical processes: (1) downward movement of oxygen-saturated water from the well-aerated surface layers (oxygen is supplied to the surface layers by exchange with the atmosphere and by pho-tosynthetic oxygen production) and (2) upward movement of oxygen-rich, colder, denser bottom

Anoxic Environments 1187
water into intermediate water zones. Three physical properties of water govern oxy
genation of bottom waters: water density increases with increasing salinity; water density increases with decreasing temperature (to 4°C); and oxygen solubility in water varies inversely with decreasing temperature and salinity.
If oxygen-rich surface water becomes denser because it is saltier or cooler than the surrounding water, it sinks to the bottom and circulates there as an aerating undercurrent and causes multilay-ered vertical stratification in water bodies of all sizes. The terms, thermocUne, pycnocline, and ha-locline, describe temperature, density, and salinity boundaries, respectively.
Stratification of seas and oceans in terms of oxygen concentration has long been recognized by oceanographers. Modem oceans would be entirely anoxic at depth without aeration of their basins by colder and denser oxygen-rich bottom water derived from the pwlar regions, mainly from the high southern latitudes.
The circulation of these deep oxygen-rich bottom waters is only partly known. Oxygen supply to the intermediate and deep oceanic water depends on patterns of water circulation due to surface-wind stress, density differences, high-latitude cooling, and the Coriolis force (caused by the earth's rotation). The dynamics of general oceanic circulation are complex, fluctuating, and not yet fully understood quantitatively.
The most common cause of anoxia is the incapacity of the oxygen supply in water to meet the biochemical oxygen demand. Hence lack of vertical mixing and oxygen renewal in deep waters is, perhaps, the most important factor controlling the location of anoxic layers and thus, indirectly, the preferred sites of deposition of oil source bed precursors. In the words of Wyrtki (1962), "Biochemical processes are responsible for the existence of oxygen minima, but circulation is responsible for the position."
CLASSIFICATION OF ANOXIC ENVIRONMENTS The world map of organic carbon (Fig. 2),
based mainly on Bezrukov et al (1977), does not distinguish the areas with organic carbon enrichment over 3%. Thus input of transported terrestrial organic matter as well as the effects of high sedimentation rates under oxic water cannot be separated from enrichment in marine organic matter created by anoxic conditions. In the writers' experience with ancient marine sediments, organic carbon contents rarely exceed 3% when transported humic material of terrestrial origin is predominant. Conversely, ancient marine sediments containing more than 3% organic carbon always contain a significant portion of aquatic or
ganic matter. Consequentiy, we reviewed in detail studies of
recent sediments reported by previous researchers to investigate the implications of enrichments higher than 3%, potentially caused by anoxic conditions. We found it possible to classify present-day anoxic environments into four main types: (1) large anoxic lakes; (2) anoxic silled basins; (3) anoxic layers caused by upwelhng; and (4) the anoxic open ocean.
LARGE ANOXIC LAKES Oxygen depletion in inland seas and large lakes
is determined by the supply-demand balance between free oxygen availability in bottom water and planktonic productivity of organic matter in the shallow layers.
Plant nutrients such as phosphates and nitrates are carried into lakes and inland seas by fluvial drainage systems that transport solutes leached from soils. These nutrients usually hmit the planktonic productivity of lakes, which then determines the amount of oxygen needed to recycle dead organic matter. Eutrophic lakes are characterized by an abundance of dissolved plant nutrients and by a seasonal oxygen deficiency in the bottom waters. Oligotrophic lakes are deficient in plant nutrients and contain abundant dissolved oxygen in their bottom water.
Oxygen supply in the bottom waters is usually good in areas of contrasting climate with seasonal overturning of the lake (Swain, 1970, p. 73-111). Also, cold, well-oxygenated stream and river water sinks to the bottom and enhances oxic conditions. Oxygen supply is lower in warm tropical climates because of lack of seasonal overturn and lower oxygen contents due to higher water temperatures.
Examples of Large Anoxic Lakes The two best studied anoxic lakes in the world
are Lake Kivu and Lake Tanganyika, both part of the East African rift-lake system.
Lake Tanganyika (Degens et al, 1971)—Lake Tanganyika, proposed as the type example for "large anoxic lakes" (Fig. 5), measures 650 km by up to 70 km. The maximum water depth is about 1,500 m and anoxic conditions lethal to metazoan hfe prevail below about 150 m. Some hydrogen sulfide is present in the anoxic water. Sediment cores from both deep and shallow water show considerable vertical variabiUty or varving.
Sediments deposited in the shallow aerated water within the lake contain 1 to 2% organic carbon, whereas the deep-water anoxic sediments range between 7 and 11% in organic carbon content. Carbon isotope ratios of recently deposited organic matter in anoxic sediments of Lake Tan-

1188 G. J. Demaison and G. T. Moore
LAKE TANGANYIKA NW SE
CH4,H^S PERMANENT THERMOCLINE
LONGITUDINAL SECTION 525 Km
Km
YJ5 1 15
ANOXIC SEP. ORG. CARB. 7 -11 % OXIC SEP. ORG. CARB. 1-2%
U = 5 - 50 cm /lOOOyrs
SIMILAR SETTINGS:
LAKE KIVU and DEAD SEA
FIG. 5—Type example of "large anoxic lake,' sedimentation rates in cm/1,000 years.
Lake Tanganyika. Symbol U in this and following figures expresses
ganyika are in a range normally associated with organic matter deposited in marine sediments: - 2 1 to - 2 2 %o (Pedee belemnite—PDB). Remains of diatoms are the most abundant fossils in these sediments.
Lake Kivu (Degens et al, 1973}—-Lake Kivu is nearly 500 m deep but is entirely anoxic below a depth of about 60 m. The pH of the surface water is around 9 but drops below 7 in the anoxic deep water where hydrogen sulfide is present in detectable quantities. The recent deep-water sediments under the anoxic zone contain up to 15% organic carbon.
The distribution of organic carbon in association with the stratigraphy of carbonate and anhydrite beds, in the past 6,000 years, reveals significant cUmatic fluctuations. Sapropel-rich beds coincide with humid climate and high water. Periods of dry climate, low water, and evaporites coincide with lower, but still significant, organic enrichments.
Examples of Large Oxic Lakes
Some East African rift lakes are oxic, notable examples being Lake Albert and Lake Victoria. Lake Albert (Tailing, 1963) is a thermally stratified lake 50 m deep. There are a few oxygen-depleted pockets down to 0.5 ml/1. The shallow depth and density currents of cooler, oxygenated water are responsible for good ventilation. Lake Victoria is large but shallow (about 100 m) and is affected by seasonal overturns. It is not anoxic (Swain, 1970).
All the very large temperate lakes of the north-em hemisphere are well oxygenated. Three examples are typical: (1) Lake Baikal, in Siberia, although the world's deepest lake (1,620 m), is well aerated and no anoxic conditions are present (Swain, 1970); (2) the Great Lakes of North America are oxic (Swain, 1970), although Lake Erie may be partly and intermittently anoxic because of excessive nutrient supply by pollution (phosphates from detergents and nitrates from sewage water); and (3) Great Slave Lake and other large lakes in western Canada are also well oxygenated (Swain, 1970).
Anoxic Enriroiuiients in Large Lake Systems
Tropical climate, with little seasonal change and moderate to high rainfall year around, promotes permanent water column stratification and therefore anoxia, particularly if water depth is in excess of 100 m. The sediments deposited are commonly both carbonate- and organic-rich. Organic carbon content is higher (commonly over 10%) than in anoxic marine sediments which may be caused by anaerobic degradation in fresh water being largely by fermentation rather than by the more efficient sulfate reduction process prevailing in anoxic marine environments.
Large anoxic lakes are not present in cold and temperate climates. Seasonal overturning of water, the higher capacity of cold water to dissolve oxygen, and density underflows of cold river waters all enhance oxic conditions in bottom lake waters of temperate and cold regions.

Anoxic Environments 1189
ETtdence of Large Anoxic Lakes fai Past Geologk Time A spectacular example of past oil shale and oil
source bed deposition in large anoxic lake systems is the Eocene Green River Formation in the western interior United States (mainly Colorado and Utah).
The Green River "oil shales" are brown, highly laminated, dolomitic marlstones containing hydrogen-rich kerogen (type I; Tissot et al, 1974). Retorting and conversion of this kerogen, during heat treatment of the whole rock, yields commercial shale oil. The Mahogany zone comprises the richest oil shales of the Green River Formation.
Several chemical and sedimentologic characteristics of these Mahogany zone "oil shales" discussed by Smith and Robb (1973) clearly demonstrate that permanent anoxic conditions existed at lake bottom during deposition. Smith and Robb's conclusions can be summarized as follows.
1. Evidence for complete lack of bioturbation is given by the absence of benthic fossils and the presence of minute seasonal laminations (varves) that can be followed laterally for tens of miles. Such features imply persistent lack of water movement and a lethal environment for benthic fauna in the bottom water.
2. The mineral composition of the "oil shale" implies a very alkaline, sodium carbonate-rich, bottom water whose higher density enhanced permanent chemical stratification of the lake. Eh-pH chemical fences (Krumbein and Garrels, 1952) implied by Mahogany "oil shale" mineral and chemical composition point to high pH (alkaline) and highly reducing (anoxic) conditions in the bottom water. These were lethal to macrohfe forms, thus explaining the lack of bioturbation and fine varving of the "oil shales." The upper surface layer of the lake above the density boundary was fully oxygenated and supported plank-tonic life, for organic matter was continually deposited in the sediment.
The lower part of the Green River Formation, below the Mahogany "oil shales," contains zones of finely laminated or varved, papery, kerogenous shales which usually assay less than 15 gallons per ton of rock (Roehler, 1974). These older anoxic shales differ from the Mahogany "oil shales" in that they were deposited in a freshwater lacustrine environment instead of an alkaline lake.
The lower part of the Green River Formation, mainly below the Mahogany zone "oil shales," has reached the principal stage of oil generation and is responsible for most of the crude oils produced in the Uinta basin (Tissot et al, 1978).
Crude oils generated from anoxic lake beds such as those present in the Green River Formation are highly paraffinic and typically feature
high pour points, when undergraded, and very low sulfur contents. The nonmarine Lower Cretaceous oils of China, Brazil, and some coastal west African basins belong in this category.
ANOXIC SILLED BASINS All silled basins have several physical barriers
(or sills) that restrict vertical mixing, thereby enhancing water stratification. Silling alone is not sufficient to create anoxic bottom conditions; certain patterns of water circulation, largely controlled by climate, need also to be present (Gras-shoff, 1975).
Basins with a positive water balance have a strong salinity contrast between fresher outflowing surface water and deeper ingoing more saline and nutrient-rich oceanic water. The development of permanently or intermittently anoxic conditions is a general feature of those semi-enclosed seas which have a positive water balance. Basins with a positive water balance also act as nutrient traps, thus enhancing both productivity and preservation of organic matter. Typical examples of anoxic landlocked silled basins with a positive water balance in a humid zone are: the Black Sea (Degens and Ross, 1974); the Baltic Sea (Grosshoff, 1975); Lake Maracaibo (Red-field, 1958); and Saanich Inlet, British Columbia (Nissenbaum et al, 1972).
In basins with a negative water balance, resulting from a hot, arid climate, there is a constant inflow of shallow oceanic water to compensate for high levels of evaporation. As shallow oceanic water enters, it replaces the hypersaline water which sinks and flows out as a density undercurrent into the ocean. Therefore, the basin bottom is both oxygenated and nutrient depleted (Fig. 6). Examples of oxic, negative-water-balance, silled basins without significant bottom organic enrichment are the Red Sea, Mediterranean Sea, and Persian Gulf.
Black Sea The Black Sea is the largest anoxic landlocked
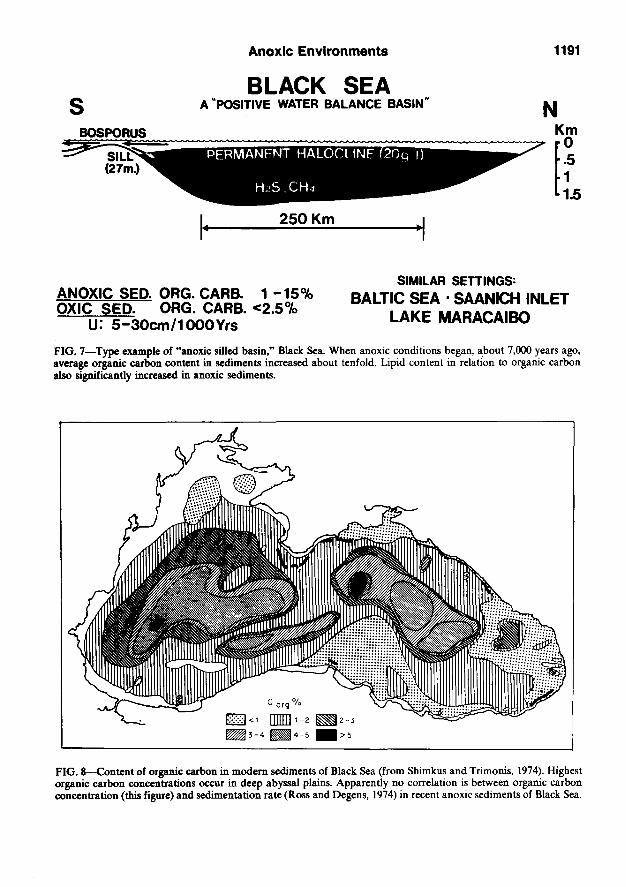
basin in the world. It has been studied in greater detail by oceanographers and geochemists than any other anoxic basin (Degens and Ross, 1974; Usher and Supko, 1978). We propose the Black Sea as the type example of an anoxic silled basin.
The Black Sea has a positive water balance with an excess outflow of fresh water resulting in relatively low salinity of surface water. As a result, a permanent halocline is present which also marks the boundary between oxic and anoxic conditions. Hydrogen sulfide is present in the anoxic zone. Its upper boundary is slightly convex, lying at a depth of about 250 m around the edges of the Black Sea and rising to about 150 m

1190 G. J. Demaison and G. T. Moore
EVAPORATION
OXYGENATED SALTIER DENSER WATER SINKS
DENSER WATER SPILLS OUT
LEAST OXYGENATED WATER
OXIC "NEGATIVE WATER BALANCE" BASIN EXAMPLES: MEDITERRANEAN - RED SEA - PERSIAN GULF
FIG. 6-Schematic model of oxic "negative water balance" basin. Hypersaline water flows out at depth over sill because it is denser than open oceanic water. This underflowing current keeps the basin bottom well ventilated and nutrient depleted.
in the center (Fig. 7). Below those depths the environment is lethal to all fish and invertebrate life.
About 22,000 years ago the Black Sea was a freshwater lake. Approximately 11,000 years ago rising Mediterranean waters began to invade the Black Sea as a consequence of climatic warm-up and ice retreat. About 7,000 years ago, the hydrogen sulfide zone began to form and present conditions were established about 3,000 years ago.
When anoxic conditions began about 7,000 years ago, the maximum organic carbon content of the sediments changed from 0.7 to 20% (Hirst, 1974). The organic-rich microlaminated black layer (7,000 to 3,000 years ago) is about 40 cm thick and is called "the old Black Sea" or "unit 2" (Degens and Ross, 1974). Unit 2 sediments commonly exceed 1% organic carbon.
Unit 1, between 3,000 years ago and the present, is about 30 cm thick and is composed of alternating white coccolith- and black organic-rich microlaminates (50 to 100 laminae per centimeter). Organic carbon content in unit 1 is still significant, but lower (about 1 to 6%) than in the underlying unit 2. In the two deepest basins, organic carbon contents in unit 1 range between 2 and over 5% (Fig. 8).
On the basis of varve chronology (Degens et al, 1978), rates of sedimentation in the deep basin were 10 cm/W years for the sapropel-rich unit 2 and 30 cm/lO' years for the more recent cocco-
lith-rich unit 1. Thus the organically richer sediments (unit 2) actually correspond to lower sedimentation rates. The correlation is inverse to that found in deep-sea sediments under oxygenated water by Heath et al (1977).
A sedimentation rate map for the whole Black Sea during the last 3,000 years (Ross and Degens, 1974) shows a possible overall negative correlation between sedimentation rate and organic matter concentrations (Shimkus and Trimonis, 1974). Sedimentation rates on the organic-lean upper slope of the Black Sea are higher by a factor of three than in the organically rich deep basins.
Further, Shimkus and Trimonis (1974) observed that "there is no correlation between phy-toplankton production and organic-matter content in sediments" of the Black Sea. "Fields of high organic-matter content correspond to areas of low primary production and vice-versa."
Recent geochemical investigations by Pelet and Debyser (1977) on a suite of Black Sea oxic unit 3 and anoxic unit 2 sediments showed that: (1) in both units organic matter is mixed, about % terrestrial plants and % marine plankton; and (2) the anoxic sediments (unit 2) have organic carbon contents from 3.85 to 14.95%, whereas the oxic sediments (unit 3) contain 0.65 to 0.69% organic carbon. The humic acid plus fulvic acid contents in relation to organic carbon are twice as high in oxic unit 3 than in anoxic unit 2. Hydrogen/car-
Anoxic Environments 1191
BLACK SEA A "POSITIVE WATER BALANCE BASIN" N
BOSPORUS
SILl? (27m.)
PERMANENT HALOC! INF f20q
H.'S.CH^
Km 0 .5
M ^J5
2 5 0 Km
ANOXIC SEP. ORG. GARB. 1 - 1 5 % OXIC SEP. ORG. GARB. <2 .5%
U: 5 - 3 0 c m / 1 0 0 0 Y r s
SIMILAR SETTINGS: BALTIC SEA • SAANICH INLET
LAKE MARACAIBO
FIG. 7—Type example of "anoxic silled basin," Black Sea. When anoxic conditions began, about 7,000 years ago, average organic carbon content in sediments increased about tenfold. Lipid content in relation to organic carbon also significantly increased in anoxic sediments.
FIG. 8—Content of organic carbon in modem sediments of Black Sea (from Shimkus and Trimonis, 1974). Highest organic carbon concentrations occur in deep abyssal plains. Apparently no correlation is between organic carbon concentration (this figure) and sedimentation rate (Ross and Degens, 1974) in recent anoxic sediments of Black Sea.

1192 G. J. Demaison and G. T. Moore
bon ratios of humic compounds are higher in sediments from anoxic unit 2 than in the oxic sediments of underlying unit 3. Significantly, the chloroform extractable lipid content, in relation to organic carbon, is five times higher in sediments from the anoxic unit 2 than in the oxic sediments of unit 3.
The organic carbon budget of the Black Sea has been estimated by Deuser (1971). He concluded that about 80% of the organic input is recycled in the top 200 m of water. The remainder falls into the anoxic hydrogen sulfide-poisoned water where approximately half of this organic material is further degraded by sulfate-reducing bacteria, which leaves about 5% of the original input for incorporation into the sediment and 5% solubilized in the anoxic water. Therefore, even in the most favorable model for organic-matter preservation (under anoxic water), 95% of the organic matter escapes fossilization and is eventually recycled.
BaMcSea
The Baltic Sea, the largest brackish water area in the world, has a positive water balance (Gras-shoff, 1975). It is affected by a permanent halo-cline with a pronounced dip to the east, from the Kattegat into the Gulf of Bothnia. A permanent oxygen deficiency exists below the halocline, with intermittent anoxia and hydrogen sulfide poisoning being present in the lower part of the water column in the Gothland Deep. Increase in bottom-water oxygen depletion during the last 75 years may be due partly to man-made pollution. Organic carbon maps are available for the Baltic Sea (Romankevich, 1977) and show a pattern of organic carbon enrichment above 3% coincident with the areas where the water column is anoxic.
Examples (rf Oxic Silled Basins
Two of the world's largest silled basins, the Red Sea and the Mediterranean Sea, are well oxygenated at depth and their modem sediments tend to be organic-poor. Both these basins are characterized by a negative water balance, that is, a larger inflow from the ocean than output to the ocean. The negative balance results from loss due to evaporation (Fig. 6).
In the Red Sea, oxygen-rich shallow water flows in from the Gulf of Aden and moves all the way up to the Gulf of Suez (Grasshoff, 1975). As it becomes saltier and denser, the water sinks and returns back at depth into the Gulf of Aden over the shallow sill of Bab el Mandeb. Localized oxygen depletion in the southern Red Sea is developed at the foot of the sill just before the dense, deep bottom water spills over southward into the Gulf of Aden. The northern end of the Red Sea,
farthest away from the sill, shows the highest levels of oxygenation in bottom water. Organic matter is low throughout the Red Sea, and sediments are largely biogenic clastic carbonates.
The Mediterranean is the world's largest landlocked silled marine basin, but bottom water is now entirely oxygenated at depth (Fairbridge, 1966). The lack of anoxic conditions in the Mediterranean Sea is due to a negative water balance circulation pattern identical to that observed in the Red Sea. There is, however, evidence from cores taken by DSDP and other oceanographic expeditions of past anoxic events in the eastern Mediterranean during the Pleistocene (Stanley, 1978). Intermittent anoxic conditions prevailed five times during the last 9,000 years. They are possibly the result of large and sudden influxes of fresh water, perhaps due to an increase in precipitations and/or ice retreat, which intermittently turned the eastern Mediterranean into a Black Sea-like, positive water balance basin.
Compared to oxidized Pleistocene sediments which contain less than 0.5% organic carbon, the anoxic muds of the upper Pleistocene of the east-em Mediterranean contain over 10 times more organic carbon (2 to 8%; Fairbridge, 1966). The latter are also laminated and devoid of benthic fauna.
The Persian Gulf is another silled basin with a negative water balance because of arid climate and intense evaporation. Bottom waters are oxygen-rich because of deep hypersaline water outflow and the highest organic carbon values measured (up to 2.5%) are in the silty clays that floor the deepest depressions in the southern part of the Persian Gulf (Hartmaim et al, 1971).
Baffin Bay, between Canada and Greenland, is a large silled marine basin where bottom water exhibits mild oxygen depletion (about 3 ml/1) but is nowhere near anoxic conditions. Dense, cold, oxygen-rich arctic water from the northern sill (Nares Strait) slowly sweeps the bottom in pulses and eventually spills out of Davis Strait, the southem sill, and into the Labrador Sea (Palfrey and Day, 1968).
Silled Depressioiis in Oxic Open Ocean
The world's oceans contain many silled depressions. They are well oxygenated except in rare examples like the Cariaco Trench and the Orca basin.
Cariaco Trench—The Cariaco Trench is a classic example of a relatively small local anoxic depression located on an oxygenated open ocean shelf (Richards and Vaccaro, 1956). This small silled depression, with a maximum depth of 1,400 m, is located on the continental margin north of Venezuela. It is anoxic below 300 m, the sill depth

Anoxic Environments 1193
being about 200 m. On the open oxygenated shelf, organic richness of recent sediments averages about 0.8% organic carbon. At the bottom of the trench, where anoxic conditions prevail, organic carbon concentrations increase as much as eight times in relation to the oxic environment, with values reaching 6% (Gormly and Sackett, 1977). Benthic foraminifers are lacking in the laminated anoxic sediments, but fish bones are very abundant. Radiocarbon measurements and studies of pelagic foraminifers indicate that stagnation of the trench was synchronous with an abrupt warming of surface water about 11,000 years ago (Richards, 1976).
Detailed geochemical investigations of the Car-iaco Trench anoxic sediments have been reported by Combaz and Pelet (1978) together with identical studies on sediments deposited under oxic water on the Demerara Abyssal Plain and on the outer deltaic cone of the Amazon River.
The anoxic sediments of the Cariaco Trench (1) are organically richer than their sedimentation rate (50 to 75 cm/10' years; Combaz and Pelet, 1978) would permit under oxic water; (2) contain insoluble organic matter that is hydrogen-rich, with kerogen in the "oil-prone" type II, category (the Amazon and Demerara kerogens and humic acids are less rich in hydrogen than their Cariaco Trench equivalents; they fit in the type III "gas-prone" category); and (3) contain larger amounts of hydrocarbons and other lipids, in relation to total organic carbon, than the oxic Amazon and Demerara sediments.
Lastly, investigation of fatty-acid content in the interstitial waters of the Cariaco, Demerara, and Amazon sediments, as compared to overlying sea-water (Saliot, 1977), shows preferential enrichment in fatty acids of the anoxic sediments in the Cariaco Trench.
Orca basin—The Orca basin is on the lower continental slope in the northwest Gulf of Mexico where complex bathymetry is largely attributed to salt diapirism and slumping of sediments. The basin is a crescent-shaped depression enclosing an area of approximately 400 sq km. The sill depth is approximately 1,900 m on the southeastern flank. The basin has two deeps slightly greater than 2,400 m at the north and south ends of the crescent.
The water in the Orca basin below a depth of 2,200 m, or about 200 m above the basin floor, is a brine. In addition to the pronounced increase in salinity at 2,200 m, there is a reversal of the temperature gradient, and an abrupt decUne in oxygen, from 5 ml/1 from a sample of 100 m above the halocline, to 0.0 ml/1 below the halocline.
Oxygen, as well as nitrate, is depleted by bacterial activity (Shokes et al, 1977). In all probabil
ity the brine in the Orca basin came from solution of a salt diapir at or near the surface somewhere around the basin slo(>e but the precise location of this hypothetical feature is not known.
Organic carbon in cored sediments under the anoxic water ranges between 0.8 and 2.9% (Sackett et al, 1978). Some of these values may be affected by dilution from slumped-in sediments from the fairly steep sides of the basin (Sackett et al, 1978).
EvMence of Anoxic Silled Basins in Past Geologic Time
Many anoxic silled basins are suspected in the geologic record but few are well documented in terms of integrated environmental and geochemical studies. Thus, in Great Britain, study of sedi-mentologic and fossil features in the Lower Jurassic (Toarcian) of Yorkshire by Morris (1979) permitted him to subdivide an apparently monotonous shale sequence into three facies (Fig. 9)— normal, restricted, and bituminous.
The normal shale is a homogeneous, bioturbat-ed sediment with abundant benthic body-fossils and common sideritic nodules. This facies is indicative of well-oxygenated bottom waters. The restricted shale consists of poorly laminated sediments with scattered calcareous concretions, sparse benthic fauna, and thin discrete pyritic burrows; this facies was deposited under oxygen-depleted water, perhaps close to 0.5 ml/1. The bituminous shale is a finely laminated sediment with pyritic concretions, little or no bioturbation, and a benthic fauna which is sparse and does not include burrowing organisms; this facies reflects anoxic conditions in the water column.
The three facies described by Morris (1979) occur in cycles and are indicative of variations in the position of the oxic-anoxic boundary within the water column (Fig. 9), replicating the tyjje settings described for modem sediments in Figures 3 and 4. We applied geochemical kerogen typing by Rock-Eval pyrolysis (Espitalie et al, 1977; Clem-entz et al, 1979) combined with organic carbon measurements on samples representative of the three facies, kindly supplied by the University of Reading (United Kingdom). The results are summarized on Figure 9; they confirm that the "normal shale" contains only "gas-prone" type III kerogen whereas the restricted shale is a mixed organic facies (type Ill-type II). Only the anoxic "bituminous" shale contains highly "oil-prone" type II kerogen. Widespread laminated oil shales in the Lower Jurassic (Toarcian) of the Paris basin as documented by Hue (1978) also resulted from anoxic basin conditions.
Alternating coccolith limestones, bioturbated clays, and laminated organic-rich oil shales de-

1194 G. J. Demaison and G. T. Moore
SHALE FACIES
AND KEROGEN TYPE
GEOCHEMICAL DATA
BIVALVE
GROUPS
TRACE
FOSSILS CONCRETIONS
ENVIRONMENTAL
INTERPRETATION
NORMAL SHALE
T Y P E I I I "GAS-PRONE"
KEROGEN
C ORG %:
0 6 6 - 3 . 4 0
HYDROGEN INDEX:
83-134
EPIFAUNAL a
I N F A U N A L
SUSPENSION,
INFAUNAL
DEPOSIT
ABUNDANT
CHONDRITES
HORIZONTAL
BURROWS
SIDERITIC
a CALCAREOUS
Oxic BoHom Woter
Oxidizing Conditions
teducing *" ' * —
%,« cing Conditions <sVOs<
RESTRICTED SHALE
TYPE n -nr " M I X E D "
K E R O G E N
C0RG7o'
2 .59-6 .75
HYDROGEN INDEX:
135-216
DOMINANT
INFAUNAL
DEPOSIT
FEW
UNBRANCHED
HORIZONTAL
BURROWS
CALCAREOUS W e a k l y O x i c Bot torn WQ t e r
BITUMINOUS S H A L E
TYPE n "OILPRONE"
K E R O G E N
CORG%:
5.61-11.42
HYDROGEN INDEX:
2 5 3 - 5 8 4
DOMINANT
EPIFAUNAL
SUSPENSION
NONE
NO BURROWS
PYRITIC
CALCAREOUS Anoxic Bottom Water.
FIG. 9—Interpretation of facies of Liassic (lower Toarcian) of Yorkshire, Great Britain. This format, excepting geocbemical data, was adapted from Morris (1979). Hydrogen Index, ratio expressing hydrogen richness of kerogen (Espitalie et al, 1977). Investigated rocks are all within same 60-m (197 ft) thick surface section. Degree of maturation of organic matter, as determined from Rock-Eval pyrolysis temperatures, is late immature to early mature.
posited during Late Jurassic time in the Kimmer-idgian of southern England (Dorset) show a strong analogy with recent Black Sea sediments. This association reflects cycUc oscillations of an oxic-anoxic boundary within a stratified water column (Tyson et al, 1979) during Late Jurassic time. Kinuneridgian to Volgian organic-rich anoxic shales are also present under parts of the North Sea. Where thermally mature, because of sufficient burial, they are the major source contributors to the North Sea giant oil reserves (Ziegler, 1979).
Anoxic, bituminous shales of very widespread areal extent were also deposited at approximately the same time (Volgian rather than Kimmeridgi-an) in the Western Siberian basin (Kontorovich, 1971). These Upper Jurassic anoxic shales, where thermally mature, are the source of most of the oil reserves entrapped in the many giant fields of this prolific petroleum province.
The best documented example of a former anoxic silled basin in North America is the Mow-ry sea which occupied in latest Albian time most of the northwestern interior United States. Byers and Larson (1979) described three distinct sedi-mentologic facies in the Mo wry Shale: (1) laminated mudstone, (2) bioturbated mudstone, and (3) bioturbated sandstone. The three facies represent the following environments: (1) a low-ener
gy, lethal environment, under anoxic water, where bioturbation is absent; (2) a low-energy environment where bioturbation is present because of sufficient oxygenation of the water column; and (3) a high-energy, intensely bioturbated and well-oxygenated environment. These were described by Byers and Larson under the Schaefer (1972) terminology of marine biotopes as (1) lethal isostrate, (2) vital isostrate, and (3) vital hete-rostrate.
The organically richest zones identified by Nixon (1973) as oil source beds of major significance coincide with the low-energy, unbioturbated, lethal facies, which was deposited under anoxic water in the Mowry sea.
ANOXIC LAYERS CAUSED BY UPWELLING
Upwelling is a process of vertical water motion in the sea wherever subsurface water rises toward the surface. Large-scale upwelling due to wind-stress-induced, Ekman transport occurs along certain coastUnes such as those of Cahfornia, Peru, Chile, South-West Africa, Morocco, and Western Australia. Upwelled water in coastal regions comes from relatively shallow depth, usually less than 200 m.
Ziegler et al (1979) described three distinct geographic situations where favorable conditions are met for coastal upwelUng: (1) meridional upwell-

Anoxic Environments 1195
S.W. AFRICAN SHELF
%o02
ANOXIC PATCHES EXTEND ABOUT 700 Km ALONG SHELF
Km 0 .5
• 1
•1.5 2
ANOXIC SEP. ORG.CARB. 3-26% OXIC SED. ORG. CARB. < 3 %
SilMILAR SETTiNG: PERU
FIG. 10—Type example of "anoxic layers caused by upwelling," South-West African Shelf (Benguela Current). Coastal upwellings are complex current and counter-current systems interacting with offshore winds. Upwelled water, rich in nutrients and low in oxygen, does not originate from great water depths, but from less than 200 m.
ing on north-south coasts between 10 and 40° lat. on the east side of oceans (e.g., California Current); (2) zonal upweUing on east-west coasts of equator-centered continents at about 15° lat. in association with the easterly trade winds (e.g., Caribbean Ciurent); and (3) monsoonal upwellings on diagonal east-facing coasts of equator-centered continents at about 15° lat. in association with easterly trade winds (e.g., Somalia Current).
Upwellings can also occur in the open ocean at water-mass boundaries, and such have been noted around the Antarctic continent, along the equator, and between Iceland and Norway. Upwelled water is rich in nutrients (nitrates and phosphates) and, therefore, promotes high biologic productivity. Recycling of dead organic matter in the water column creates a very high oxygen demand which can trigger anoxic conditions in deeper water layers under the upwelling. A classic example of anoxicity in the water column (essentially created by excessive biochemical oxygen demand due to coastal upwelling) occurs on the shelf offshore South-West Africa (Namibia) in association with the Benguela Current. We propose it as our type example (Fig. 10).
Benguela Cuirent
Considerable oceanographic and geochemical research has been conducted offshore of South-West Africa since the mid-1960s, notably by Calvert and Price (1971a, b). Anoxic conditions oc
cur on the South-West African Shelf (Fig. 10), particularly off Walvis Bay. The oxygen-depleted zone at sea bottom is an elongate area approximately 50 by 340 km (Calvert and Price, 1971a) parallel with and close to the coastline. Beyond the shelf break (about 100 km offshore) normal oxygenated conditions return to the bottom, as the regional oxygen-minimum layer in the Atlantic, south of Walvis Ridge, is only very weakly developed (Bubnov, 1966). Anoxic conditions are created on this narrow shelf by the high oxygen demand from decomposition of large amoimts of plankton resulting from the Benguela Current upwelling. The upwelling is due to a combination of a cold coastal current (the Benguela Current) and persistent offshore winds blowing northwest. Shallow surface water is skimmed off by the wind, permitting nutrient-rich subsurface water to ascend from a depth of about 200 m.
In three dimensions the system visualized is one where oxygen-poor, but nutrient-rich water constantly moves up and mixes in the euphotic zone with oxygenated water, causing high biologic productivity along a narrow coastal band. Dead plankton eventually falls to the bottom under the upwelling, and nutrients associated with the organic matter are brought back to the surface by the upweUing instead of being dispersed into the open sea. WTiat is buried and lost to the bottom sediments is replaced by nutrients brought in by the Benguela Current.
Brongersma-Sanders (1972) wrote: "Upwell-

1196 G. J. Demaison and G. T. Moore
ORGANIC CARBON CVJ
• /SYLVIA HILL
FIG. 11—Content of organic carbon in modem sediments offshore Walvis Bay (Namibia; from Calvert and Price, 1971). Highest values are off Walvis Bay and Cape Cross, where anoxic conditions and bacterial nitrate reduction intermittently prevail in bottom waters.
ings are fertile in the first place because nutrient-rich water is brought to the well-lighted surface layers, but this is not the only or even the main point, which is that upwelling is a kind of counter current system. A counter current system acts as a trap in which nutrients tend to accumulate."
Organic carbon concentrations in sediments under the oxygen-depleted zone range between 5 and 24% (Fig. 11). The highest values (over 20%) are off Walvis Bay and Cape Cross where oxygen-depleted conditions reach the anoxic level
(below 0.5 ml/1). Bacterial nitrate reduction occurs in bottom water. Free hydrogen sulfide has been detected in the water of Walvis Bay, causing occasional mass mortahty of fish and detectable hydrogen sulfide odor inland. There appears to be a clear positive correlation between level of oxygen depletion in the bottom water and organic carbon enrichment of the sediment.
Most organic matter is planktonic in origin because runoff from the land is intermittent and from a practically rainless desert (Calvert and Price, 1971b). Sedimentation rates were calculated by Veeh et al (1974) for anoxic sediments offshore Walvis Bay. Rates range between 29 and 103 cm/lO^ years. Organic carbon values range between 5.65 and 16.36% in the same samples and show no systematic correlation with sedimentation rates.
The anoxic, organic-rich sediments in the Ben-guela Current contain abnormal concentrations of copper, nickel, uranium, and phosphorus. Copper, molybdenum, and nickel distributions show similarities to that of organic carbon (Calvert and Price, 1971a). Uranium concentrations in sediments of the South-West African Shelf have been investigated by Veeh et al (1974). They reached the following conclusions: (1) the uranium has a positive correlation with organic carbon; (2) the uranium was derived from modem seawater; and (3) fixation of uranium in the sediments was conditioned by the presence of phosphorus together with reducing (anoxic) conditions; uranium is incorporated into apatite and is concentrated in phosphatic nodules and laminae.
When upwellings occur in areas of broad and well-developed intermediate oxygen-depleted layers, the oxygen starvation brou^t about by the local upwelling reinforces the regional oxygen anomaly. Notable coastal upwellings of this type have been investigated off California and Peru in association with regionally oxygen-depleted intermediate water layers.
Peru Current The Peru (Humboldt) Current is a well-docu
mented example of an anoxic environment associated with coastal upwelling (Fig. 12; Fairbridge, 1966). The Peru Current refers to a system of relatively shallow currents flowing northward along the west coast of South America. It is a complex system involving two surface currents, an undercurrent, and a countercurrent (Idyll, 1973).
The degree of upweUing offshore of Peru is related to the intensity of the wind stress. Prevailing trade winds off the coasts of northern Chile, Peru, and Ecuador blow principally from the northeast and south. This wind flow pushes the surface wa-

w
Anoxic Environments
PERUVIAN SHELF
1197
100 Km.
ANOXIC BAND ~XTENDS ALONG
HELP FOR OVER 1000 Km.
ANOXIC SEDL ORaCARR 3-11% OXIC SEP. ORG. CARa 05-3%
Km. rO .5
•1
•2
•3
L5
FIG. 12—Example of "anoxic layers caused by upwelling," Peru Current. Geochemical observations on sediments at bottom of Peru Shelf and Trench clearly demonstrate preferential preservation of lipids in those sediments under anoxic water.
ter northward at the same time that the CorioUs force deflects it to the west, thus skimming off the surface layers and letting cold subsurface water well up. Study of the isotherm patterns estabhshes that the depth of upwelling ranges from about 50 to 240 m (Fairbridge, 1966).
The upwelled water is undersaturated in oxygen but rich in nutrients (phosphates, nitrates). The ample supply of nutrients is associated with an exceedingly high rate of primary productivity. The combination of high productivity in the surface waters and depleted oxygen in the water column generates conditions favorable to enhanced preservation of organic matter in the underlying marine sediments. The concentration is further enhanced by the lack of significant water movement toward the ocean. TTie Peru Current, like the Benguela Current, is a self-regenerating nutrient trap and an outstanding primary producer of organic matter.
The organic matter in the bottom sediments of the Peruvian Shelf has been investigated by Ger-shanovich et al (1976). The highest concentrations of organic carbon (average 3.33% with values up to 11%) are in silty clays and laminated diatomaceous oozes under the anoxic zone at water depths between 100 and 500 m (Figs. 12, 13).
Organic-matter quality of its alkali-extractable portion was measured by elemental analysis. Hydrogen/carbon ratios of this humic material reach 1.37 to 1.43, indicating that they are not fulvic and humic acids identical to those of soils
or peats but are related to the "sapropelic acids" of Russian researchers. This type of marine humic material is considered as the main precursor of oil-prone kerogens. Hydrogen richness of kerogen precursor material is lower (H/C = 1.23) in the more oxygenated bottom sediments of the trench below 500 m. The same oxygenated sediments contain three times less extractable lipids in relation to organic carbon than those under the anoxic zone. Concurrently, the ratio of total lipid to total organic carbon of the bottom waters versus the sediments was investigated for the anoxic and oxic zones by Beliaeva and Romankevich (1976). Under oxygenated bottom waters in the trench, the soluble lipid to organic carbon ratio decreases by a factor of 22 from the water column into the sediments. On the anoxic shelf, however, the same ratio only decreases by a factor of 7. These observations clearly demonstrate preferential preservation of lipids in sediments under anoxic water.
The sediments of the Peruvian Shelf deposited under anoxic water contain significantly increased quantities of sapropel-type organic matter (70 to 90% of the organic matter); enhanced contents of soluble organic matter (bitumen or total lipids) and significant quantities of hydrocarbon gases including higher hydrocarbons up to hexane (Gershanovich et al, 1976) are also present. Uranium-enriched phosphorite nodules are present along the lower and upper boundaries of the oxygen-minimum layer at the edges of the

1198 G. J. Demaison and G. T. Moore
FIG. 13—Content of organic carbon in recent sediments offshore Peru (from Logvinenko and Romanke-vich, 1973).
anoxic zone (Veeh et al, 1973).
Examples of Upwellings WHhout Anoxic Lay«s
Not all areas of upwelling and high primary productivity are associated with anoxic layers in the water column and good concomitant organic-matter preservation in bottom sediments. Many areas of high primary productivity are underlain by highly oxygenated water which permits thorough aerobic degradation of the organic matter. Documented examples of such upwelling zones where the oxygen supply at sea bottom exceeds the biochemical oxygen demand are in Antarctica, the northern Pacific, and off southeastern Brazil.
In Antarctica, despite locally high surface pro
ductivity, the circumpolar waters are perhaps the most highly oxygenated on earth, largely because of very low surface temperatures (Fig. 14). Organic carbon values in bottom sediments around Antarctica reported in the Geological-Geophysical Atlas of the Indian Ocean (Udintsev, 1975) range between 0.10 and 0.94%.
In the northern Pacific, offshore Japan and the Kuriles, very high productivity caused by upwelling is offset by a constantly renewed supply of oxygen-rich intermediate and deep water coming from the Bering Sea.
Concerning offshore southeastern Brazil, Sum-merhayes et al (1976) concluded after deUiled study, "Beneath the biologically productive up-welled surface waters, there is remarkably httle sedimentation of organic matter. Also, although phosphorite deposits are usually associated with upwelhng centers, there are no phosphatic sediments off southeastern Brazil . . . this probably results from the high degree of oxygenation of upwelled water which contains 6 ml/1 of dissolved oxygen."
EvMence of Anoxic Layen Caused by Upwelling in Past Geologic Time
Paleogeographic reconstructions as applied to occurrence of Phanerozoic marine phosphorites (Ziegler et al, 1979) and petroleum source beds in the Paleozoic (Parrish et al, 1979) clearly point to the following.
1. Phanerozoic phosphorites were deposited in association with upwelling zones at low paleolati-tudes.
2. Phanerozoic phosphorites are not randomly distributed in time. Most deposits were formed in upwelling zones during periods of worldwide transgression and expansion of the oxygen-minimum layer.
3. Phosphorites and organic-rich sediments are commonly associated, as in the Permian Phos-phoria Formation (Parrish et al, 1979) in North America. The Phosphoria black shale members are major source contributors to Paleozoic oil accumulations in the western interior United States (Claypool et al, 1978; Momper and Williams, 1979), including several giant fields.
Evidence of anoxic sedimentation associated with coastal upwelling zones also is present in the Tertiary of California. Sedimentary suites characteristic of past deposition under the oxygen minimum layer of an upwelling are present at various levels in all the oil producing basins of Cahfomia. These sediments typically fit the following descriptions: laminated, phosphatic, bituminous, dark-brown shales, and laminated, hard, siliceous, dark-brown, commonly diatomaceous shales. Uranium-bearing phosphatic material is

oxyg
en m
l/1
• >1
.5
E
1.5-
0.5
• <0
.5
FIG
. 14
—M
axim
imi
exte
nsio
n of
oxy
gen-
depl
eted
lay
ers
in w
orld
oce
an.
Pref
eren
tial
dist
ribu
tion
of o
xyge
n-m
inim
um l
ayer
s in
ope
n oc
ean
is a
t lo
w la
titud
es a
nd o
n ea
ster
n si
des
of b
asin
s, la
rgel
y be
caus
e of
dee
p oc
eani
c ci
rcul
atio
n w
hich
is
cont
rolle
d by
glo
bal
clim
ate
and
Cor
ioU
s fo
rce.

1200 G. J. Demaison and G. T. Moore
abundant throughout, whether in nodules or finely disseminated, and radioactive background is relatively high. These rocks are generally devoid of macrofossils.
Anoxic organic-rich, bituminous, phosphatic shales are best developed in: the lower Pliocene (base of the Repetto Formation in the Los Angeles basin); the upper Miocene (lower part of die Puente Formation "nodular shale" in the Los Angeles basin, also the McLure and Antelope shales in the San Joaquin Valley); and the middle Miocene (major parts of the Monterey Formation in the Ventura basin, Santa Maria basin, and south-em Coast Ranges; middle Miocene, i.e., Luisian, time marks the peak of phosphatic deposition in the Tertiary of California; Dickert, 1966). The formations listed have been geologically recognized as prolific oil sources in the oil-producing California basins, long before the advent of modem petroleum geochemistry (Jenkins, 1943).
The most widespread anoxic sediments caused by past upwelling in California are in the middle Miocene (Luisian Stage) as well as in the lower part of the upper Miocene (Mohnian Stage). These stages, particularly the Mohnian, correspond with highest sea stands and maximum transgression during Miocene time on a global scale (Vail and Mitchum, 1979). Anoxic conditions associated with upwelling were also reinforced locally, in some places, by silled-basin topography. The overall oceanographic setting was clearly similar, however, to that present today in the Pera Current upwelling.
ANOXIC OPEN OCEAN
The intermediate layers of the open ocean both north and south of the equator in the eastern Pacific and northem Indian Ocean have been found to be oxygen depleted over areas of considerable size (Fig. 14). The volumes of anoxic water in today's open oceans greatly exceed the volumes of anoxic water present in the Black Sea.
It has been suggested that the distribution and position of these widespread oxygen-minimum layers result essentially from biochemical oxygen demand created by high planktonic productivity. Oceanographic observations on a global scale, however, do not agree with this hypothesis. Biochemical oxygen demand is indeed the triggering mechanism of anoxicity in general, but deep circulation patterns govern the distribution and position of these regional anomaUes.
The Grand Banks of Newfoundland in the northwest Atlantic form one of the world's great fishing grounds with productivities comparable to those offshore California or northwest Africa, yet no oxygen depletion of the intermediate or bottom water has been observed there. The overall
water column is well oxygenated (over 6 ml/1) and organic carbon in bottom sediments is present only in low concentrations, averaging 0.5% (Emery and Uchupi, 1972, p. 292, 371-373). The overall lack of correlation between areas of high oceanic productivity and oxygen depletion is also clearly demonstrated in the Pacific and Indian Oceans (Figs. 1, 14). The oxygen-depleted areas in these oceans are cul-de-sacs or are in the lee of sources of cold oxygenated bottom water. Oxygen concentrations decrease very gradually between Antarctica and the northem Indian Ocean and northeast Pacific (Reid, 1%5; Gorshkov, 1974) demonstrating very broad regional circulation effects, rather t^an local anomaUes related to over-productive coastal upwellings.
In only two important regions do substantial amounts of cold oxygenated surface water continuously sink to abyssal depth: the North Atlantic and around Antarctica. TTie deep oxygen concentration in the other oceans diminishes with increasing distance from these polar sources. The deep, cold, oxygen-rich bottom-water underflow from the poles counterbalances the evaporation from shallow waters in the subtropical regions of the oceans.
Strong bottom-current activity is prevalent in the cold, high-latitude regions and on the westem side of basins; these are the areas of most turbid bottom water (HoUister et al, 1978). Red pelagic clays also occur on the westem floors of oceanic basins (Fairbridge, 1966). These observations are coincident with better overall oxygenation of bottom and intermediate water at high latitudes and on the western sides of the Pacific and Atlantic Oceans (Fig. 14). This unequal latitudinal distribution of the regional oxygen-minimum layer (Fig. 14) indicates a tendency toward stagnation on the eastern sides of oceanic basins. It reflects, according to Stommel (1958), the effects of the Coriolis force upon deep circulation of oxygenated bottom water. It is clearly unrelated to areas of high primary productivity present on both sides of the same oceanic basins. Because this asymmetry is created by the earth's rotation, the same effect must have persisted throughout geologic time. Thus it could have had a profound influence on the paleogeographic distribution of oil source beds on continental shelves and slopes.
Significant organic enrichment has been observed around the world wherever the regional oxygen-minimum layer falls at or below 0.5 ml/l and intersects the continental slope and shelf. This correlation has been recognized and described in the northeastem Pacific Basin (Gross et al, 1972), in the Gulf of California (van Andel, 1964), and in the Indian Ocean (Stackelberg, 1972; Konjukhov, 1976).

Anoxic Environments 1201
INDIAN OCEAN
ANOXIC INTERMEDIATE LAYER (< 0.5)
r - ^ UPWELLING
SIIMILAR SETTING:
N.W. PACIFIC
CROZET BASIN
ARABIAN BASIN
4 mil/liter OXYGEN IN WATER
FIG. 15—Type example of "anoxic open ocean," Indian Ocean. Highest organic carbon concentrations are present wherever oxygen minimum layer faUs at or below 0.5 ml/1 and intersects continental slope and shelf.
Indian Ooean
The Indian Ocean is the type example of the "anoxic open ocean" of this classification.
The upper continental slope of the Indian Ocean from the Gulf of Aden to the Andaman Islands is occupied by a very large and relatively shallow anoxic layer marked in vertical hachures on Figure 15. Wherever this layer impinges on the shelf and slope between 250 and 1,200 m (Fig. 16), abnormally high organic carbon concentrations (between 2 and 10%) have been found by Stackelberg (1972), Konjukhov (1976), and other investigators (Udintsev, 1975). Organic carbon content on the shelf and other parts of the slope under oxic water is lower (between 0.5 and 1%). Similar observations were made in the Gulf of Oman (Hartmann et al, 1971).
Primary productivity varies from high, related to upwelling (marked in horizontal hachures on Fig. 15) on the west side of India, to low in the Gulf of Bengal. Sedimentation rates also vary widely from low, off the Arabian coast and in the Gulf of Oman, to very high off the Indus and Ganges deltas (Lisitzin, 1972, p. 135-171). The strongest correlation which overcomes differences in proiductivity and sedimentation rates is that between anoxic conditions and organic enrichment.
Pacific Ocean Offsiiote Waaidngton-Oiegon
In the northeast Pacific, offshore Washington and Oregon, a positive correlation is evident between organic carbon content of bottom sediments and oxygen depletion of bottom water (Gross et al, 1972). Where the dissolved-oxygen concentrations fall below 1 ml/1, organic carbon values range between 1 and 3%. Between 1 and 2 ml/1, organic carbon contents of bottom sediments fall to around 1%. Other factors such as grain size and sedimentation rate are significant, but it is statistically clear that the highest organic carbon concentrations occur where the oxygen-minimum layer impinges on the continental slope. In this general area, "there is surprisingly httle correlation between known areas of high primary productivity at the ocean surface and the amount of organic matter preserved in the sediment" (Gross et al, 1972).
Gulf of CaUfomia
The recent sediments of the Gulf of California (Mexico) have been investigated in detail by Calvert (1964), Parker (1964), Phleger (1964), and van Andel (1964) in relation to oxygen content of intermediate and deep waters (Roden, 1964) and primary productivity of surface water. Determi-

1202 G. J. Demaison and G. T. Moore
Water Depth ^2 1 Z 3 It 5mlA_
Mineral sond Relicts ond \ J I Vr^\,
Mica a R. ijiud X x V I C o ' - g . I Corb. jfecpell
tooo
2000
3oqo_
FIG. 16—Section through oxygen minimum layer on western shelf of India (modified from Stackelberg, 1972).
nations of organic carbon were made on approximately 100 core samples throughout this area. The organic carbon content appeared at first to correlate to the amount of clay in the sample. However, inspection of samples with comparable clay contents showed that the largest amounts of carbon (5% and above) are not in the silled basins within the gulf itself, but on the slopes of the southern gulf and in the open sea. In these areas the oxygen-minimum layer in the Pacific Ocean (below 0.5 ml/1) impinges on the slope between about 300 and 1,200 m. These areas of high organic enrichment and anoxic conditions in the water column coincide with finely laminated, richly diatomaceous sediments and the absence or scarcity of burrowing organisms. Conversely, lack of lamination due to homogenization by active bioturbation, as well as lower organic carbon content (below 2%), were demonstrated for those portions of the slopte where oxygen concentrations in the water column are higher than 0.5 ml/1 (Fig. 17).
Calvert (1964) also observed that oxygen concentrations lower than 0.5 ml/1 in the water col
umn did not completely inhibit the development of benthonic foraminifers but virtually suppressed the existence of macroinvertebrates and burrowing organisms.
Adandc Ocean
Although an oxygen-minimum layer has long been recognized here as well, the Atlantic Ocean in general tends to be oxygenated, for it is an open corridor between the two polar regions where cold, oxygen-rich bottom waters originate. There are only two notable oxygen-depleted intermediate zones, both in the eastern South Atlantic basins (Fig. 14). They are on the west side of Africa, on both sides of the equator, one off Cape Verde (Senegal) and one in the Angola basin (Bubnov, 1966). Only the Angola basin reaches the anoxic level (0.5 ml/l), but detailed geo-chemical observations on bottom sediments are lacking.
Offshore northwest Africa (Mauritania and Cape Verde), dissolved-oxygen concentrations in the oxygen-minimum layer are still in the oxic range (above 1 ml/1) whereas surface productivity

Anoxic Environments 1203
200-
400-
600'
^ 800
1000-X >-S I ZOO-
IE
5 1400-
1600'
1800'
2000
NUMBER OF CORES I 2 3 4 5 6 7 8 9 10 II 12 13 14 IS 16 I I I t I 1 I I 1 I I t I t I I
ZJ >.<iH(ni If 1 111 [I
H y H g i l l ' ••'• >f •• '
ifH'' t
OXYGEN CONCENTRATION ml/L 0.5 I 2 3 4 5 6
I I 1-
^ g LAMINATED m SEDIMENTS
f ' i .V'- .V
FIG. 17—Degree of bioturbation in relation to oxygen concentration in bottom water observed in Gulf of California (modified from Calvert, 1964). Laminated sediments are present at oxygen concentrations lower than 0.5 ml/1. This level of oxygen depletion, not total absence of oxygen, is effective threshold to lack of bioturbation.
is high owing to local upwelling. Organic carbon measurements by Gaskell et al (1975), Hartmann et al (1976), and Debyser et al (1977) demonstrate that organic preservation is variable in this oxic environment with a range of less than 0.5 to about 4% organic carbon. These organic carbon fluctuations under oxic water are controlled by sedimentation rates and do not reflect present high productivities created by upwelling conditions (Hartmann et al, 1976).
Offshore Ivory Coast and Ghana, oxygen depletion is also present in the intermediate water (about 1.5 ml/1), but does not reach the anoxic level. Organic carbon values range between 3 and 5% but organic matter is largely composed of transported terrestrial plant remains (Klingebiel, 1976). The predominance of transported terrestrial organic matter, originating from rain forests, into an open-marine environment under 1,200 m of water, is an interesting present-day example of a situation common in ancient open-marine sediments. Such a setting implies abundant rainfall on nearby continental masses.
In the Niger delta, in nearly identical climatic surroundings, organic carbon content in prodelta clays is low and ranges between 0.5 and 1.0% (Klingebiel, 1976). Sedimentation rates are very
high, suggesting that dilution of organic matter must occur there.
Evidence of Widespread Anoxic Open-Ocean Conditions in Past Geologic Time
Oceanographic observations are made today on a cold earth. Present time represents a relatively brief oscillation within the Pleistocene Ice Age sequence. The modern oceans and seas are relatively well oxygenated at depth because contrasted climatic belts and polar ice caps enhance vigorous convective water mass movements (Berggren and HoUister, 1977). The active bottom circulation that characterizes the present Atlantic, for example, was not initiated until about 3 m.y. ago. Q)ld-earth conditions as we know them today were heralded by major cUmatic cooling during late Eocene-Oligocene time followed by late Miocene and Phocene-Pleistocene global marine regressions and glaciations. The long-term trend since Eocene time is one of global climatic cooling concurrent with a lowering of the mean sea level in the world ocean (Vail and Mitchum, 1979). In the geologic past, deep oceanic circulation was not so active during periods of warm equable global cUmate which corresponded with high mean sea levels and the absence or extreme

1204 G. J. Demaison and G. T. Moore
reduction of polar ice caps. For example, sedimentary and paleontologic data interpreted within the framework of plate tectonics suggest that the Arctic Ocean and north-polar regions were ice-free in Late Jurassic (Hallam, 1975, p. 52-63) and Late Cretaceous time (Clark, 1977).
Evidence of worldwide oceanic anoxic events (Schlanger and Jenkyns, 1976) in the generally warm Mesozoic Era is indicated by remarkably widespread organic-rich black shales during the Late Jurassic and middle Cretaceous. The global extent and synchroneity of the Cretaceous anoxic events have been shown by the Deep Sea Drilling Project (Degens and Stoffers, 1976; Schlanger and Jenkyns, 1976; Ryan and Cita, 1977; Thiede and van Andel, 1977; Tissot et al, 1979). These paleoanoxic events have also been increasingly recognized on the continental shelves by petroleum exploration drilling.
Identical worldwide oceanic anoxic events are inferred also to have prevailed intermittently in Paleozoic time because the widespread Lower Or-dovician, Silurian, and lower Carboniferous organic-rich marine black shales coincide with times of widespread marine transgression and ice cap melting (Berry and Wilde, 1978).
Lastly, fiiere is statistical evidence that the known oil source bed systems present in the stratigraphic record are not randomly distributed in time but coincide with periods of worldwide transgression and oceanic anoxia (Arthur and Schlanger, 1979; Tissot, 1979). Prolific petroleum source beds deposited over part of the Jurassic and Cretaceous account for 70% of the oil in 148 investigated petroleum zones, which, themselves, total about 85% of the world's reserves (Tissot, 1979). Yet this privileged period for oil source bed deposition (Jurassic-Cretaceous) only amoimts to 17% of Phanerozoic time. Prolific petroleum regions where parts of the Cretaceous and/or Jurassic play a major oil source role are the Middle East (Kent and Warman, 1972, p. 150), the North Sea (Ziegler, 1979), the West Siberian basin (Kon-torovich, 1971), and Mexico (Arthur and Schlanger, 1979).
CONCLUSIONS The explosive growth in knowledge by recent
observations in oceanography, geochemistry, and geomicrobiology has made it necessary to review many traditional concepts relating to oil source bed genesis. The proposed revised concepts follow.
1. Potential oil source beds are organic-rich sediments containing a kerogen type (type I or II) that is sufficiently hydrogen-rich to convert mainly to oil during thermal maturation.
2. Measurement of organic carbon content, alone, is insufficient to identify potential oil source beds. For example, "gas-prone" transported terrestrial organic matter (OM), oxidized aquatic OM deposited at high sedimentation rates, or reworked OM from a previous sedimentary cycle commonly create misleadingly high levels of organic carbon in ancient marine sediments. High organic carbon content does not necessarily equate with oil source potential, hence precise measurement of kerogen type is indispensable to the identification of oil source beds. Insight into the early environmental factors that determine kerogen type, furthermore, is the principal key to understanding the temporal as well as paleogeographic distribution of oil source beds.
2. Organic matter in sediments below anoxic water (defined as water containing dissolved oxygen in amounts less than 0.5 ml/1) is commonly more reduced (hydrogen-rich), more lipid-rich, and more abundant than under oxic water.
3. The organic residues of anaerobic bacterial degradation are more reduced (hydrogen-rich) and more lipid-rich than those resulting from oxidation by aerobic microbes. Hence anaerobic residues are, potentially, the precursors to "oil-prone" kerogens. The latter are essential constituents of potential oil source beds. The mechanisms leading to this preferential enrichment in lipids are little understood and in need of fundamental research in microbial biochemistry.
4. The overall range of organic carbon content under anoxic water is significantly wider and higher (about 1% to over 20% organic carbon) than that in sediments deposited under oxygenated water. It is suggested that variations in sedimentation rate, if perhaps one of the underlying reasons for organic richness fluctuations under anoxic water, are not the main cause of enhanced preservation in reducing environments. Sediments deposited under anoxic marine wato- also contain abnormally high concentrations of U, Cu, Mo, Ni, P, and S which tend to correlate positively with organic carbon concentrations. Organic matter deposited under anoxic water, wherever investigated, is hydrogen-rich and therefore "oil-prone."
5. Organic carbon content in sediments under oxygenated water is primarily a function of sedimentation rates up to the point (yet ill-defined) where organic matter dilution significantly depresses concentrations. Surface primary productivity appears to have little influence on organic carbon concentrations in sediments under "oxic" water because it is systematically compensated by organic matter consumption at the seafloor by benthic organisms. Organic carbon contents un-

Anoxic Environments 1205
der "oxic" water typically range between less than 0.5% to maxima between 3 and 4%, regardless of surface productivities. Organic matter deposited under oxic water, wherever investigated, is hydrogen depleted and therefore "gas prone."
6. Bioturbation, or biologic sediment mixing by metazoans, is ubiquitous at all water depths under "oxic" water. It causes acceleration of geo-chemical processes at and below the sediment-water interface. Added to this effect is the grazing of the sediment bacterial biomass by the mobile deposit feeders (burrowers) responsible for bioturbation. Bioturbated sediments under "oxic" water are homogenized whereas sediments deposited under anoxic water are varved or laminated and devoid of burrowing, deposit-feeding infau-na.
7. Maximum oxygen saturation in seawater is about 6 to 8.5 ml/1, but benthic metazoans are unaffected by lower oxygen concentrations, down to about 1 ml/1 of water. Between 0.1 and 0.5 ml/l, a low diversity, highly stressed, suspension-feeding (non-burrowing) epifauna can still survive above the sediment-water interface. Below 0.5 to 0.3 ml/1, bioturbation by deposit feeders (burrowers) virtually ceases and the overall benthic biomass is sharply reduced. Below 0.1 ml/1, all metazoans disappear leaving anaerobic bacteria as the only effective reworkers. Hence 0.5 ml/1 (the threshold of arrested bioturbation), not total absence of oxygen in water, is proposed as the effective "biochemical fence" between potentially good or poor quahtative and quantitative organic matter preservation.
8. The tolerance of protozoans, such as benthic foraminifers, to very low oxygen concentrations appears to be higher than that of metazoans. The ecology of benthic foraminifers under oxygen-depleted water is clearly in need of fundamental research.
9. Anoxic water is defined, for this study, as water containing less than 0.5 miUiliters oxygen per hter of water. Anoxic conditions occur where the natural demand for oxygen in water exceeds the supply. Oxygen demand relates to surface primary productivity, whereas oxygen supply below the depth of surface mixing depends on water circulation.
10. Lakes are rarely permanently anoxic at depth. It takes special conditions such as overfer-tilization, lack of overturning of the water in warm climates, and preferably, but not necessarily, deep water to create stable anoxic conditions. When they occur, as in Lake Tanganyika (east Africa), sediments rich in organic matter are deposited. Warm paleoclimatic conditions with lack of seasonal contrast and moderate rainfall should
be most favorable to lacustrine oil source bed deposition.
11. At present, the majority of silled basins, whether landlocked or on open shelves, are not anoxic and thus do not favor deposition of oil source bed precursor sediments. Only those silled basins that have a water circulation pattern implying a "positive water balance" become anoxic, for example, the Black Sea or the Baltic Sea. The degree of ventilation in silled depressions on open oceanic shelves follows that one prevailing at comparable depth in the adjoining open oceanic water masses. Hence silled-basin geometry in ancient sedimentary basins does not automatically imply the presence of oil source beds. Silled basins should be prone to anoxia and oil source bed deposition at times of worldwide transgression, both at high and low paleolatitudes. At times of worldwide regression, those landlocked basins in temperate paleoclimatic belts with high rainfall should be more favorable than those in arid zones.
12. Coastal upweUings, which are commonly associated with high primary productivity of the surface layers, do not originate from great water depths. Only upwellings associated with anoxic layers show any abnormal organic preservation as well as phosphorite deposition in bottom sediments (e.g., Benguela and Peru Currents). Extensive oil source beds and phosphorites resulting from upwelling conditions should be sought at low paleolatitudes and preferably at times of worldwide transgression.
13. Large volumes of anoxic water are present in the oxygen-minimum layers of open oceans such as the Indian Ocean without silling or physical barriers. Significant organic enrichment in sediments has been found wherever the oxygen-minimum layer reaches the anoxic level and intersects the continental slope and shelf. The preferential distribution of oxygen-minimum layers in the world ocean is at low latitudes and on the western side of continents, primarily because of deep oceanic circulation spurred and controlled by climatic contrast of the poles and the equator and the Coriolis force. Inferred are similar global patterns, determining the position of oxygen-minimum layers during the geologic past, but with much more widespread extensions of anoxia invading whole oceanic basins. This is suggested by remarkably widespread organic-rich marine black shales at times of global climatic warm-ups and major transgressions as in Late Jurassic and middle Cretaceous time. Known marine oil source bed systems in the stratigraphic record are not randomly distributed in time but coincide with periods of worldwide transgression and oceanic

1206 G. J. Demaison and G. T. Moore
ANOXIC BASIN TYPE
ANOXIC LAKES
ANOXIC SILLED BASINS
ANOXIC LAYERS W/ UPWELLINGS
ANOXIC OPEN OCEAN
PALE0GE06RAPHIC SETTING
EQUABLE. WARM. RAINY. EARLY RIFTS. INTERMOUNTAIN BASINS.
TEMPERATE TO WARM. RAINY. INTRACRATONIC SEAS. ALSO ANOXIC POCKETS ON SHELVES.
OCEANIC SHELVES AT LOW LATITUDES. WEST SIDE OF CONTINENTS.
BEST DEVELOPED AT TIMES OF GLOBAL WARM-UPS & MAJOR TRANSGRESSIONS.
STRATI6RAPHIC DISTRIBUTION OF ANOXIC SEDIMENTS
CONTINUOUS WITHIN SAME ANOXIC LAKE SYSTEM.
VARIABLE. TENDS TO BE RICHEST AT BOTTOM OF BASIN OR POCKET.
OFTEN NARROW TRENDS. BUT CAN BE WIDESPREAD. PHOSPHORITES. DIATOMITES.
VERY WIDESPREAD W/LITTLE VARIATION. OFTEN SYNCHRONOUS WORLDWIDE.
FIG. 18—Classification of anoxic depositional models. Certain settings in marine realm may combine two or even three anoxic basin types, for example, local anoxic layers caused by upwelling may reinforce anoxic open-ocean oxygen-minimum layer impinging upon shelf with silled depressions.
anoxia. 14. Geochemistry now provides the means to
identify paleoanoxic events in the stratigraphic record. Paleogeographic recognition of the proposed anoxic models—large anoxic lakes, anoxic silled basins, anoxic layers caused by upweUing, and anoxic open ocean—in ancient sedimentary basins should help geologists and geochemists in the regional mapping of oil shales and oil source beds as shown in Figure 18.
REFERENCES CITED
Aller, R. C, 1978, The effects of animal-sediment interactions on geochemical processes near the sediment-water interface, in Estuarine interactions: New York, Academic Press, p. 157-172.
Arthur, M. A., and S. O. Schlanger, 1979, Cretaceous "oceanic anoxic events" as causal factors in development of reef-reservoired giant oil fields: AAPG BuU., V. 63, p. 870-885.
BeUaeva, A. N., and E. A. Romankevich, 1976, Chemical conversion of lipids during oceanic sedimentogen-esis, in N. B. Vassoevich, ed., Issledovania Organish-eskogo Veshchestva Sovremenyk Iskopaemyk Osadkov: Moscow, Adka. Nauk SSSR, p. 81-102 (in Russian).
Berggren, W. A., and C. D. HoUister, 1977, Plate tectonics and paleo-circulations—commotion in the ocean: Tectonophysics, v. 38, p. 11-4S.
Berry, W. B. N., and P. Wilde, 1978, Progressive ventilation of the oceans: Am. Jour. Sci., v. 278, p. 257-275.
Bezrukov, P. L., et al, 1977, Organic carbon in the upper sedimentary layer of the world ocean: Oceanology, v. 17, no. 5, p. 561-564.
Bordovsky, O. K., 1%5, Accumulation and transformation of organic substance in marine sediments: Ma
rine Geology, v. 3, p. 3-114. Brongersma-Sanders, M., 1966, The fertiUty of the sea
and its bearing on the origin of oil: Advancement of Science, v. 23, p. 41-46.
1972, Hydrological conditions leading to the development of bituminous sediments in the pre-evapo-rite phase, in Geology of saline deposits: Unesco Earth Sci. Ser., no. 7, p. 19-21.
Bubnov, V. A., 1966, The distribution pattern of minimum oxygen concentrations in the Atlantic: Oceanology, V. 6, p. 193-201.
Byers, C. W., and D. W. Larson, 1979, Paleoenviron-ments of Mowry Shale (Lower Cretaceous), western and central Wyoming: AAPG Bull., v. 63, p. 359-361.
Calvert, S. E., 1964, Factors affecting distribution of laminated diatomaceous sediments in Gulf of CaU-fomia, in Marine geology of the Gulf of CaUfomia: AAPGMem. 3, p. 311-330.
1976, The mineralogy and geochemistry of near-shore sediments, in J. P. Riley et al, eds.. Chemical oceanography, v. 6, 2d ed.: New York, Academic Press, p. 187-280.
' and N. B. Price, 1971a, Upwelling and nutrient regeneration in the Benguela Current, October, 1968: Deep-Sea Research, v. 13, p. 505-523.
1971b, Recent sediments of the Southwest African Shelf, in Atlantic continental margins: London, Inst. Geol. Sci., p. 175-185.
Clark, D. L., 1977, Climatic factors of the late Mesozoic and Cenozoic Arctic Ocean, in Polar Oceans: Calgary, Alberta, Arctic Inst. North America, p. 603-615.
Claypool, G. E., and I. R. Kaplan, 1974, The origin and distribution of methane in marine sediments, in I. R. Kaplan, ed.. Natural gases in marine sediments: New York, Plenum Press, p. 99-140.
A. H. Love, and E. K. Maughan, 1978, Organic geochemistry, incipient metamorphism, and oil generation in black shale members of Phosphoria Forma-

Anoxic Environments 1207
tion. Western Interior United States: AAPG Bull., v. 62, p. 98-120.
Clementz, D. M., G. J. Demaison, and A. R. Daly, 1979, Well site geochemistry by programmed pyroly-sis: Offshore Technology Conf., v. 1, p. 465-470.
Combaz, A., and R. Pelet, eds., 1978, Geochimie orga-nique des sediments marins profonds, ORGON II, Atlantique-N.E. Bresil: Paris, Editions CNRS, 456 p.
E>ebyser, Y., R. Pelet, and M. Dastillung, 1977, Geochimie organique des sediments marins recents, Mer Noire, Baltique, Atlantique (Mauritanie), in Advances in organic geochemistry, 1975: Madrid, ENA-DIMSA, p. 289-320.
Degens, E. T., 1974, Cellular processes in Black Sea sediments, in The Black Sea: AAPG Mem. 20, p. 296-307.
and K. Mopper, 1976, Factors controlling the distribution and early diagenesis of organic material in marine sediments, in J. P. Riley et al, eds.. Chemical oceanography, v. 6, 2d ed.: New York, Academic Press, p. 59-113.
and D. A. Ross, eds., 1974, The Black S e a -geology, chemistry and biology: AAPG Mem. 20, 633 P-
and P. Stoffers, 1976, Stratified waters as a key to the past: Nature, v. 263, p. 22-27.
R. P. Von Herzen, and H.-K. Wong, 1971, Lake Tanganyika; water chemistry, sediments, geological structure: Naturwissenschaften, v. 58, p. 224-291.
- et al, 1973, Lake Kivu; structure, chemistry and biology of an East African rift lake: Geol. Rundschau, v. 62, p. 245-277.
- et al, 1978, Varve chronology: estimated rates of sedimentation in the Black Sea deep basin: Initial Rept. Deep Sea Drilling Project, v. 42, pt. 2, p. 499-508.
Demaison, G. J., and M. Shibaoka, 1975, Contribution to the study of hydrocarbon generation from hydrogen-poor kerogens: 9th World Petroleum Cong., Tokyo, Proc., V. 2, p. 195-197.
Denser, W. G., 1971, Organic-carbon budget of the Black Sea: Deep-Sea Research, v. 18, p. 995-1004.
1975, Reducing environments, in J. P. Riley and R. Chester, eds.. Chemical oceanography, v. 3: New York, Academic Press, p. 1-37.
Dickert, P. F., 1966, Neogene phosphatic facies in California: PhD thesis, Stanford Univ.
Didyk, B. M., et al, 1978, Organic geochemical indicators of paleoenvironmental conditions of sedimentation: Nature, v. 272, p. 216-222.
Dow, W. G., 1977, Kerogen studies and geological interpretations: Jour. Geochem. Exploration, v. 7, p. 79-99.
and D. B. Pearson, 1975, Organic matter in Gulf Coast sediments: 7th Offshore Technology Conf., Proc., v. 3, OTC 2343, p. 85-94.
Emery, K. O., and E. Uchupi, 1972, Western North Atlantic Ocean: topography, rocks, structures, water, life, and sediments: AAPG Mem. 17, 532 p.
Espitalie, J., et al, 1977, Source rock characterization method for petroleum exploration: Offshore Technology Conf. Proc., v. 3, OTC 2935, p. 439-444.
Fairbridge, R. W., ed., 1966, The encyclopedia of oceanography: New York, Van Nostrand-Reinhold,
1021 p. Foree, E. G., and P. L. McCarty, 1970, Anaerobic de
composition of algae: Environ. Sci. Technology, v. 4, p. 842-849.
Gaskell, S. J., et al, 1975, The geochemistry of a recent marine sediment off northwest Africa. An assessment of source of input and early diagenesis: Deep-Sea Research, V. 22, p. 777-789.
Gerschanovich, D. E., V. V. Veber, and A. E. Konjuk-hov, 1976, Organic matter in the bottom sediments of the Peru region of the Pacific Ocean, in N. B. Vassoe-vich, ed., Issledovania Organisheskogo Sovremenyk Iskopaemyk Osadkov: Moscow, Akad. Nauk, SSSR, p. 121-128 (in Russian).
Gormly, J. R., and W. M. Sackett, 1977, Carbon isotope evidence for the maturation of marine lipids, in Advances in organic geochemistry, 1975: Madrid, ENA-DIMSA, p. 321-339.
Gorshkov, S. G., 1974, World ocean atlas, v. 1, Pacific Ocean: New York, Pergamon Press, 338 p. (in Russian).
1979, World ocean atlas, v. 2, Atlantic and Indian Oceans: New York Pergamon Press, 350 p. (in Russian).
Grasshoff, K., 1975, The hydrochemistry of landlocked basins and fjords, in J. P. Riley and J. Skirrow, eds.. Chemical oceanography, v. 2, 2d ed.: New York, Academic Press, p. 456-597.
Gross, G. M., et al, 1972, Distribution of organic carbon in surface sediment, northeast Pacific Ocean, in The Columbia River Estuary and adjacent ocean waters: Seattle, Univ. Washington Press, p. 254-264.
Hallam, A., 1975, Jurassic environments: Cambridge, U.K., Cambridge Univ. Press, 269 p.
Hartmann, M., et al, 1971: Oberflachensedimente im Persischen Gulf und Gulf von Oman: "Meteor" For-schungsergebnisse, Reihe C, No. 4, 76 p.
et al, 1976, Chemistry of late Quaternary sediments and their interstitial waters from the NW African continental margin: "Meteor" Forschungaser-gebnisse, Reihe C, No. 24, p. 1-67.
Heath, G. R., T. C. Moore, Jr., and J. P. Dauphin, 1977, Organic carbon in deep-sea sediments, in R. N. Andersen, ed., The fate of fossil fuel CO2 in the oceans: New York, Plenum Press, p. 627-639.
Hirst, D. M., 1974, Geochemistry of sediments from eleven Black Sea cores, in The Black Sea: AAPG Mem. 20, p. 430455.
HoUister, C. D., R. Flood, and I. W. McCave, 1978, Geological aspects of the benthic boundary layer: Oceanus, v. 21, p. 5-13.
Hue, A-Y., 1978, Geochimie organique des schistes bi-tumineux du Toarcian du Bassin de Paris: PhD thesis, Univ. Louis Pasteur, Strasbourg, France.
Hunt, J. M., 1979, Petroleum geochemistry and geology: San Francisco, W. H. Freeman and Co., 498 p.
Idyll, C. P., 1973, The anchovy crisis, reprinted 1977, in Ocean science; readings from Scientific American: San Francisco, W. H. Freeman and Co., p. 223-230.
Jenkins, O. P., ed., 1943, Geologic formations and economic development of the oil and gas fields of CaU-fomia: California Div. Mines Bull. 118, 778 p.
Jones, G. E., 1973, An ecological survey of open ocean and estuarine microbial populations. I, TTie impor-

1208 G. J. Demaison and G. T. Moore
tance of trace metal ions to microorganisms in the sea, in L. H. Stevenson and R. R. Colwell, eds., Es-tuarine microbial ecology: Columbia, S.C, Univ. South Carolina Press, p. 233-241.
Kent, P. E., and H. R. Warman, 1972, An environmental review of the world's richest oil-bearing region; the Middle East: 24th Intemat. Geol. Cong., Montreal, Proc., sec. 5, p. 142-152.
Klingebiel, A., 1976, Sediments et milieuz sedimentaires dans le Golfe du Benin: Centre Recherche Pau-SNPA, Bull. 10, p. 129-148.
Koblenz-Mishke, O. I., V. V. Volkonsky, and J. G. Ka-banova, 1970, Plankton primary production of the world ocean, in W. S. Wooster, ed.. Symposium on scientific exploration of the southern Pacific, Scripps Inst Oceanog., 1968, 9th Mtg. SCOR: Washington, D.C., Natl. Acad. Sci., p. 183-193.
Konjukhov, A. E., 1976, Fades characteristics of contemporary sediments from the western submarine margin of the Indian Peninsula, in N. B. Vassoevich, ed., Issledovania Organisheskogo Veshchestva Sovre-menyk Iskopaemyk Osadkov: Moscow, Akad. Nauk SSSR, p. 111-120 (in Russian).
Kontorovich, A. E., ed., 1971, Geochemistry of petroleum bearing Mesozoic formations in the Siberian basins: USSR Ministry of Geology Publication, SNIIG-IMS Bull. 118, 85 p. (in Russian).
Krumbein, W. C , and R. M. Garrels, 1952, Origin and classification of chemical sediments in terms of pH and oxidation-reduction potentials: Jour. Geology, v. 60, p. 1-33.
Lijmbach, G. W. M., 1975, On the origin of petroleum: 9th World Petroleum Cong., Tokyo, Proc, v. 2, p. 357-368.
Lisitzin, A. P., 1972, Sedimentation in the world ocean: SEPM Spec. Pub. 17, 218 p.
Logvinenko, N. V., and Ye A. Romankevich, 1973, Recent sediments of the Pacific Ocean on the coast of Peru and Chile: Lithology and Mineral Resources, v. 8, p. 1-11; also in Litol. Polez. Iskop., no. 1, p. 3-16, 1973 (in Russian).
Manheim, F. T., 1976, Interstitial waters of marine sediments, in J. P. Riley et al, eds., Chemical oceanography, V. 6, 2d ed.: New York, Academic Press, p. 115-186.
Momper, J. A., and J. A. Williams, 1979, Geochemical exploration in Powder River basin, northeastern Wyoming and southeastern Montana (abs.): AAPG Bull., V. 63, p. 497.
Morris, K. A., 1979, A classification of Jurassic marine shale sequences: an example from the Toarcian (Lower Jurassic) of Great Britain: Paleogeography, Paleoclimatology, Paleoecology, v. 26, p. 117-120.
Nissenbaum, A., M. J. Baedecker, and I. R. Kaplan, 1972, Dissolved organic matter from interstitial organic water of a reducing marine fjord, in Advances in organic geochemistry, 1971: Intemat. Ser. Mon. Earth Sci., v. 33, p. 427-440.
Nixon, R. P., 1973, Oil source beds in Cretaceous Mow-ry Shale of Northwestern Interior United States: AAPG BuU., V. 57, p. 136-161.
Oremland, R. S., and B. F. Taylor, 1978, Sulfate reduction and methanogenesis in marine sediments: Geo-chim. et Cosmochim. Acta, v. 42, p. 209-214.
Orr, W. L., and A. G. Gaines, 1974, Observations on rate of sulfate reduction and organic matter oxidation in the bottom waters of an estuarine basin: the upper basin of the Pettaquamscutt River (Rhode Island), in Advances in organic geochemistry, 1974: Paris, Tech-nip, p. 790-812.
Palfrey, M. K., and G. G. Day, 1968, Oceanography of Baffin Bay and Nares Strait in the summer of 1968: Washington, D.C., Coast Guard Rept. 373-16.
Parker, R. H., 1964, Zoogeography and ecology of macro-invertebrates of GtUf of California and continental slope of westem Mexico, in Marine geology of the Gulf of California: AAPG Mem. 3, p. 331-376.
Parrish, J. T., K. S. Hansen, and A. M. Ziegler, 1979, Atmospheric circulation and upwelling in Paleozoic, with reference to petroleum source beds (abs.): AAPG Bull., V. 63, p. 507-508.
Pelet, R., and Y. Debyser, 1977, Organic geochemistry of Black Sea cores: Geochim. et Cosmochim. Acta, v. 41, p. 1575-1586.
Peng, T. H., et al, 1977, Benthic mixing in deep-sea cores as determined by C'* dating and its implication regarding climate, stratigraphy and the fate of fossil fuel CO2, in N. R. Andersen, ed.. The fate of fossil fuel CO2 in the oceans: New York, Plenum Press, p. 355-373.
Phleger, F. B, 1964, Patterns of living benthonic Foram-inifera, Gulf of California, in Marine geology of the Gulf of California: AAPG Mem. 3, p. 377-394.
Redfield, A. C , 1958, Preludes to the entrapment of organic matter in the sediments of Lake Maracaibo, in L. G. Weeks, ed., Habitat of Oil: AAPG, p. 968-981.
Reid, J. L., 1965, Intermediate waters of the Pacific Ocean: Baltimore, Johns Hopkins Univ. Press, 85 p.
Rhoads, D. C , 1974, Organism-sediment relationship on the muddy sea floor: Oceanogr. Marine Biology Ann. Rev., v. 12, p. 263-300.
Richards, F. A., 1970, The enhanced preservation of organic matter in anoxic marine environments, in Symposium on organic matter in natural waters: Univ. Alaska Inst. Marine Sci. Occasional Pub. 1, p. 399-411.
1976, The Cariaco basin (trench): Oceanogr. Marine Biology Ann. Rev., v. 13, p. 11-67.
and R. F. Vaccaro, 1956, The Cariaco Trench, an anaerobic basin in the Caribbean Sea: Deep-Sea Res., V. 7, p. 163-172.
Roden, G. I., 1964, Oceanographic aspects of Gulf of California, in Marine geology of the Gulf of California: AAPG Mem. 3, p. 30-58.
Roehler, H. W., 1974, Depositional environment of rocks in the Piceance Creek basin, Colorado: Rocky Mtn. Assoc. Geologists Field Conf. Guidebook 25, p. 57-64.
Romankevich, E. A., 1977, Organic geochemistry of oceanic sediments: Moscow, Izd. Nauka, p. 13 (in Russian).
Rosenberg, R., 1977, Benthic macrofaunal dynamics, production and dispersion on an oxygen-deficient estuary of west Sweden: Jour. Exp. Marine Biology Ecology, v. 26, p. 107-133.
Ross, D. A., and E. T. Degens, 1974, Recent sediments of Black Sea, in The Black Sea: AAPG Mem. 20, p.

Anoxic Environments 1209
78. Theede, H., et al, 1969, Studies on the resistance of ma
rine bottom invertebrates to oxygen deficiency and hydrogen sulfide: Mar. Biology, v. 2, p. 325-337.
Thiede, J., and T. H. van Andel, 1977, The paleoenvi-ronment of anaerobic sediments in the late Mesozoic South Atlantic Ocean: Earth and Planetary Sci. Letters, V. 33, p. 301-309.
Tissot, B. P., 1979, Effects on prolific petroleum source rocks and major coal deposits caused by sea-level changes: Nature, v. 277, p. 377-380.
and D. H. Welte, 1978, Petroleum formation and occurrence: New York, Springer-Verlag, 521 p.
G. Deroo, and A. Hood, 1978, Geochemical study of the Uinta basin; formation of petroleum from the Green River Formation: Geochim. et Cos-mochim. Acta, v. 42, p. 1469^1486.
- et al, 1974, Influence of nature and diagenesis of organic matter in formation of petroleum: AAPG BuU., V. 58, p. 499-506.
et al, 1979, Paleoenvironment and petroleum potential of mid-Cretaceous black shales in Atlantic basins (abs.): AAPG Bull., v. 63, p. 542.
Turekian, K. K., J. K. Cochran, and D. J. De Master, 1978, Bioturbation in deep-sea deposits: rates and consequences: Oceanus, v, 21, p. 34-41.
Tyson, R. V., R. C. L. Wilson, and C. Downie, 1979, A stratified water column enviromnental model for the type Kimmeridge clay: Nature, v. 277, p. 377-380.
Udintsev, Gb., ed., 1975, Geological-geophysical atlas of the Indian Ocean: New York, Pergamon Press, 151 P-
Usher, J. L., and P. Supko, 1978, Initial reports of the Deep Sea Drilling Project, v. 42, pt. 2, 1243 p.
Vail, P. R., and R. M. Mitchum, 1979, Global cycles of sea level change and their role in exploration: 10th World Petroleum Cong., Preprint, Bucharest Panel Discussion 2, Paper 4.
van Andel, T. H., 1964, Recent marine sediments of Gulf of California, in Marine geology of Gulf of California: AAPG Mem. 3, p. 216-310.
Veber, V. V., 1977, Generation of hydrocarbon gases in modem sedimentary formation of the eastern and southern parts of the Atlantic Ocean: Geologyia Nef-ti i Gaza, no. 1, p. 59-64.
Veeh, H. H., 1979, Modem environment of phosphorite formation and the geochemical balance of phosphorus in the ocean, in P. J. Cook and J. H. Shergold, eds., Proterozoic-Cambrian phosphorites: Project 156 of UNESCO-IUGS: Canberra, Australian Natl. Univ. Press, p. 59.
W. C. Bumett, and A. Soutar, 1973, Contemporary phosphorites on the continental margin of Pern: Science, v. 181, p. 844-845.
• S. E. Calvert, and N. B. Price, 1974, Accumulation of uranium in sediments and phosphorites on the southwest African shelf: Marine Chemistry, v. 2, p. 189-202.
Wolfe, R. S., 1971, Microbial formation of methane: Adv. Microbial Physiology, v. 6, p. 107-146.
Wyrtki, K., 1962, The oxygen minimum in relation to ocean circulations: Deep-Sea Research, v. 9, p. 11-23.
Ziegler, P. A., 1979, Factors controlling North Sea hydrocarbon accumulations: World Oil, v. 189, no. 6, p. 111-124.
J. T. Parrish, and R. C. Humphreville, 1979, Pa-leogeography, upwelling and phosphorites (abs.), in P. J. Cook and J. H. Shergold, eds., Proterozoic-Cambrian phosphorites. Project 156 of UNESCO-IUGS: Canberra, Australian Natl. Univ. Press, p. 21.
183-199. Ryan, W. B. F., and M. B. Cita, 1977, Ignorance con
cerning episodes of ocean-wide stagnation: Marine Geology, v. 23, p. 197-215.
Sackett, W. M., et al, 1978, A carbon inventory for Orca brines and sediments: Earth and Planetary Sci. Letters, V. 44, p. 73-81.
SaUot, A., 1977, Etude comparative des acides gras de I'eau et de I'eau interstitieUe a Tinterface eau de mer profonde/sediment: Acad. Sci. Comptes Rendus, ser. D-857, p. 285 (10 Oct. 1977).
Schaefer, W., 1972, Ecology and palaeoecology of marine environments: Chicago, Univ. Chicago Press, 568 p.
Schlanger, S. O., and H. C. Jenkyns, 1976, Cretaceous oceanic anoxic events: causes and consequences: Geologic en Mijnbouw, v. 55, p. 179-184.
Scranton, M. I., and J. W. Farrington, 1977, Methane production in the waters off Widvis Bay: Jour. Geo-phys. Research, v. 82, p. 4947-4953.
Shimkus, K. M., and E. S. Trimonis, 1974, Modem sedimentation in Black Sea, in The Black Sea: AAPG Mem. 20, p. 249-278.
Shokes, R. F., et al, 1977, Anoxic hypersaline basin in the northem Gulf of Mexico: Science, v. 196, p. 1443-1446.
Smith, J. W., and W. A. Robb, 1973, Aragonite and the genesis of carbonates in Mahogany zone oil shales of Colorado's Green River Formation: U.S. Bur. Mines Rept. 7727, 21 p.
Spencer, D. W., H. Susumu, and D. G. Brewer, 1978, Particles and particle fluxes in the ocean: Oceanus, v. 21, p. 20-25.
Stackelberg, U. V, 1972, Faziesverteilung in Sedimenten des indisch-pakistanischen Kontinentalrandes (Ara-bisches Meer): "Meteor" Forschungsergebnisse, Rei-he C, No. 9, p. 1-173.
Stanley, D. J., 1978, Ionian Sea sapropel distribution and late Quaternary paleo-oceanography in the east-em Mediterranean: Nature, v. 274, p. 149-151.
Stevenson, F. J., and C. N. Cheng, 1972, Organic geochemistry of the Argentine basin sediments: carbon-nitrogen relationships and Quaternary correlations: Geochim. et Cosmochim. Acta, v. 36, p. 653-671.
Stommel, H., 1958, The circulation of the abyss, reprinted, 1977, in Ocean science, readings from Scientific American: San Francisco, W. H. Freeman and Co., p. 139-144.
Summerhayes, C. P., U. de Melo, and H. T. Baretto, 1976, The influence of upwelling on suspended matter and shelf sediments off southeastern Brazil: Jour. Sed. Petrology, v. 46, p. 819-828.
Swain, F. M., 1970, Non-marine organic geochemistry: Cambridge, U.K., Cambridge Univ. Press, 445 p.
Swanson, V. E., 1960, Oil yield and uranium content of black shales: U.S. Geol. Survey Prof. Paper 356-A, p. 1-44.
TaUing, J. F., 1963, Origin of stratification in an African rift lake: Limnology and Oceanography, v. 8, p. 68-