arthropod-borne virus infections of aborigines
TRANSCRIPT

Bull. Org. mond. Sante 1 1964 30, 211-219Bull. Wld Hlth Org. J
Arthropod-borne Virus Infections of AboriginesIndigenous to Australasia
A Preliminary Report *
C. L. WISSEMAN Jr,' D. C. GAJDUSEK,2 F. D. SCHOFIELD3 & E. C. ROSENZWEIG'
Epidemics of presumed arthropod-borne virus infections have occurred over the yearsin the Australasian region but as yet little is known about the identity, distribution andfrequency of occurrence of the viruses present. Therefore, beginning with specimens col-lected in 1956, a systematic sero-epidemiological study was undertaken to provide pre-liminary information on arthropod-borne virus infections of aborigines inhabiting partsofAustralia, New Guinea, New Britain and islands ofMicronesia and Melanesia. Distinct-ive antibody patterns are emerging which have important implications. For example, in-habitants of most coastal lowland regions studied gave evidence of a high incidence ofinfection with a multiplicity ofgroup A and group B viruses. In New Guinea, dissociationbetween group A and group B viruses apparently occurs as altitude increases, with group Bantibodies disappearing above about 1500 metres, while group A antibodies are still.foundat considerably higher altitudes. Circumscribed areas show unusual antibody patterns.
Epidemics of arthropod-borne or presumedarthropod-borne virus infections (dengue anddengue-like, polyarthritis with rash, central nervoussystem infections) have been recognized from timeto time over the years in various parts of the Austral-asian region. For the most part, however, littleis known of the identity of the viral agents present,their distribution or the frequency of their occurrenceas causes of infection in man.Both epidemic and sporadic cases of acute viral
infections of the central nervous system have beenobserved in many areas of New Guinea and the
* Revised version of a paper submitted to the Seminaron Japanese Encephalitis and Other Arthropod-borneVirus Infections convened in Tokyo in November 1962 bythe WHO Regional Office for the Western Pacific. A portionof this paper was previously presented to the Symposiumon Medical Ecology held at the Tenth Pacific Science Con-gress in Honolulu, Hawaii, and is to be published in thePapua and New Guinea Medical Journal. The work receivedpartial support from Contract No. DA-49-007-MD-660with the Medical Research and Development Command,Office of the Surgeon General, Department of the Army(USA) under the aegis of the Commission on Immunizationof the Armed Forces Medical Board.
1 Department of Microbiology, University of MarylandSchool of Medicine, Baltimore, Md., USA.
2 National Institute of Neurological Diseases and Blind-ness, National Institutes of Health, Bethesda, Md., USA.
' Director of Medical Research, Public Health Depart-ment, Port Moresby, Territory of Papua and New Guinea.
surrounding islands of Melanesia and MicronesiaIn some regions of New Guinea where we havehad an opportunity for intensive observation,acute encephalitis and aseptic meningo-encephalitis,particularly in infants and small children, havebeen found to be common problems. This isparticularly true in the Sepik District and in theKup region of western Chimbu in the highlands.In both of these regions, as well as in several otherareas of study in New Guinea, many cases of spasticor flaccid pareses, mental defect, sensory defect,and cranial nerve syndromes have been identifiedin direct association with acute episodes of febrileencephalitic disease. Many of the so-called cerebralmalarial deaths are undoubtedly due to acute viralencephalitis, and this should become increasinglyevident as malaria control in New Guinea is ex-tended.
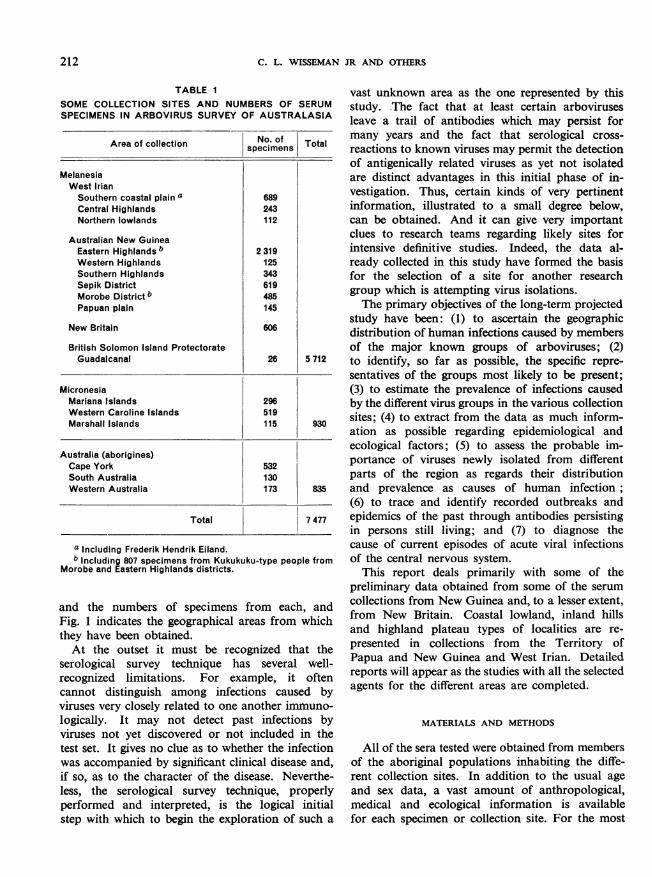
Beginning with serum collections obtained in1956, extensively supplemented by more recent andcontinuing collections, a systematic study wasbegun of arbovirus infections of aborigines in-habiting parts of Australia, New Guinea, NewBritain and many of the islands of Melanesia andMicronesia by testing serum specimens for thepresence of antibodies. Table 1 presents a partiallist of the sites sampled and catalogued to date
1393 -211 -
C. L. WISSEMAN JR AND OTHERS
TABLE ISOME COLLECTION SITES AND NUMBERS OF SERUMSPECIMENS IN ARBOVIRUS SURVEY OF AUSTRALASIA
Area of collection No. of Totalspecimens ___________________
MelanesiaWest IrianSouthern coastal plain a 689Central Highlands 243Northern lowlands 112
Australian New GuineaEastern Highlands b 2 319Western Highlands 125Southern Highlands 343Sepik District 619Morobe District b 485Papuan plain 145
New Britain 606
British Solomon Island ProtectorateGuadalcanal 26 5 712
MicronesiaMariana Islands 296Western Caroline Islands 519Marshall Islands 115 930
Australia (aborigines)Cape York 532South Australia 130Western Australia 173 835
Total 7 477
a Including Frederik Hendrik Eiland.b Including 807 specimens from Kukukuku-type people from
Morobe and Eastern Highlands districts.
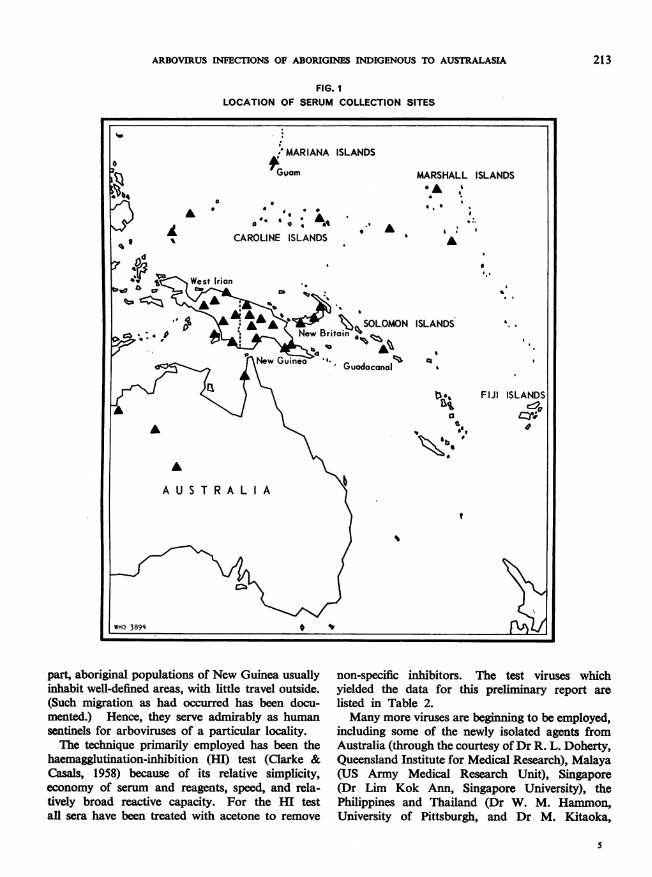
and the numbers of specimens from each, andFig. 1 indicates the geographical areas from whichthey have been obtained.At the outset it must be recognized that the
serological survey technique has several well-recognized limitations. For example, it oftencannot distinguish among infections caused byviruses very closely related to one another immuno-logically. It may not detect past infections byviruses not yet discovered or not included in thetest set. It gives no clue as to whether the infectionwas accompanied by significant clinical disease and,if so, as to the character of the disease. Neverthe-less, the serological survey technique, properlyperformed and interpreted, is the logical initialstep with which to begin the exploration of such a
vast unknown area as the one represented by thisstudy. The fact that at least certain arbovirusesleave a trail of antibodies which may persist formany years and the fact that serological cross-reactions to known viruses may permit the detectionof antigenically related viruses as yet not isolatedare distinct advantages in this initial phase of in-vestigation. Thus, certain kinds of very pertinentinformation, illustrated to a small degree below,can be obtained. And it can give very importantclues to research teams regarding likely sites forintensive definitive studies. Indeed, the data al-ready collected in this study have formed the basisfor the selection of a site for another researchgroup which is attempting virus isolations.The primary objectives of the long-term projected
study have been: (1) to ascertain the geographicdistribution of human infections caused by membersof the major known groups of arboviruses; (2)to identify, so far as possible, the specific repre-sentatives of the groups most likely to be present;(3) to estimate the prevalence of infections causedby the different virus groups in the various collectionsites; (4) to extract from the data as much inform-ation as possible regarding epidemiological andecological factors; (5) to assess the probable im-portance of viruses newly isolated from differentparts of the region as regards their distributionand prevalence as causes of human infection;(6) to trace and identify recorded outbreaks andepidemics of the past through antibodies persistingin persons still living; and (7) to diagnose thecause of current episodes of acute viral infectionsof the central nervous system.
This report deals primarily with some of thepreliminary data obtained from some of the serumcollections from New Guinea and, to a lesser extent,from New Britain. Coastal lowland, inland hillsand highland plateau types of localities are re-presented in collections from the Territory ofPapua and New Guinea and West Irian. Detailedreports will appear as the studies with all the selectedagents for the different areas are completed.
MATERIALS AND METHODS
All of the sera tested were obtained from membersof the aboriginal populations inhabiting the diffe-rent collection sites. In addition to the usual ageand sex data, a vast amount of anthropological,medical and ecological information is availablefor each specimen or collection site. For the most
212
ARBOVIRUS INFECTIONS OF ABORIGNS INDIGENOUS TO AUSTRALASIA
FIG. ILOCATION OF SERUM COLLECTION SITES
part, aboriginal populations of New Guinea usuallyinhabit well-defined areas, with little travel outside.(Such migration as had occurred has been docu-mented.) Hence, they serve admirably as humansentinels for arboviruses of a particular locality.The technique primarily employed has been the
haemagglutination-inhibition (HI) test (Clarke &Casals, 1958) because of its relative simplicity,economy of serum and reagents, speed, and rela-tively broad reactive capacity. For the HI testall sera have been treated with acetone to remove
non-specific inhibitors. The test viruses whichyielded the data for this preliminary report arelisted in Table 2.Many more viruses are beginning to be employed,
including some of the newly isolated agents fromAustralia (through the courtesy ofDr R. L. Doherty,Queensland Institute for Medical Research), Malaya(US Army Medical Research Unit), Singapore(Dr Lim Kok Ann, Singapore University), thePhilippines and Thailand (Dr W. M. Hammon,University of Pittsburgh, and Dr M. Kitaoka,
213

C. L. WISSEMAN JR AND OTHERS
TABLE 2VIRAL ANTIGENS EMPLOYED IN
SEROLOGICAL SURVEY
Group A Group B Group C
Chikungunya (Chik.) Dengue type I (DI) Apeu
Semliki Forest (Seml.) Dengue type 2 (D2) Oriboca
Dengue type 3 (D3) Marituba
Dengue type 4 (D4)
Murray Valleyencephalitis (MVE)
Japaneseencephalitis (JE)
Yellow fever (YF)
National Institute of Health, Tokyo). The resultsof tests with these agents either have not beenanalysed or are not complete at this time. When theresults of the HI test indicate it, neutralization testsare performed on selected sera for more precise
identification of the agents involved.
RESULTS
The preliminary results available indicate thatarbovirus infections have been prevalent amonghuman inhabitants of many regions of New Guineaand New Britain, that a multiplicity of viral agentsis involved and that certain factors apparentlylimit the spread of one group of viruses to a greaterextent than another group. Selected data are
presented below to illustrate these points.
Relationship of ecological setting to prevalence ofarbovirus antibodies
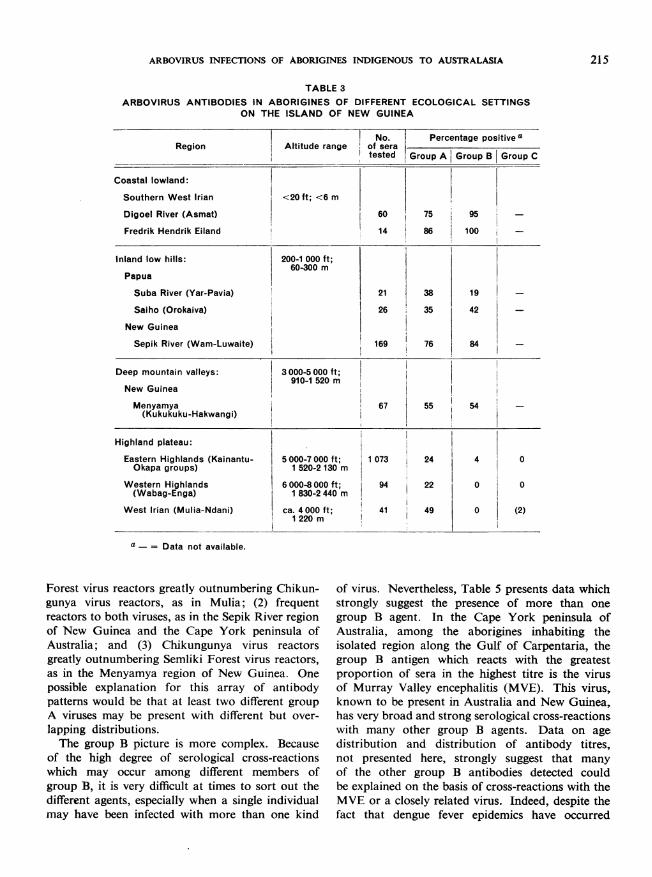
In New Guinea, antibodies to both group A andgroup B viruses were very prevalent among theinhabitants of low-altitude regions tested, whichincluded coastal and inland areas. However,antibody patterns show a striking dissociationbetween the group A and the group B viruses athigher altitudes. Thus, in the Eastern Highlands,a region of inhabited valleys ranging between about5000 and 7000 feet (about 1520-2130 m) in elevation,the proportion of specimens reacting with any of thegroup B viral antigens markedly decreased relativeto the proportion reacting to group A viral antigens(Table 3). In the Western Highlands, no group Breactors were detected, despite the fact that approxi-
mately 22% of the specimens were positive withgroup A antigens. This suggests that some factorassociated with altitude may affect the transmissionof the group B viruses of the area, possibly eitherby limiting the distribution of vectors or reservoirhosts or by affecting virus multiplication within thevector (for example, a temperature effect). The samefactors do not seem to limit the transmission of thegroup A viruses to the same extent.At first glance, the difference in prevalence of
group B viral antibodies between the Menyamyaregion (about 3000-5000 feet; or about 910-1520 m)and the Mulia region (about 4000 feet; or about1220 m) might detract from the altitude hypothesis.However, the two regions are not strictly comparable.The Menyamya region studied is an area of highmountain ridges (where the villages are situated)and deep valleys (where the gardens are located).Thus, the people of this region descend to the lower,malarious zones regularly to tend their gardens.On the other hand, Mulia is a high, narrow valleywhere descent to lower altitudes is not common.
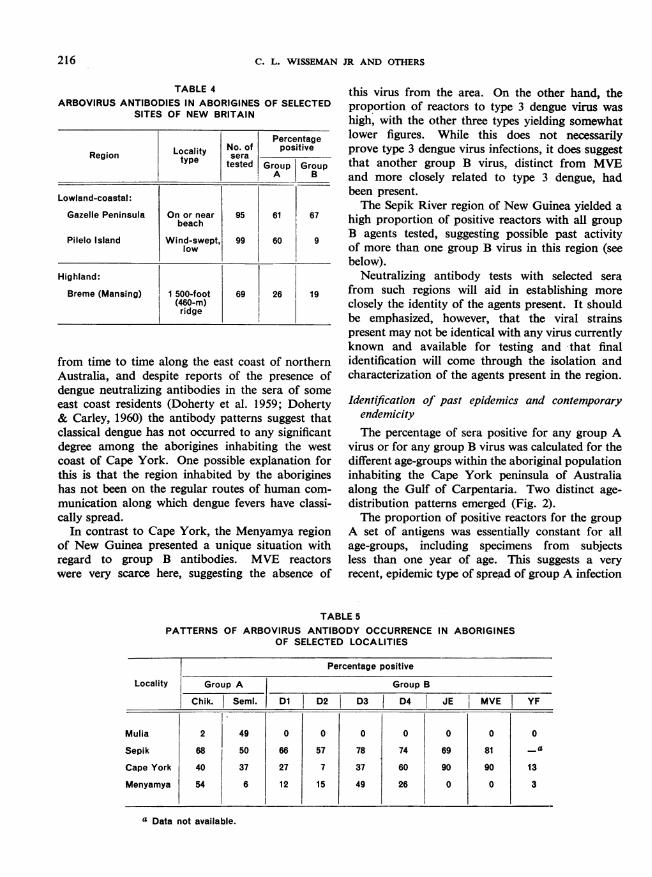
In New Britain, only sera from inhabitants oflow coastal areas and relatively low mountain ridgeshave been tested to date (Table 4). Both group Aand group B antibodies are present. An interestingdissociative trend-not absolute, however-wasnoted in the collection from Pilelo Island, a low,wind-swept island separated from the southerncoast of New Britain by a narrow strip of water.Here group A antibodies were prevalent whereasgroup B antibodies were relatively rare. Theexplanation for this has not yet been uncovered.However, it is anticipated that a variety of interestingpatterns reflecting different ecological and epidemio-logical features will evolve as samples are tested fromthe many smaller, more isolated islands where thenumbers of vertebrates and insects become smallerboth absolutely and in array of species.
Multiplicity of arboviruses
While little or no evidence for the presence ofgroup C virus antibodies has yet been encounteredin any area studied, it is apparent that representativesof both group A and group B are widely distributed.Even though the number of agents included inthe test sets of these two groups is still small, thedata from selected areas strongly suggest that morethan one representative of each of these two groupsmay be present in different areas (Table 5). Thus,with only two group A viruses, at least three differentantibody patterns have been found: (1) Semliki
214
ARBOVIRUS INFECTIONS OF ABORIGINES INDIGENOUS TO AUSTRALASIA
TABLE 3
ARBOVIRUS ANTIBODIES IN ABORIGINES OF DIFFERENT ECOLOGICAL SETTINGSON THE ISLAND OF NEW GUINEA
No. Percentage positive aRegion Altitude range of sera
tested Group A Group B Group C
Coastal lowland:
Southern West Irian <20 ft; <6 m
Digoel River (Asmat) 60 75 95
Fredrik Hendrik Eiland 14 86 100
Inland low hills: 200-1 000 ft;60-300 m
Papua
Suba River (Yar-Pavia) 21 38 19
Saiho (Orokaiva) 26 35 42
New Guinea
Sepik River (Wam-Luwaite) 169 76 84
Deep mountain valleys: 3000-5 000 ft;910-1 520 m
New Guinea
Menyamya 67 55 54 -(Kukukuku-Hakwangi)
Highland plateau:
Eastern Highlands (Kainantu- 5000-7000 ft; 1 073 24 4 0Okapa groups) 1 520-2 130 m
Western Highlands 6000-8000 ft; 94 22 0 0(Wabag-Enga) 1 830-2 440 m
West Irian (Mulia-Ndani) ca. 4000 ft; 41 49 0 (2)1220 m
a - = Data not available.
Forest virus reactors greatly outnumbering Chikun-gunya virus reactors, as in Mulia; (2) frequentreactors to both viruses, as in the Sepik River regionof New Guinea and the Cape York peninsula ofAustralia; and (3) Chikungunya virus reactorsgreatly outnumbering Semliki Forest virus reactors,as in the Menyamya region of New Guinea. Onepossible explanation for this array of antibodypatterns would be that at least two different groupA viruses may be present with different but over-lapping distributions.The group B picture is more complex. Because
of the high degree of serological cross-reactionswhich may occur among different members ofgroup B, it is very difficult at times to sort out thedifferent agents, especially when a single individualmay have been infected with more than one kind
of virus. Nevertheless, Table 5 presents data whichstrongly suggest the presence of more than onegroup B agent. In the Cape York peninsula ofAustralia, among the aborigines inhabiting theisolated region along the Gulf of Carpentaria, thegroup B antigen which reacts with the greatestproportion of sera in the highest titre is the virusof Murray Valley encephalitis (MVE). This virus,known to be present in Australia and New Guinea,has very broad and strong serological cross-reactionswith many other group B agents. Data on agedistribution and distribution of antibody titres,not presented here, strongly suggest that manyof the other group B antibodies detected couldbe explained on the basis of cross-reactions with theMVE or a closely related virus. Indeed, despite thefact that dengue fever epidemics have occurred
215
C. L. WISSEMAN JR AND OTHERS
TABLE 4ARBOVIRUS ANTIBODIES IN ABORIGINES OF SELECTED
SITES OF NEW BRITAIN
PercentageLocal ity No. of positiveRegion tye seratype tested Group Group_'A B
Lowland-coastal:
Gazelle Peninsula On or near 95 61 67beach
Pilelo Island Wind-swePt. 99 60 9low
Highland:
Breme (Mansing) 1 500-foot 69 26 19(460-m)ridge
from time to time along the east coast of northernAustralia, and despite reports of the presence ofdengue neutralizing antibodies in the sera of someeast coast residents (Doherty et al. 1959; Doherty& Carley, 1960) the antibody patterms suggest thatclassical dengue has not occurred to any significantdegree among the aborigines inhabiting the westcoast of Cape York. One possible explanation forthis is that the region inhabited by the aborigineshas not been on the regular routes of human com-
munication along which dengue fevers have classi-cally spread.
In contrast to Cape York, the Menyamya regionof New Guinea presented a unique situation withregard to group B antibodies. MVE reactorswere very scarce here, suggesting the absence of
this virus from the area. On the other hand, theproportion of reactors to type 3 dengue virus was
high, with the other three types yielding somewhatlower figures. While this does not necessarilyprove type 3 dengue virus infections, it does suggestthat another group B virus, distinct from MVEand more closely related to type 3 dengue, hadbeen present.The Sepik River region of New Guinea yielded a
high proportion of positive reactors with all groupB agents tested, suggesting possible past activityof more than one group B virus in this region (seebelow).
Neutralizing antibody tests with selected serafrom such regions will aid in establishing more
closely the identity of the agents present. It shouldbe emphasized, however, that the viral strainspresent may not be identical with any virus currentlyknown and available for testing and that finalidentification will come through the isolation andcharacterization of the agents present in the region.
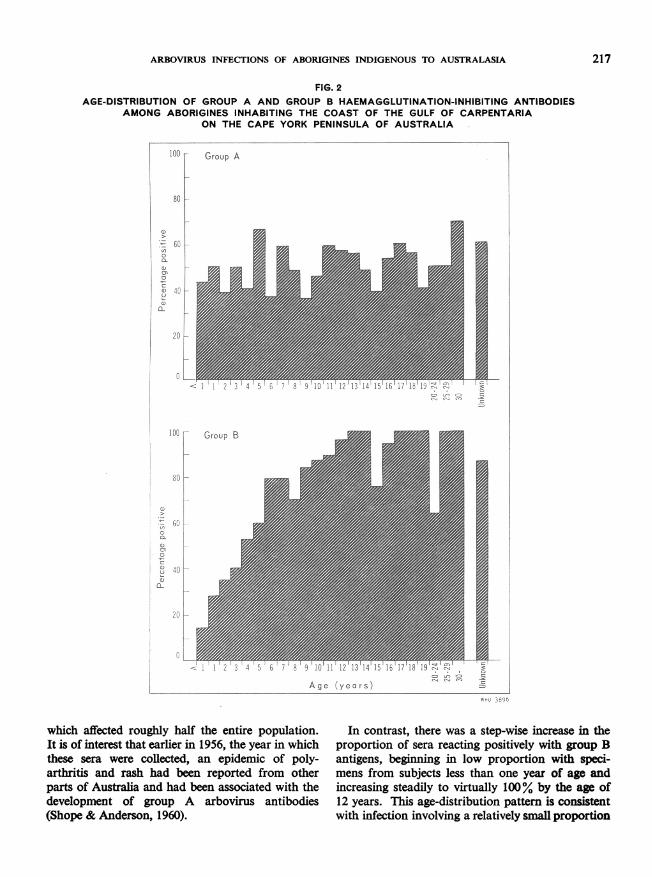
Identification of past epidemics and contemporaryendemicityThe percentage of sera positive for any group A
virus or for any group B virus was calculated for thedifferent age-groups within the aboriginal populationinhabiting the Cape York peninsula of Australiaalong the Gulf of Carpentaria. Two distinct age-distribution patterns emerged (Fig. 2).The proportion of positive reactors for the group
A set of antigens was essentially constant for allage-groups, including specimens from subjectsless than one year of age. This suggests a veryrecent, epidemic type of spread of group A infection
TABLE 5PATTERNS OF ARBOVIRUS ANTIBODY OCCURRENCE
OF SELECTED LOCALITIESIN ABORIGINES
Percentage positive
Locality Group A Group B
Chik. Seml. D1 D2 D3 D4 JE MVE| YF
Mulia 2 49 0 0 0 0 0 0 0
Sepik 68 50 66 57 78 74 69 81 -_aCape York 40 37 27 7 37 60 90 90 13
Menyamya 54 6 12 15 49 26 0 0 3
a Data not available.
216
ARBOVIRUS INFECTIONS OF ABORIGINES INDIGENOUS TO AUSTRALASIA
FIG. 2AGE-DISTRIBUTION OF GROUP A AND GROUP B HAEMAGGLUTINATION-INHIBITING ANTIBODIES
AMONG ABORIGINES INHABITING THE COAST OF THE GULF OF CARPENTARIAON THE CAPE YORK PENINSULA OF AUSTRALIA
100 Group A
80 -
8)
600
0a
a
* 40Pa
0.
0
°00r
80 F
t 600CL
cn
u( 40a-
201-
Age (years)whO 3896
Iwhich affected roughly half the entire population.It is of interest that earlier in 1956, the year in whichthese sera were collected, an epidemic of poly-arthritis and rash had been reported from otherparts of Australia and had been associated with thedevelopment of group A arbovirus antibodies(Shope & Anderson, 1960).
In contrast, there was a step-wise increase in theproportion of sera reacting positively with group Bantigens, beginning in low proportion with speci-mens from subjects less than one year of age andincreasing steadily to virtually 100% by the age of12 years. This age-distribution pattern is consistentwith infection involving a relatively small proportion
.De
=5
217
C= ull O
061-b e;.i 3:C:. ';. C. 0cli cm m ..
LTetp I cn, I
C. L. WISSEMAN JR AND OTHERS
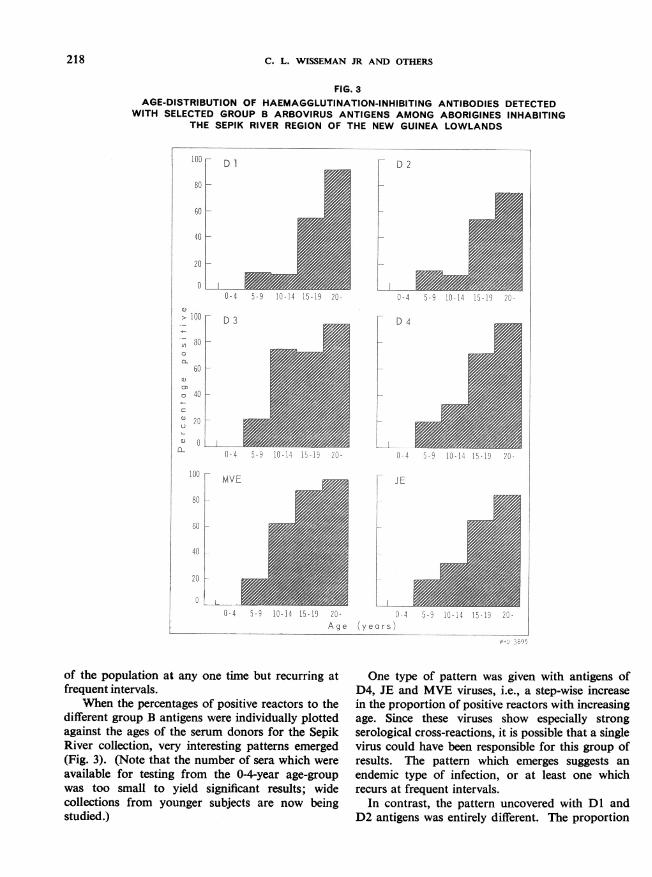
FIG. 3AGE-DISTRIBUTION OF HAEMAGGLUTINATION-INHIBITING ANTIBODIES DETECTED
WITH SELECTED GROUP B ARBOVIRUS ANTIGENS AMONG ABORIGINES INHABITINGTHE SEPIK RIVER REGION OF THE NEW GUINEA LOWLANDS
5- 9 10- 14 15 -19 20-
D 2
0-4
D 4
0 -4
JE
- I I...L20- 0-4Age (years)
5b- 10-14 15-19 ZU-
of the population at any one time but recurring atfrequent intervals.
'When the percentages of positive reactors to thedifferent group B antigens were individually plottedagainst the ages of the serum donors for the SepikRiver collection, very interesting patterns emerged(Fig. 3). (Note that the number of sera which wereavailable for testing from the 0-4-year age-groupwas too small to yield significant results; widecollections from younger subjects are now beingstudied.)
One type of pattern was given with antigens ofD4, JE and MVE viruses, i.e., a step-wise increasein the proportion of positive reactors with increasingage. Since these viruses show especially strongserological cross-reactions, it is possible that a singlevirus could have been responsible for this group ofresults. The pattern which emerges suggests anendemic type of infection, or at least one whichrecurs at frequent intervals.
In contrast, the pattern uncovered with Dl andD2 antigens was entirely different. The proportion
'wF Dl
80
60 -
40 _
0-4
D3
3,, iu-i4q - 1 LZU-
20
0
0> 100
, 80
ac0
o 40C
u' 20
0 0cL
F
_-
0-4
°° F MVE80 k
601-
40 -
20
00-4
WHO 3895
218
3>I s U-14 1>w1M

ARBOVIRUS INFECTIONS OF ABORIGINES INDIGENOUS TO AUSTRALASIA 219
of reactors was uniformly very low in the youngerage-groups. Then, in the 15-19-year age-group andabove, the proportion of positive reactors showedan abrupt and marked increase. This kind ofpattern is consistent with an epidemic spread ofinfection which occurred between 15 and 19 yearsprior to the collection of the sera and which involveda very high proportion of the persons alive at thattime. The low proportion of reactors in the youngerage-groups could be explained on the basis of cross-reactions with related viruses known to be present(see above). It is of interest to note that this suddenincrease in reactors with the Dl and D2 antigenscoincides with the age-groups who were alive at thetime of the extensive dengue epidemics on NewGuinea during the Second World War from whichboth type 1 and type 2 viruses were isolated bySabin (1952).
CONCLUSIONS
Despite its shortcomings, the serological surveytechnique for the study of arthropod-borne virusinfections of man shows promise of yielding a largeamount of valuable information regarding thepresence or absence of major groups of arbovirusinfections in man, their geographic distribution,their prevalence and some clues to epidemiologicalfeatures and ecological factors in the Australasianregion. Clinical study has revealed that acutefebrile encephalitic disease, particularly in youngerchildren, is a major problem in many areas of theNew Guinea lowlands and highlands. It is expectedthat, as the results of these studies become available,there will be a more rational basis for the selectionof uniquely appropriate sites for isolation of arthro-pod-borne viruses, the identification of the clinicaldiseases and the study of the ecology of specific viruses.
RESUME
Des epidemies de maladies dues ou supposees dues ades virus transmis par les arthropodes (dengue, infectionsdu systeme nerveux central, par exemple) ont sevi aucours des annees en Australasie. Mais on savait peu dechose sur l'identite, la repartition et le taux de presencedes virus en cause. Pour combler ces lacunes, les auteursont entrepris une etude sero-6pidemiologique systema-tique, sur les aborigenes vivant dans certaines parties del'Australie, de la Nouvelle-Guin6e, de la Nouvelle-Bretagne et des iles de la Micronesie et Melanesie.ils en apportent les premiers resultats, qui ont faitapparaitre d'interessants schemas de repartition desanticorps.Les anticorps des groupes A et B des arbovirus sont
largement representes, tandis que ceux du groupe C sontquasi inexistants. II semble que chacun de ces groupes soit
represente par plus d'un virus - le groupe A par le virusde Semliki et de Chikungunya, diversement repartis, l'unou l'autre dominant suivant les regions. Les r6ponsesserologiques aux virus du groupe B sont plus difficiles itinterpreter, en raison d'une forte tendance aux reactionscroisees entre virus de ce groupe. 1i se peut que des virusencore inconnus soient presents, qu'il s'agira de recher-cher et d'isoler au cours d'etudes ulterieures. En Nouvelle-Guinee, une dissociation entre les groupes A et B semanifeste, quand l'altitude augmente: les anticorps dugroupe B disparaissent au-dessus de 1500 m, tandis queles anticorps A se rencontrent A des altitudes tres supe-rieures. Les etudes cliniques, d'autre part, ont montreque l'encephalite febrile aigue pose un grave problemechez les jeunes enfants surtout, dans plusieurs zones desterres basses et hautes de Nouvelle-Guinee.
REFERENCES
Clarke, D. H. & Casals, J. (1958) Amer. J. trop Med. Hyg., 7, 561Doherty, R. L. & Carley, J. G. (1960) Aust. J. exp. Biol. med. Sci., 38, 427Doherty, R. L., Carley, J. G. & Lee, P. E. (1959) Aust. J. exp. Biol. med. Sci., 37, 365Sabin, A. B. (1952) Amer. J. trop. Med. Hyg., 1, 30Shope, R. E., & Anderson, S. G. (1960) Med. J. Aust., 1, 156