arabidopsis hsp90 protein modulates rpp4-mediated...
TRANSCRIPT

Arabidopsis HSP90 protein modulates RPP4-mediatedtemperature-dependent cell death and defense responses
Fei Bao1, Xiaozhen Huang1, Chipan Zhu2, Xiaoyan Zhang1, Xin Li2 and Shuhua Yang1
1State Key Laboratory of Plant Physiology and Biochemistry, College of Biological Sciences, National Plant Gene Research Center, China Agricultural University, Beijing 100193, China;
2Michael Smith Laboratories, University of British Columbia, Vancouver, Canada
Author for correspondence:Shuhua Yang
Tel: +86 10 6273 4838Email: [email protected]
Received: 12 January 2014
Accepted: 4 February 2014
New Phytologist (2014)doi: 10.1111/nph.12760
Key words: Arabidopsis, cell death, HSP90proteins, low temperature, RPP4.
Summary
� Plant defense responses are regulated by temperature. In Arabidopsis, the chilling-sensitive
mutant chs2-1 (rpp4-1d) contains a gain-of-function mutation in the TIR-NB-LRR (Toll and
interleukin 1 transmembrane receptor-nucleotide binding-leucine-rich repeat) gene, RPP4
(RECOGNITION OF PERONOSPORA PARASITICA 4), which leads to constitutive activation
of the defense response at low temperatures.� Here, we identified and characterized two suppressors of rpp4-1d from a genetic screen,
hsp90.2 and hsp90.3, which carry point mutations in the cytosolic heat shock proteins
HSP90.2 and HSP90.3, respectively.� The hsp90 mutants suppressed the chilling sensitivity of rpp4-1d, including seedling lethal-
ity, activation of the defense responses and cell death under chilling stress. The hsp90mutants
exhibited compromised RPM1 (RESISTANCE TO PSEUDOMONAS MACULICOLA 1)-, RPS4
(RIBOSOMAL PROTEIN S4)- and RPP4-mediated pathogen resistance. The wild-type RPP4
and the mutated form rpp4 could interact with HSP90 to form a protein complex. Further-
more, RPP4 and rpp4 proteins accumulated in the cytoplasm and nucleus at normal tempera-
tures, whereas the nuclear accumulation of the mutated rpp4 was decreased at low
temperatures. Genetic analysis of the intragenic suppressors of rpp4-1d revealed the impor-
tant functions of the NB-ARC and LRR domains of RPP4 in temperature-dependent defense
signaling. In addition, the rpp4-1d-induced chilling sensitivity was largely independent of the
WRKY70 orMOS (modifier of snc1) genes.� This study reveals that Arabidopsis HSP90 regulates RPP4-mediated temperature-depen-
dent cell death and defense responses.
Introduction
Temperature is one of the most important factors affectingplant growth and geographical distribution and the plantdefense response to pathogens (Guy, 1990; Alcazar & Parker,2011; Hua, 2013). High temperatures (32°C) inhibit anincrease in salicylic acid (SA) levels and prevent the inductionof the pathogenesis-related (PR) genes and resistance of tobaccoplants when infected with Tobacco mosaic virus (TMV). Theapplication of exogenous SA results in the recovery of PR geneexpression, but it cannot overcome the inhibition of the hyper-sensitive response (HR) (Malamy et al., 1992). Consistently,low temperatures can induce PR gene expression. Cold stresshas been shown to activate the transcription factor NAC,resulting in its translocation from the plasma membrane to thenucleus, which, in turn, induces PR genes by direct binding totheir promoters independent of SA (Seo et al., 2010). Anotherknown factor to mediate both the cold and biotic stressresponses is DEAR1 (DREB and EAR protein 1). DEAR1 isinduced by both pathogen infection and cold treatment. The
overexpression of DEAR1 in Arabidopsis results in dwarf andcell death phenotypes, as well as the constitutive expression ofPR genes and SA accumulation. However, the induction ofCBF (C-repeat binding factor) genes by cold treatment is sup-pressed in transgenic plants overexpressing the DEAR1 gene,leading to a decreased freezing tolerance (Tsutsui et al., 2009).It has been reported that the mekk1 mutant grown at 22°Csuffers from severe dwarfism and has a constitutively activedefense response, and this phenotype was greatly suppressedwhen the plants were grown at 28°C (Ichimura et al., 2006).In soybean plants, different temperature regimens during seeddevelopment can affect the expression of defense-related genes,resulting in varying resistance to pathogens (Upchurch &Ramirez, 2011).
The plant Resistance (R) genes encode immune receptors thatrecognize, directly or indirectly, pathogen effectors (Jones &Dangl, 2006). R proteins belonging to the largest class containnucleotide-binding (NB) and leucine-rich repeat (LRR) domains.Emerging evidence has shown that temperature can modulate theplant defense responses through the R proteins. The null mutant
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 1
www.newphytologist.com
Research

bon1-1 was found to display a miniature phenotype when grownat 22°C, but showed a wild-type appearance when grown at 28°C(Hua et al., 2001). Further study indicated that the NB-LRR geneSNC1 (Suppressor of npr1-1, constitutive 1) was activated in thebon1-1 mutant, leading to temperature-dependent constitutivedefense responses and reduced cell growth at 22°C, but not at28°C (Yang &Hua, 2004). It is intriguing to find that other pointmutations in SNC1 retain disease resistance at 28°C. Similarmutations introduced into the tobacco R gene N conferreddefense responses at an elevated temperature (Y. Zhu et al., 2010).Our previous studies have shown that mutations in the NB-LRRtype of R genes, including RPP4/CHS2 (RECOGNITION OFPERONOSPORA PARASITICA 4/CHILLING-SENSITIVE 2)and CHS3, result in the activation of the defense responses at lowtemperatures (Huang et al., 2010; Yang et al., 2010). Moreover,hybrid necrosis and genetic incompatibility were also found to bemodulated by the NB-LRR genes in a temperature-dependentmanner (Bomblies et al., 2007; Alcazar et al., 2009). In additionto these R proteins, our recent study has reported that a missensemutation of a TIR-NB (Toll and interleukin 1 transmembranereceptor-nucleotide binding) protein, CHS1, shows chilling-induced defense responses (Wang et al., 2013). Thus, temperatureis an important factor in regulating plant growth homeostasis bycontrolling the activation of some R or R-like proteins.
Several key components have been shown to be geneticallyrequired for R gene-dependent defense activation in Arabidopsis.The different requirements for the two important components,EDS1 (ENHANCED DISEASE SUSCEPTIBILITY 1) andNDR1 (NON-EXPRESSOR OF PR GENEs 1) (Century et al.,1995; Parker et al., 1996), in pathogen defense define two majorR gene-mediated signaling pathways in Arabidopsis (Aarts et al.,1998; McDowell et al., 2000; Bittner-Eddy & Beynon, 2001; Liet al., 2001; Borhan et al., 2004). Further studies have identifiedadditional components that function in SA-dependent signalingduring disease resistance in Arabidopsis. PAD4 (PHYTOALEXINDEFICIENT 4) and SAG101 (SENESCENCE-ASSOCIATEDGENE101) interact individually with EDS1 to form several spa-tially distinct complexes to activate SA signaling (Feys et al.,2001; Wagner et al., 2011; Zhu et al., 2011). The protein com-plex SGT1-RAR1-HSP90 (SUPPRESSOR OF G-TWOALLELE OF Skp1-REQUIRED FOR Mla12 RESISTANCE-HEAT SHOCK PROTEIN 90), acting as a chaperone, isrequired for the defense mediated by several R genes, includingRPM1 (RESISTANCE TO PSEUDOMONAS MACULICOLA1), RPS2 (RIBOSOMAL PROTEIN S2) and RPS4 in Arabidopsis(Hubert et al., 2003; Takahashi et al., 2003; Zhang et al., 2004),N in Nicotiana benthamiana (Liu et al., 2004) and Rx in potato(Boter et al., 2007).
Our previous study showed that a gain-of-function mutant ofa TIR-NB-LRR-type R protein RPP4, chs2-1/rpp4-1d, leads tocell death during chilling stress. The rpp4-1d -conferred chillingsensitivity required EDS1, RAR1 and SGT1b, but did not requirePAD4 or SA. In this study, we report a genetic screen for rpp4-1dsuppressors. We show that mutants which carry point mutationsin two isoforms of the cytosolic heat shock protein HSP90,namely HSP90.2 and HSP90.3, suppress the rpp4-1d-associated
phenotypes at chilling temperatures, and HSP90 interacts withRPP4/rpp4. Therefore, HSP90 acts as an important chaperone tomodulate RPP4-mediated temperature-dependent cell death anddefense responses.
Materials and Methods
Plant material and growth conditions
Arabidopsis thaliana (L.) Heynh. plants of the Columbia (Col-0)and Landsberg erecta (Ler) accessions were used in this study.The mutants used in this study were hsp90.3-3, mos1-5 (Li et al.,2010), mos2-2 (Zhang et al., 2005), mos3-2 (Zhang & Li, 2005),mos4-2 (Palma et al., 2007), mos6-4 (Palma et al., 2005), mos7-1(Cheng et al., 2009) and wrky70 (Knoth et al., 2007). Plants weregrown under a 16-h light (100 lmol m�2 s�1) : 8-h dark regimeat 22 or 16°C in the soil or on an agar plate containing Murashigeand Skoog medium (Sigma).
Genetic mapping and cloning of the SUCH1 (SUPPRESSOROF CHS2-1) genes
The suppressors of chs2-1/rpp4-1d were screened and mapped asdescribed previously (Huang et al., 2010). Approximately 500homozygous or heterozygous rpp4-1d plants with wild-type mor-phology at 4°C were used for mapping. The candidate genes werePCR amplified and the mutations were identified from themutant plants.
Plasmid construction and plant transformation
A 4.4-kb genomic fragment containing the HSP90.2 gene wasamplified by PCR using the HSP90.2-p1F and HSP90.2-p1Rprimers (Supporting Information Table S1), and cloned into thepCAMBIA1300 vector to generate pHSP90.2:HSP90.2. A 3.2-kb genomic fragment containing the HSP90.3 gene was ampli-fied using the HSP90.3-p1F and HSP90.3-p1R primers(Table S1), and cloned into pCAMBIA1300 to generatepHSP90.3:HSP90.3.
The HSP90.3 and hsp90.3 cDNAs were amplified by PCRfrom wild-type and hsp90.3-1 mutant plants using the primersHSP90.3-p2F and HSP90.3-p2R (Table S1). Both fragmentswere then cloned into pSuper1300 (Yang et al., 2010) with aSuper promoter (Ni et al., 1995) to generate pSuper:HSP90.3and pSuper:hsp90.3.
The Agrobacterium tumefaciens-mediated transformation wasperformed according to the floral dip method (Clough & Bent,1998).
Genetic analysis
To generate double mutants, rpp4-1d was crossed with thewrky70, mos1-5, mos2-2, mos3-2, mos4-2, mos6-4 and mos7-1mutant lines. The F2 progeny were specifically genotyped, andthe homozygosity of the rpp4-1d mutation was identified asdescribed by Huang et al. (2010).
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2

Ion leakage, proline (Pro) content and SA measurement
The electrolyte leakage assay was performed as described by Leeet al. (2002). Pro content was measured as described by Bateset al. (1972). Free SA and total SA were extracted from 3-wk-oldplants grown at different temperatures and measured as describedpreviously (Huang et al., 2010).
Chemical treatments
Trypan blue staining and 3,30-diaminobenzidine (DAB) stainingwere performed as described previously (Bowling et al., 1997;Thordal-Christensen et al., 1997). For HSP90 inhibitor treat-ment, the true leaves were injected with 10 lM geldanamycin(GDA) (Sigma) and the same volume of DMSO was used asa negative control. The phenotype was observed at day 7 aftertreatment.
Co-immunoprecipitation assay
The wild-type and mutated forms of RPP4 were cloned into pSu-per1300 containing the Myc tag. The wild-type and mutatedforms of HSP90.3 were cloned into pSuper1300 containing thegreen fluorescent protein (GFP) tag. The purified plasmids weretransformed into Arabidopsis mesophyll protoplasts following aprevious protocol (Zhai et al., 2009). After overnight incubation,the protoplasts were lysed and the RPP4/rpp4-Myc proteins wereimmunoprecipitated by anti-Myc agarose conjugate (Sigma). Theco-immunoprecipitation products were detected by immunoblotanalysis with anti-HSP90 antibody (Abmart, Shanghai, China).
Firefly luciferase (Luc) complementation imaging (LCI)assay
HSP90.3/hsp90.3 was fused with N-Luc in the pCAMBIA-nLUC vector, and the TIR, NB-ARC and LRR domains of theRPP4 protein were fused with C-Luc in the pCAMBIA-cLUCvector. The vectors were transformed into N. benthamiana leavesusing an Agrobacterium-mediated method. After 72 h of infiltra-tion, Luc activity was observed as described previously (Chenet al., 2008).
Protein fractionation and immunoblot assays
The protoplasts transformed with rpp4-Myc were treated at 4 or22°C for the indicated time. Soluble and nuclear proteins fromprotoplasts were isolated using a Plant Nuclei Isolation/Extrac-tion Kit (Sigma). HSP90 protein was used as a cytosolic marker,and histone H3 was used as a nuclear marker. Nuclear proteinextracts (N) were 99 concentrated compared with soluble frac-tions (S). The rpp4-Myc fusion protein was detected using ananti-Myc antibody (Sigma). HSP90 protein was detected usingan anti-HSP90 antibody. The accumulation of nuclear RPP4protein was analyzed based on the N/S ratio of RPP4. Theexperiments were repeated three times, and a representative figureis shown.
Bacterial growth assay
A pathogen resistance test on Pseudomonas syringae pv tomato(Pst) strain DC3000 was performed as described previously (Kim& Delaney, 2002). The pathogen concentrations wereOD600 = 0.02 for virulent strain Pst DC3000 and OD600 = 0.2for avirulent strains Pst DC3000 with avrRpm1, avrRpt2 andavrRps4. Four-week-old plants were dipped into the bacterial sus-pension containing 10 mM MgCl2 and 0.025% Silwet L-77.The leaves were collected 2 h after inoculation and at 3 d post-inoculation. For each time point, three replicate samples werecollected to determine the susceptibility of Pst DC3000.
A pathogen resistance test on Peronospora parasitica Emwa1was performed as described by Yang & Hua (2004). Twelve-day-old seedlings were sprayed at a concentration of 105 spores ml�1
of water. The oomycete spores on the leaf surface were counted10 d after inoculation.
Results
Identification of the such1mutants
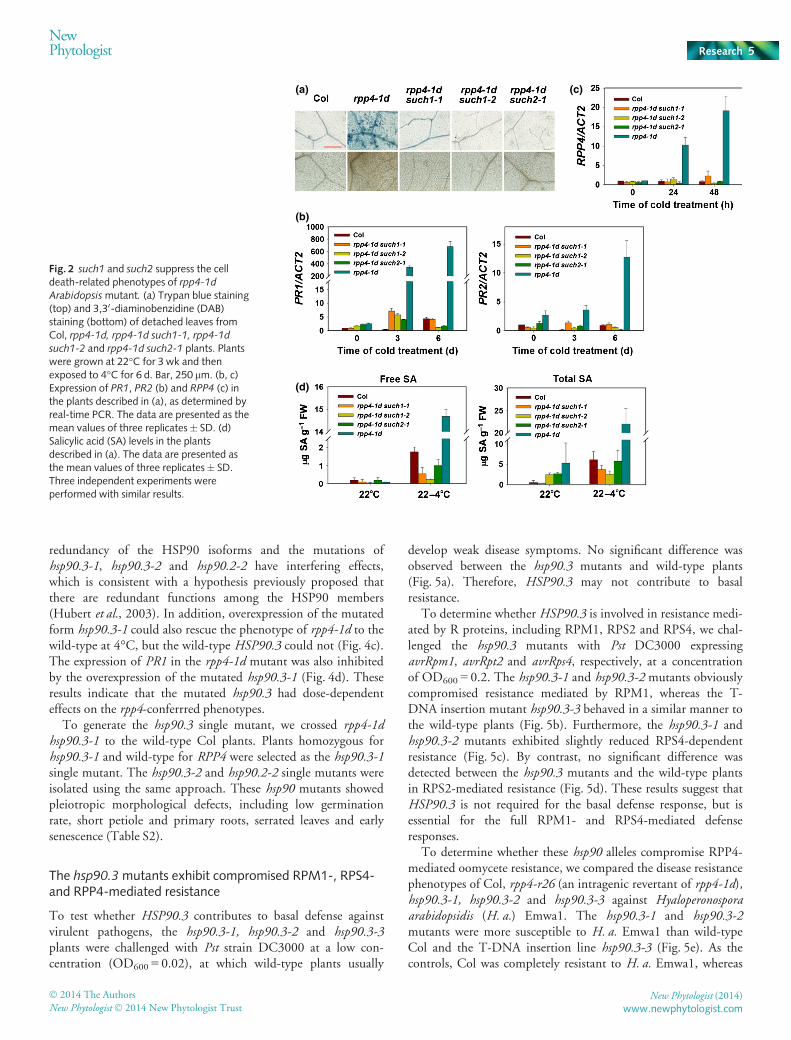
Our previous study showed that a gain-of-function mutant ofRPP4, chs2-1/rpp4-1d, confers sensitivity to low temperature(Huang et al., 2010). To identify new components involved inthe RPP4-mediated pathway, the such (suppressor of chs2-1)mutants were screened from an ethane methyl sulfonate (EMS)-mutagenized pool of rpp4-1d plants, as described previously(Huang et al., 2010). The rpp4-1d plants have a phenotype ofyellow, wilted leaves at chilling temperatures (Huang et al.,2010), and the suppressor screening was performed to isolatemutants that reverted to the wild-type morphology under chillingconditions. We identified two allelic mutants of such1, such1-1and such1-2, and one such2 mutant. The such1 and such2 mutantsreverted to the sensitive phenotype of the rpp4-1d mutant at both4 and 16°C (Fig. 1a). Ion leakage is an indicator of the integrityof the plasma membrane. The increased ion leakage observed inrpp4-1d plants at 4°C was significantly suppressed by the such1and such2 mutations (Fig. 1b). Accumulation of Pro, an impor-tant osmolyte, was dramatically reduced in the rpp4-1d mutantunder cold stress (Huang et al., 2010). To determine whether thesuch1 or such2 mutation affected Pro accumulation in rpp4-1d,Pro was extracted from the rpp4-1d such1 and rpp4-1d such2 dou-ble mutants and measured. The Pro levels in the rpp4-1d such2and rpp4-1d such2 double mutants were significantly increasedcompared with those in rpp4-1d (Fig. 1c).
The cell death-related phenotypes are suppressed by such1and such2
A previous study showed that low temperatures induced exten-sive cell death in the rpp4-1d mutant (Huang et al., 2010).However, cell death was dramatically suppressed in the rpp4-1dsuch1 and rpp4-1d such2 double mutants following 4°C treat-ment (Fig. 2a). Moreover, the accumulation of hydrogen perox-ide (H2O2), as revealed by DAB staining, was obviously
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 3

inhibited in the rpp4-1d such1 and rpp4-1d such2 doublemutants at 4°C (Fig. 2a).
In the rpp4-1d plants, the RPP4 and PR genes were up-regu-lated after cold treatment (Huang et al., 2010). However, expres-sion of RPP4, PR1 and PR2 was suppressed in the rpp4-1d such1and rpp4-1d such2 double mutants relative to rpp4-1d (Fig. 2b,c).The SA levels were elevated in the rpp4-1d mutant following coldtreatment (Huang et al., 2010). As shown in Fig. 2(d), the levelsof free and total SA in the double mutants were drasticallyreduced compared with those in rpp4-1d. Therefore, such1 and
such2 largely suppressed all of the known rpp4-conferred pheno-types under chilling stress.
Map-based cloning of SUCH1 and SUCH2
To examine the nature of the such1 mutation, rpp4-1d such1 wasbackcrossed to rpp4-1d, and the F1 progeny exhibited the rpp4-conferred chilling sensitivity. Of 200 F2 plants, 156 exhibitedrpp4-like morphology (expected ratio is three of four). Theseresults indicate that such1 is a single, recessive nuclear mutation.
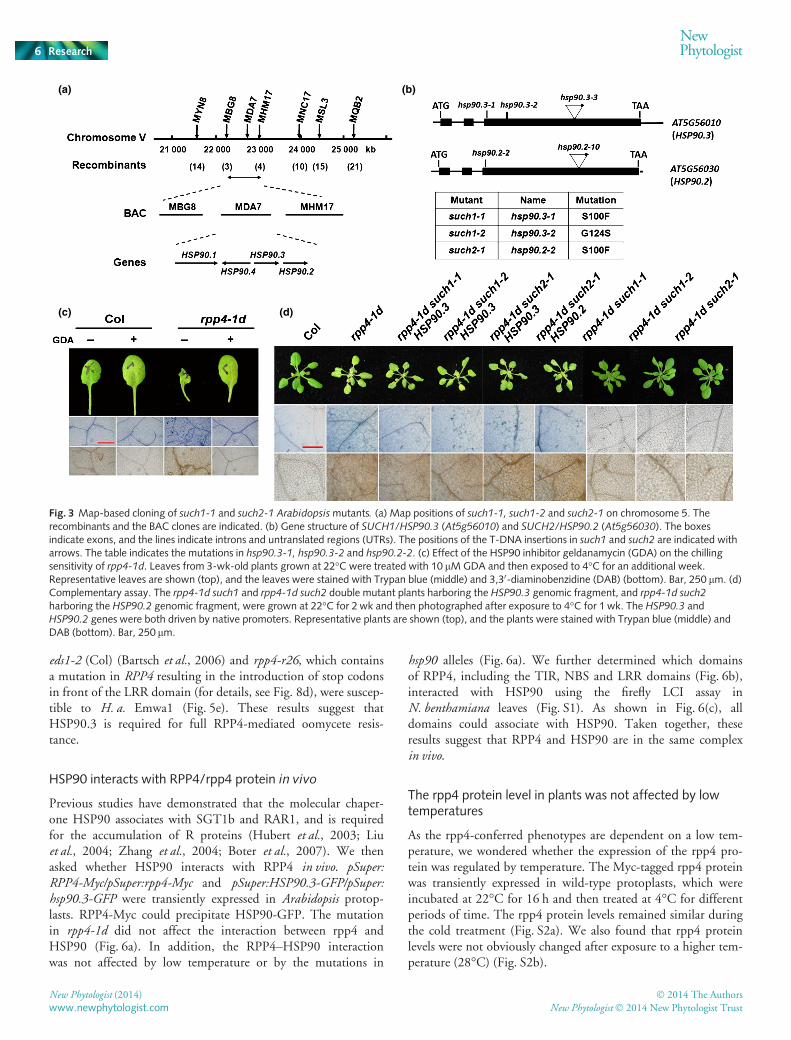
A map-based cloning approach was used to identify the muta-tion in such1 that suppressed the rpp4-1d phenotypes. The rpp4-1d such1-1 double mutant in the Col ecotype was crossed withLer. At 4°C, wild-type-looking plants that were homozygous orheterozygous at the rpp4-1d locus in the F2 progeny were usedfor rough mapping. The such1-1 locus was found to have linkagewith markers on the bottom arm of chromosome 5. Further finemapping using 201 F2 mutant plants that harbored the rpp4-1dmutation narrowed the location of the such1-1 mutation to an843-kb region between the MBG8 and MHM17 markers onchromosome 5 (Fig. 3a). Sequencing analysis of this regionrevealed a C299T transition in At5g56010 (HSP90.3), whichresulted in the S100F substitution (Fig. 3b). In such1-2, a G to Atransition at nucleotide 370 resulted in the G124S mutation inthe same gene. Although genetic analysis showed that such1 andsuch2 were at the same locus (see later for details), there was nomutation in the At5g56010 gene in the rpp4-1d such2-1 doublemutants. In Arabidopsis, there are five genes encoding HSP90,and four of these, including HSP90.3, are arranged in tandem.Therefore, we sequenced the other three HSP90 genes in therpp4-1d such2-1 double mutant and found a point mutation inthe third exon of At5g56030 that resulted in the mutation of the100th amino acid (S to F), which was previously named hsp90.2-2 (Hubert et al., 2003).
To examine whether the mutations in the hsp90 genes wereresponsible for the suppression of the rpp4-1d phenotypes, theHSP90 inhibitor GDA was used. Application of GDA dramati-cally inhibited the chilling sensitivity, cell death and H2O2 accu-mulation of rpp4-1d under chilling stress (Fig. 3c). Next, thewild-type fragments of At5g56010/HSP90.3 and At5956030/HSP90.2, under the control of their own promoters, were trans-formed into rpp4-1d such1 and rpp4-1d such2-1, respectively. Alltransformed plants restored the rpp4-conferred phenotypes underchilling stress, indicating that SUCH1 is HSP90.3, and SUCH2is HSP90.2 (Fig. 3d). Furthermore, the rpp4-1d such2-1 pheno-types were also suppressed by HSP90.3 (Fig. 3d), suggesting theredundant function of HSP90.2 and HSP90.3. For concisenessand consistency, the mutant such1-1 is hereafter named hsp90.3-1,such1-2 is hsp90.3-2 and such2-1 is hsp90.2-2.
We also obtained two T-DNA insertion lines, designatedhsp90.3-3 and hsp90.2-10, from ABRC. However, the hsp90.3-3and hsp90.2-10 loss-of-function mutants could not rescue thephenotype of rpp4-1d under chilling stress (Fig. 4a). In addition,we found that the F1 progeny of rpp4-1d hsp90.3 and rpp4-1dhsp90.2-2 showed the same phenotypes as both of its parents at4°C (Fig. 4b). These results suggest that the functional
(a)
(b)
(c)
Fig. 1 such1 and such2 suppress the chilling sensitivity of rpp4-1dArabidopsismutant. (a) Morphology of soil-grown wild-type (Col), rpp4-1d, rpp4-1d such1-1, rpp4-1d such1-2 and rpp4-1d such2-1 plants atdifferent temperatures. Row 1 shows the plants grown at 22°C, whereasrow 2 shows the plants grown at 4°C for 1 wk after growth at 22°C for2 wk, and row 3 shows plants grown at 16°C for 3 wk. (b) The ion leakageof Col, rpp4-1d, rpp4-1d such1-1, rpp4-1d such1-2 and rpp4-1d such2-1
plants. Plants were grown at 22°C for 3 wk and then exposed to 4°C forthe indicated times. The data are presented as the mean values of fivereplicates� SD. (c) Proline content of the Col, rpp4-1d, rpp4-1d such1-1,
rpp4-1d such1-2 and rpp4-1d such2-1 plants. Plants were grown at 22°Cfor 3 wk and then exposed to 4°C for 6 d. The data are presented as themean values of three replicates� SD. *, P < 0.01 (Student’s t-test). Threeindependent experiments were performed with similar results.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4
redundancy of the HSP90 isoforms and the mutations ofhsp90.3-1, hsp90.3-2 and hsp90.2-2 have interfering effects,which is consistent with a hypothesis previously proposed thatthere are redundant functions among the HSP90 members(Hubert et al., 2003). In addition, overexpression of the mutatedform hsp90.3-1 could also rescue the phenotype of rpp4-1d to thewild-type at 4°C, but the wild-type HSP90.3 could not (Fig. 4c).The expression of PR1 in the rpp4-1d mutant was also inhibitedby the overexpression of the mutated hsp90.3-1 (Fig. 4d). Theseresults indicate that the mutated hsp90.3 had dose-dependenteffects on the rpp4-conferrred phenotypes.
To generate the hsp90.3 single mutant, we crossed rpp4-1dhsp90.3-1 to the wild-type Col plants. Plants homozygous forhsp90.3-1 and wild-type for RPP4 were selected as the hsp90.3-1single mutant. The hsp90.3-2 and hsp90.2-2 single mutants wereisolated using the same approach. These hsp90 mutants showedpleiotropic morphological defects, including low germinationrate, short petiole and primary roots, serrated leaves and earlysenescence (Table S2).
The hsp90.3mutants exhibit compromised RPM1-, RPS4-and RPP4-mediated resistance
To test whether HSP90.3 contributes to basal defense againstvirulent pathogens, the hsp90.3-1, hsp90.3-2 and hsp90.3-3plants were challenged with Pst strain DC3000 at a low con-centration (OD600 = 0.02), at which wild-type plants usually
develop weak disease symptoms. No significant difference wasobserved between the hsp90.3 mutants and wild-type plants(Fig. 5a). Therefore, HSP90.3 may not contribute to basalresistance.
To determine whether HSP90.3 is involved in resistance medi-ated by R proteins, including RPM1, RPS2 and RPS4, we chal-lenged the hsp90.3 mutants with Pst DC3000 expressingavrRpm1, avrRpt2 and avrRps4, respectively, at a concentrationof OD600 = 0.2. The hsp90.3-1 and hsp90.3-2 mutants obviouslycompromised resistance mediated by RPM1, whereas the T-DNA insertion mutant hsp90.3-3 behaved in a similar manner tothe wild-type plants (Fig. 5b). Furthermore, the hsp90.3-1 andhsp90.3-2 mutants exhibited slightly reduced RPS4-dependentresistance (Fig. 5c). By contrast, no significant difference wasdetected between the hsp90.3 mutants and the wild-type plantsin RPS2-mediated resistance (Fig. 5d). These results suggest thatHSP90.3 is not required for the basal defense response, but isessential for the full RPM1- and RPS4-mediated defenseresponses.
To determine whether these hsp90 alleles compromise RPP4-mediated oomycete resistance, we compared the disease resistancephenotypes of Col, rpp4-r26 (an intragenic revertant of rpp4-1d),hsp90.3-1, hsp90.3-2 and hsp90.3-3 against Hyaloperonosporaarabidopsidis (H. a.) Emwa1. The hsp90.3-1 and hsp90.3-2mutants were more susceptible to H. a. Emwa1 than wild-typeCol and the T-DNA insertion line hsp90.3-3 (Fig. 5e). As thecontrols, Col was completely resistant to H. a. Emwa1, whereas
(a) (c)
(b)
(d)
Fig. 2 such1 and such2 suppress the celldeath-related phenotypes of rpp4-1dArabidopsismutant. (a) Trypan blue staining(top) and 3,30-diaminobenzidine (DAB)staining (bottom) of detached leaves fromCol, rpp4-1d, rpp4-1d such1-1, rpp4-1d
such1-2 and rpp4-1d such2-1 plants. Plantswere grown at 22°C for 3 wk and thenexposed to 4°C for 6 d. Bar, 250 lm. (b, c)Expression of PR1, PR2 (b) and RPP4 (c) inthe plants described in (a), as determined byreal-time PCR. The data are presented as themean values of three replicates� SD. (d)Salicylic acid (SA) levels in the plantsdescribed in (a). The data are presented asthe mean values of three replicates� SD.Three independent experiments wereperformed with similar results.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 5
eds1-2 (Col) (Bartsch et al., 2006) and rpp4-r26, which containsa mutation in RPP4 resulting in the introduction of stop codonsin front of the LRR domain (for details, see Fig. 8d), were suscep-tible to H. a. Emwa1 (Fig. 5e). These results suggest thatHSP90.3 is required for full RPP4-mediated oomycete resis-tance.
HSP90 interacts with RPP4/rpp4 protein in vivo
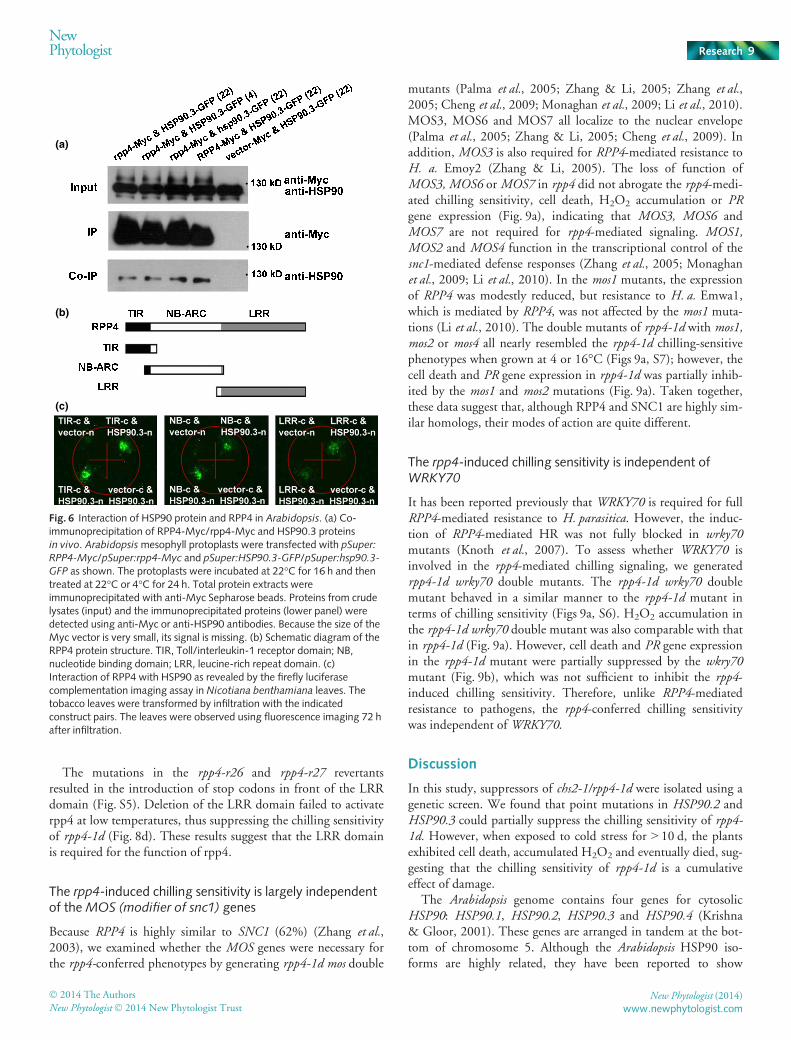
Previous studies have demonstrated that the molecular chaper-one HSP90 associates with SGT1b and RAR1, and is requiredfor the accumulation of R proteins (Hubert et al., 2003; Liuet al., 2004; Zhang et al., 2004; Boter et al., 2007). We thenasked whether HSP90 interacts with RPP4 in vivo. pSuper:RPP4-Myc/pSuper:rpp4-Myc and pSuper:HSP90.3-GFP/pSuper:hsp90.3-GFP were transiently expressed in Arabidopsis protop-lasts. RPP4-Myc could precipitate HSP90-GFP. The mutationin rpp4-1d did not affect the interaction between rpp4 andHSP90 (Fig. 6a). In addition, the RPP4–HSP90 interactionwas not affected by low temperature or by the mutations in
hsp90 alleles (Fig. 6a). We further determined which domainsof RPP4, including the TIR, NBS and LRR domains (Fig. 6b),interacted with HSP90 using the firefly LCI assay inN. benthamiana leaves (Fig. S1). As shown in Fig. 6(c), alldomains could associate with HSP90. Taken together, theseresults suggest that RPP4 and HSP90 are in the same complexin vivo.
The rpp4 protein level in plants was not affected by lowtemperatures
As the rpp4-conferred phenotypes are dependent on a low tem-perature, we wondered whether the expression of the rpp4 pro-tein was regulated by temperature. The Myc-tagged rpp4 proteinwas transiently expressed in wild-type protoplasts, which wereincubated at 22°C for 16 h and then treated at 4°C for differentperiods of time. The rpp4 protein levels remained similar duringthe cold treatment (Fig. S2a). We also found that rpp4 proteinlevels were not obviously changed after exposure to a higher tem-perature (28°C) (Fig. S2b).
(a) (b)
(c) (d)
Fig. 3 Map-based cloning of such1-1 and such2-1 Arabidopsismutants. (a) Map positions of such1-1, such1-2 and such2-1 on chromosome 5. Therecombinants and the BAC clones are indicated. (b) Gene structure of SUCH1/HSP90.3 (At5g56010) and SUCH2/HSP90.2 (At5g56030). The boxesindicate exons, and the lines indicate introns and untranslated regions (UTRs). The positions of the T-DNA insertions in such1 and such2 are indicated witharrows. The table indicates the mutations in hsp90.3-1, hsp90.3-2 and hsp90.2-2. (c) Effect of the HSP90 inhibitor geldanamycin (GDA) on the chillingsensitivity of rpp4-1d. Leaves from 3-wk-old plants grown at 22°C were treated with 10 lMGDA and then exposed to 4°C for an additional week.Representative leaves are shown (top), and the leaves were stained with Trypan blue (middle) and 3,30-diaminobenzidine (DAB) (bottom). Bar, 250 lm. (d)Complementary assay. The rpp4-1d such1 and rpp4-1d such2 double mutant plants harboring the HSP90.3 genomic fragment, and rpp4-1d such2
harboring the HSP90.2 genomic fragment, were grown at 22°C for 2 wk and then photographed after exposure to 4°C for 1 wk. The HSP90.3 andHSP90.2 genes were both driven by native promoters. Representative plants are shown (top), and the plants were stained with Trypan blue (middle) andDAB (bottom). Bar, 250 lm.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6

A previous study showed that the mutations in the ATP-bind-ing domain of HSP90 severely reduced RPM1 accumulation andthus compromised RPM1 function (Hubert et al., 2003).Because the mutations in the hsp90.2-2, hsp90.3-1 and hsp90.3-2mutants were all located in the conserved ATPase domain ofHSP90, it is possible that these mutations may affect the accumu-lation of the rpp4 protein at low temperatures. To test this possi-bility, we analyzed the rpp4 protein level at 4°C in protoplastsprepared from the hsp90 mutants. The rpp4 protein was notaffected by the hsp90.3-1, hsp90.3-2 or hsp90.2-2 mutation oncold stress (Fig. S2a). To further determine whether the hsp90.3mutation affects rpp4 protein stability, we expressed rpp4-Myc inprotoplasts of Col and hsp90.3 plants for 16 h at 22°C, followedby the addition of the protein synthesis inhibitor cycloheximide(CHX), and examined the protein levels of rpp4. As shown inFig. S3, the stability of the rpp4 protein was not influenced bythe mutation in hsp90.3. Taken together, these results indicatethat the rpp4 protein level is not regulated by temperature or thehsp90 mutations.
Nuclear accumulation of rpp4 is decreased at lowtemperatures
To further investigate the function of HSP90 in the rpp4-medi-ated chilling response, we investigated the subcellular localizationof rpp4 and RPP4 at 22 and 4°C. These proteins were fused toMyc at the C-terminus and were expressed in Arabidopsis protop-lasts. RPP4-Myc was mainly located in the cytoplasm andnucleus at 22 and 4°C according to immunoblot analysis(Fig. 7a). The mutated rpp4 protein was also located in both thecytoplasm and the nucleus at 22°C; however, the protein level ofrpp4 in the nucleus was dramatically decreased at 4°C relative to22°C (Fig. 7a).
To examine whether HSP90 mediates the decreased nuclearaccumulation of rpp4 at low temperatures, we treated the pro-toplasts expressing rpp4-Myc with 10 lM GDA. The decreasednuclear accumulation of rpp4 at 4°C was not affected by theapplication of GDA (Fig. 7b). In addition, rpp4-Myc wasexpressed in the protoplasts of wild-type Col and hsp90.3-2
(a)
(b) (c)
(d)
Fig. 4 Genetic analysis of rpp4-1d and different hsp90 alleles of Arabidopsis. Plants were grown at 22°C for 2 wk and then photographed after coldtreatment at 4°C for 1 wk. Representative plants are shown (top), and the plants were stained with Trypan blue (middle) and 3,30-diaminobenzidine (DAB)(bottom). Bar, 250 lm. (a) Phenotypes of the double mutants of rpp4-1d crossed to the hsp90 T-DNA insertion alleles. (b) Phenotypes of the F1 progenybetween two of rpp4-1d hsp90.3-1, rpp4-1d hsp90.3-2 and rpp4-1d hsp90.2-2. (c) Phenotypes of rpp4-1d harboring the hsp90.3-1mutant and HSP90.3wild-type genomic fragments. (d) PR expression in the plants described in (a) and (c). The data are presented as the mean values of three replicates� SD.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 7

plants for 16 h at 22°C which were then treated with coldstress. As shown in Fig. 7(c), the decrease in nuclear rpp4 wasnot compromised by the mutation in hsp90. Together, theseresults suggest that the suppression of the rpp4-conferrred phe-notype by hsp90.3 does not result from the altered localizationof rpp4.
To further dissect which domains are responsible for theentrance of rpp4 into the nucleus, we examined the subcellularlocalization of the TIR and NB-ARC domains of rpp4 inN. benthamiana leaves transiently expressing different domains ofRPP4. Immunoblot analysis showed that the TIR domain waslocalized in both the cytoplasm and the nucleus, whereas bothwild-type NB-ARC and mutated NB-ARC were only detected inthe nucleus (Fig. 7d). The expression of the LRR domain was toolow to be detected. These results suggest that the TIR andNB-ARC domains may function by coordinately regulating theratio of RPP4 in the cytoplasm and the nucleus.
Identification and characterization of intragenic suppressorsof rpp4-1d
We also identified 25 intragenic revertants of rpp4-1d (rpp4-r)from the M2 population. The chilling sensitivity of rpp4-1dwas suppressed in these rpp4-1d revertants when the tempera-ture was shifted from 22 to 4°C (Fig. S4a). The ion leakage ofthe rpp4-1d plants was also dramatically inhibited by thesemutations at 4°C (Fig. S4b). Sequencing analyses showed thatmost mutations of these suppressors were located in the NB-ARC domain, and a few mutations were located in the LRR
domain (Fig. S5). These results suggest that these amino acidsin the NB-ARC and LRR domains are required for the func-tion of RPP4.
In addition, we isolated intragenic suppressors resulting fromthe introduction of stop codons into RPP4, which we used tostudy the functions of the different domains of RPP4. We foundthat the deletion mutation in the N-terminus of the NB-ARCdomain of rpp4 repressed the chilling sensitivity of rpp4-1d; how-ever, this early terminated mutation activated the defenseresponse. For example, in rpp4-r25, the substitution of G to A atposition 933 resulted in the formation of a truncated proteincontaining only the TIR domain and part of the NB-ARCdomain (Fig. S3c). This mutation suppressed the chilling lethal-ity of rpp4-1d at 4°C (Fig. 8a). Intriguingly, this mutationresulted in extensive cell death and H2O2 accumulation at 22°C(Fig. 8b). However, the difference between rpp4-r25 and rpp4-1dwas that reactive oxygen species (ROS) accumulation and celldeath in the rpp4-r25 mutation was constitutive and independentof the temperature, and the mutant was not lethal, whereas rpp4-1d was very sensitive to temperature and exhibited lethality at4°C (Fig. 8a).
To further explore the function of the TIR domain of RPP4,we generated transgenic plants overexpressing the TIR domain inthe wild-type Col background (TIR-OE) (Fig. S6). The trans-genic plants exhibited a dwarf stature, obvious cell death andH2O2 accumulation, as well as up-regulated PR expression atboth 22 and 4°C, which is consistent with the phenotypes ofrpp4-r25 (Fig. 8a–c). Taken together, these results suggest thatthe complete TIR domain is sufficient to trigger weak HR.
(a) (b) (c)
(d) (e)
Fig. 5 The effect of the hsp90.3-1, hsp90.3-2 and hsp90.3-3 Arabidopsismutants on RPM1-, RPS4- and RPP4-mediated pathogen resistance inArabidopsis. (a) Response of the hsp90.3mutants to virulent bacteria. Four-week-old plants were dipped with Pseudomonas syringae pv tomato (Pst)DC3000 (OD600 = 0.02), and bacterial growth was measured as described. (b–d) Response of the hsp90.3mutants to avirulent bacteria. Four-week-oldplants were dipped with Pst DC3000 (avrRpm1) (b), Pst DC3000 (avrRpt2) (c) and Pst DC3000 (avrRps4) (d) at OD600 = 0.2. The data are presented as themean values of three replicates� SD. *, P < 0.05 (Student’s t-test). Three independent experiments were performed with similar results. (e) Quantificationof Hyaloperonospora arabidopsidis (H. a.) Emwa1 sporulation on the indicated genotypes. Twelve-day-old plants were sprayed with H. a. Emwa1 at aconcentration of 105 spores ml�1 of water. The oomycete spores on the leaf surface were counted 10 d later after inoculation. The data are presented asmeans� SD (n = 5 with four plants for each). Two independent experiments were performed with similar results.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8
The mutations in the rpp4-r26 and rpp4-r27 revertantsresulted in the introduction of stop codons in front of the LRRdomain (Fig. S5). Deletion of the LRR domain failed to activaterpp4 at low temperatures, thus suppressing the chilling sensitivityof rpp4-1d (Fig. 8d). These results suggest that the LRR domainis required for the function of rpp4.
The rpp4-induced chilling sensitivity is largely independentof theMOS (modifier of snc1) genes
Because RPP4 is highly similar to SNC1 (62%) (Zhang et al.,2003), we examined whether the MOS genes were necessary forthe rpp4-conferred phenotypes by generating rpp4-1d mos double
mutants (Palma et al., 2005; Zhang & Li, 2005; Zhang et al.,2005; Cheng et al., 2009; Monaghan et al., 2009; Li et al., 2010).MOS3, MOS6 and MOS7 all localize to the nuclear envelope(Palma et al., 2005; Zhang & Li, 2005; Cheng et al., 2009). Inaddition, MOS3 is also required for RPP4-mediated resistance toH. a. Emoy2 (Zhang & Li, 2005). The loss of function ofMOS3, MOS6 orMOS7 in rpp4 did not abrogate the rpp4-medi-ated chilling sensitivity, cell death, H2O2 accumulation or PRgene expression (Fig. 9a), indicating that MOS3, MOS6 andMOS7 are not required for rpp4-mediated signaling. MOS1,MOS2 and MOS4 function in the transcriptional control of thesnc1-mediated defense responses (Zhang et al., 2005; Monaghanet al., 2009; Li et al., 2010). In the mos1 mutants, the expressionof RPP4 was modestly reduced, but resistance to H. a. Emwa1,which is mediated by RPP4, was not affected by the mos1 muta-tions (Li et al., 2010). The double mutants of rpp4-1d with mos1,mos2 or mos4 all nearly resembled the rpp4-1d chilling-sensitivephenotypes when grown at 4 or 16°C (Figs 9a, S7); however, thecell death and PR gene expression in rpp4-1d was partially inhib-ited by the mos1 and mos2 mutations (Fig. 9a). Taken together,these data suggest that, although RPP4 and SNC1 are highly sim-ilar homologs, their modes of action are quite different.
The rpp4-induced chilling sensitivity is independent ofWRKY70
It has been reported previously that WRKY70 is required for fullRPP4-mediated resistance to H. parasitica. However, the induc-tion of RPP4-mediated HR was not fully blocked in wrky70mutants (Knoth et al., 2007). To assess whether WRKY70 isinvolved in the rpp4-mediated chilling signaling, we generatedrpp4-1d wrky70 double mutants. The rpp4-1d wrky70 doublemutant behaved in a similar manner to the rpp4-1d mutant interms of chilling sensitivity (Figs 9a, S6). H2O2 accumulation inthe rpp4-1d wrky70 double mutant was also comparable with thatin rpp4-1d (Fig. 9a). However, cell death and PR gene expressionin the rpp4-1d mutant were partially suppressed by the wkry70mutant (Fig. 9b), which was not sufficient to inhibit the rpp4-induced chilling sensitivity. Therefore, unlike RPP4-mediatedresistance to pathogens, the rpp4-conferred chilling sensitivitywas independent ofWRKY70.
Discussion
In this study, suppressors of chs2-1/rpp4-1d were isolated using agenetic screen. We found that point mutations in HSP90.2 andHSP90.3 could partially suppress the chilling sensitivity of rpp4-1d. However, when exposed to cold stress for > 10 d, the plantsexhibited cell death, accumulated H2O2 and eventually died, sug-gesting that the chilling sensitivity of rpp4-1d is a cumulativeeffect of damage.
The Arabidopsis genome contains four genes for cytosolicHSP90: HSP90.1, HSP90.2, HSP90.3 and HSP90.4 (Krishna& Gloor, 2001). These genes are arranged in tandem at the bot-tom of chromosome 5. Although the Arabidopsis HSP90 iso-forms are highly related, they have been reported to show
(a)
(b)
(c)
Fig. 6 Interaction of HSP90 protein and RPP4 in Arabidopsis. (a) Co-immunoprecipitation of RPP4-Myc/rpp4-Myc and HSP90.3 proteinsin vivo. Arabidopsismesophyll protoplasts were transfected with pSuper:RPP4-Myc/pSuper:rpp4-Myc and pSuper:HSP90.3-GFP/pSuper:hsp90.3-GFP as shown. The protoplasts were incubated at 22°C for 16 h and thentreated at 22°C or 4°C for 24 h. Total protein extracts wereimmunoprecipitated with anti-Myc Sepharose beads. Proteins from crudelysates (input) and the immunoprecipitated proteins (lower panel) weredetected using anti-Myc or anti-HSP90 antibodies. Because the size of theMyc vector is very small, its signal is missing. (b) Schematic diagram of theRPP4 protein structure. TIR, Toll/interleukin-1 receptor domain; NB,nucleotide binding domain; LRR, leucine-rich repeat domain. (c)Interaction of RPP4 with HSP90 as revealed by the firefly luciferasecomplementation imaging assay in Nicotiana benthamiana leaves. Thetobacco leaves were transformed by infiltration with the indicatedconstruct pairs. The leaves were observed using fluorescence imaging 72 hafter infiltration.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 9

(a)
(b)
(c)
(d)
Fig. 7 Immunoblot analysis of rpp4/RPP4protein in Arabidopsis under cold stress.Protoplasts expressing PRR4-Myc and rpp4-Myc were incubated at 22°C for 16 h andthen treated at 22 or 4°C for 24 h. Total (T),soluble (S) and nuclear (N) proteins wereextracted from the protoplasts and subjectedto immunoblot analysis with antibodiesagainst Myc and HSP90. HSP90 protein wasused as a cytosolic marker and histone H3was used as a nuclear marker. Nuclearprotein extracts (N) were 99 (a–c) and 149(d) concentrated compared with the solublefractions (S). The accumulation of nuclearRPP4 protein was analyzed based on theratio of N/S of RPP4. (a) Immunoblotanalysis of RPP4, rpp4 and HSP90 at 22 and4°C. (b) The effect of geldanamycin (GDA)on rpp4 nuclear accumulation at 22 and 4°C.(c) The effect of hsp90mutation on rpp4nuclear accumulation at 22 and 4°C. (d)Localization of the Toll/interleukin-1 receptor(TIR) and nucleotide binding (NB) domains ofthe RPP4/rpp4 protein.
(a)
(b)
(c)
(d)
Fig. 8 Characterization of the Toll/interleukin-1 receptor (TIR) and leucine-rich repeat (LRR) domains of Arabidopsis RPP4. (a) Phenotypes of the rpp4-r25and TIR-overexpressing plants grown at 22°C for 2 wk and then exposed to 4°C for 1 wk. Representative plants are shown (top), and the plants werestained with Trypan blue (middle) and 3,30-diaminobenzidine (DAB) (bottom). Bar, 250 lm. (b) Phenotypes of the rpp4-r25 and TIR-overexpressing plantsgrown at 22°C for 3 wk. Representative plants are shown (top), and the plants were stained with trypan blue (middle) and DAB (bottom). Bar, 250 lm. (c)Expression of the PR genes in the TIR-overexpressing plants grown on MS plates at 22°C for 2 wk and then treatment at 4�C for the indicated times, asrevealed by quantitative reverse transcription-polymerase chain reaction (qRT-PCR). The data are presented as the mean values of three replicates� SD.(d) Phenotypes of the rpp4-r25 and rpp4-r26 plants grown at 22°C for 2 wk and then photographed after exposure to 4°C for 1 wk. Representative plantsare shown (top), and the plants were stained with trypan blue (middle) and DAB (bottom). Bar, 250 lm.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10

different expression profiles (Prasinos et al., 2005). In addition,their loss-of-function mutants display developmental defectswith some variation (Samakovli et al., 2007). Consistently, wenoticed that point mutations in HSP90.2 and HSP90.3 alsoshowed different, mild morphological alterations, indicating thatthey played both redundant and distinct roles in plant develop-ment. In terms of the stress responses, HSP90.1 is the only cyto-solic HSP90 in Arabidopsis to be significantly induced by PstDC3000. HSP90.1 interacts with RAR1 and SGT1 and is criti-cal for RPS2-mediated disease resistance (Takahashi et al.,2003). In this study, we showed that at least HSP90.2 andHSP90.3 are involved in rpp4-mediated chilling sensitivity,because their mutations could compromise the rpp4-inducedchilling sensitivity. However, the T-DNA insertion lines ofhsp90.2 and hsp90.3 could not rescue the rpp4-1d phenotype,suggesting the functional redundancy of HSP90. The geneticanalysis showed that the F1 progeny of hsp90.2-2 crossed tohsp90.3-1 or hsp90.3-2 suppressed the rpp4-1d phenotype,
which is indicative of dosage dependence. That is, the inhibitionof the rpp4-1d phenotype only occurred in the presence of atleast two copies of the mutated HSP90 genes in the rpp4-1dmutant. This was further confirmed in transgenic plants overex-pressing a mutated form of hsp90.3.
A previous study has shown that virus-induced gene silencingof HSP90.1 affects cell death in tobacco caused by RPS4 overex-pression (Zhang et al., 2004). Consistent with this, we found thathsp90.3-1 and hsp90.3-2 exhibited weak susceptibility to PstDC3000 harboring avrRPS4 and avrRPM1. Furthermore,HSP90.3 was shown to be essential for full RPP4-dependentpathogen resistance. These data suggest that RPS4-, RPM1- andRPP4-mediated pathogen resistance is dependent on differentHSP90 isoforms. It is also possible that different R proteins mayrequire different quantities of HSP90s.
R proteins are located in different subcellular locations, includ-ing the cytoplasm, nucleus, plasma membrane, endoplasmicreticulum and chloroplasts, and their localization is tightly linked
(a)
(b)Fig. 9 Phenotypes of the rpp4-1d mos andrpp4-1d wrky70 Arabidopsis double mutantsat 4°C. (a) Plants were grown at 22°C for1 wk and then photographed after exposureto 4°C for 1 wk. Representative plants areshown (top), and the plants were stainedwith Trypan blue (middle) and 3,30-diaminobenzidine (DAB) (bottom). Bars,250 lm. (b) Expression of PR1 in the plantsdescribed in (a), as determined by real-timePCR. The data are presented as the meanvalues of three replicates� SD.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 11

to their functions. For instance, the nuclear accumulation of Rproteins, such as N, MLA (mildew-resistance locus A) and RPS4,is essential for the triggering of pathogen resistance (Burch-Smithet al., 2007; Shen et al., 2007; Wirthmueller et al., 2007). Inaddition, snc1-mediated temperature-dependent cell death isassociated with the nuclear accumulation of SNC1 (Y. Zhu et al.,2010). A previous study has reported that nuclear processes ofavrRps4 are essential for bacterial growth restriction. The pro-grammed cell death and transcriptional defense amplificationrequire nucleocytoplasmic coordination of avrRps4. EDS1 couldinteract with RPS4 and avrRps4 in the nucleus, respectively.However, when the defense response is activated, the interactionsbetween EDS1 and RPS4 or avrRps4 are only detected in thecytoplasm. This study thus proposes that the effector triggers dis-tinct, but coordinated, subcellular defense branches through anRPS4–EDS1 complex that can accumulate in the cytoplasm andnucleus (Heidrich et al., 2011). A study of the barley MLA pro-tein showed that the nuclear MLA10 pool alone cannot inducecell death, but is sufficient to trigger disease resistance. Enhancedaccumulation of MLA10 in the cytoplasm promotes cell deathactivity, but compromises MLA10-mediated disease resistance(Bai et al., 2012). In this study, we found that the rpp4-mediatedtemperature-dependent cell death was accompanied by a decreasein the nuclear accumulation of the rpp4 protein. It is possiblethat the decrease in the nuclear pool of rpp4 may account for thiscell death phenotype. It is also possible that the distribution ofrpp4 protein between the cytoplasm and nucleus changes, whenchilling activates the defense response in the rpp4-1d mutant,similar to that which occurs when the effector is recognized bythe R protein. However, attempts to generate transgenic plantsexpressing rpp4 protein fused to a nuclear localization sequence(NLS) or a nuclear export sequence (NES) were unsuccessful.Therefore, we could not test whether the decrease in nuclear rpp4was the reason for the rpp4-conferred chilling sensitivity.
Emerging evidence supports the notion that the activation ofR proteins is modulated by temperature (Alcazar & Parker,2011; Hua, 2013). Our previous study showed that one aminoacid change in rpp4-1d caused its sensitivity to chilling tempera-tures (Huang et al., 2010). Genetic analysis of intragenic suppres-sors revealed that the TIR domain of RPP4 alone could activatedefense signaling, although this defense response was not asstrong as the rpp4-activated response. The phenotype of therpp4-1d mutant is blocked when only the TIR-NB-ARCdomains are present, implying that NB-ARC may inhibit theactivation of the TIR domain. This also suggests that the LRRdomain is required to maintain the chilling response of rpp4-1d.The N-terminal TIR or coiled coil (CC) domain of the R proteinis thought to be a defense signaling domain, whose activity iscontrolled by other domains (Swiderski et al., 2009; Bernouxet al., 2011; Maekawa et al., 2011). Consistent with our study,overexpression of the TIR domain of RPS4 in N. benthamianaleaves can trigger cell death (Zhang et al., 2004; Swiderski et al.,2009). Transient expression of the CC domain of MLA10 dis-plays auto-active cell death (Bai et al., 2012). Previous studieshave reported that the LRR domain has dual functions in theregulation of the Rx protein through its interaction with
CC-NB-ARC. Interference with the intramolecular interactionsby the introduction of point mutations in the NB-ARC or LRRdomain can activate the Rx protein (Moffett et al., 2002; Rairdan& Moffett, 2006). This indicates that NB-ARC and LRR mayplay a negative role in controlling the activity of the Rx protein.However, deletion of the LRR domain fails to induce HRalthough the elicitor exists, suggesting that the LRR domain isnecessary for the full activation of the integrated Rx protein. Fur-thermore, mutations in the LRR of SNC1 confer activity at hightemperature, whereas further mutations in NB-ARC abolish thehigh-temperature but not low-temperature activity (Y. Zhu et al.,2010). Together with our analysis on the intragenic suppressorsof rpp4-1d, these results suggest that specific interaction betweenNB-ARC and LRR is important for temperature-modulatedactivity.
The wrky70 loss-of-function mutant failed to induce resistanceto H. a. Emoy2, and could not fully block the induction ofRPP4-mediated HR (Knoth et al., 2007), suggesting thatWRKY70 may be a component that is mainly required in RPP4-mediated pathogen resistance, but not in the cell death pathway.Here, we showed that, although the expression of PR genes waspartially repressed, the rpp4-conferred chilling sensitivity was notsuppressed by the wrky70 mutant, which might be a result of thefunctional redundancy of WRKY genes. It is conceivable that therpp4-mediated chilling sensitivity is similar to HR induced bypathogen infection. HR induced by pathogen infection onlyoccurs in a few of the cells that directly contact the pathogen.Different from pathogen-induced HR, rpp4-induced HR pre-sumably happens in the whole plant, thus leading to extensive celldeath of whole plants.
Fig. 10 A proposed model for RPP4-mediated temperature-dependent celldeath and defense responses controlled by HSP90 in Arabidopsis. TheRPP4 protein has two states, resting (RPP4-r) and active (RPP4-a), whichare dependent on the interactions among different domains. Thepathogen or mutation in RPP4 which suppresses the interactions couldtransfer RPP4 from the resting state to the active state. Under chillingstress, facilitated by the SGT1-RAR1-HSP90 complex, the mutated rpp4protein is activated and triggers EDS1-dependent cell death and WRKY70-dependent defense signaling. The mutated hsp90 protein has a dose-dependent effect on interfering wild-type HSP90-mediated downstreamsignaling. TIR-NB-ARC truncated protein cannot, but TIR domain alonecan, partially activate the cell death and defense response at normal andchilling temperatures.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist12

Extensive studies of the mos mutants revealed that SNC1-mediated signaling requires nucleocytoplasmic trafficking, tran-scriptional regulation, RNA processing and protein modification(Palma et al., 2005; Zhang et al., 2005; Zhang & Li, 2005; Gori-tschnig et al., 2008; Palma et al., 2007; Cheng et al., 2009; Liet al., 2010; Z. Zhu et al., 2010; Xu et al., 2011). RPP4 andSNC1 are close homologs located at the RPP5 locus and arecoordinately regulated (Yi & Richards, 2007). Nevertheless, ourgenetic analysis demonstrated that the rpp4-induced chilling sen-sitivity was not dependent on these MOS genes. Moreover,SGT1b and RAR1 are not required for snc1 (Goritschnig et al.,2007), but are required for rpp4. These results suggest that rpp4and snc1 modulate temperature-dependent defense responsesthrough distinct mechanisms.
In summary, we propose a working model for RPP4-mediatedtemperature-dependent cell death and defense responses con-trolled by HSP90 (Fig. 10). RPP4 protein maintains a restingstate as a result of its intramolecular interactions at normaltemperature. However, pathogen attack and mutations in theNB-ARC domain of RPP4 (rpp4-1d) could activate the RPP4protein at normal and chilling temperatures, respectively, via achange in its intramolecular interactions facilitated by theSGT1b-RAR1-HSP90 complex, which, in turn, leads to EDS1-and WRKY70-dependent cell death and the defense response. Asa result, continuous chilling stress-induced HR causes chillingsensitivity of plants. Although the mutated hsp90 can form acomplex with RPP4/rpp4, it has a dose-dependent interferingeffect on the wild-type HSP90, which consequently preventsRPP4/rpp4 from moving from the resting state to the active state.TIR-NB-ARC fails to activate defense responses, whereas theTIR domain alone released from the inhibition of NB-ARC canactivate cell death and the defense response at both normal andchilling temperatures.
Acknowledgements
This work was supported by China National Funds for Distin-guished Young Scientists (31225003) and the National NaturalScience Foundation of China (31121002 and 31330006).
References
Aarts N, Metz M, Holub E, Staskawicz BJ, Daniels MJ, Parker JE. 1998.
Different requirements for EDS1 and NDR1 by disease resistance genes define
at least two R gene-mediated signaling pathways in Arabidopsis. Proceedings ofthe National Academy of Sciences, USA 95: 10306–10311.
Alcazar R, Garcia AV, Parker JE, Reymond M. 2009. Incremental steps toward
incompatibility revealed by Arabidopsis epistatic interactions modulating
salicylic acid pathway activation. Proceedings of the National Academy of Sciences,USA 106: 334–339.
Alcazar R, Parker JE. 2011. The impact of temperature on balancing immune
responsiveness and growth in Arabidopsis. Trends in Plant Science 16: 666–675.Bai S, Liu J, Chang C, Zhang L, Maekawa T, Wang Q, Xiao W, Liu Y, Chai J,
Takken FL et al. 2012. Structure–function analysis of barley NLR immune
receptor MLA10 reveals its cell compartment specific activity in cell death and
disease resistance. PLoS Pathogens 8: e1002752.Bartsch M, Gobbato E, Bednarek P, Debey S, Schultze JL, Bautor J, Parker JE.
2006. Salicylic acid-independent ENHANCED DISEASE
SUSCEPTIBILITY1 signaling in Arabidopsis immunity and cell death is
regulated by the monooxygenase FMO1 and the nudix hydrolase NUDT7.Plant Cell 18: 1038–1051.
Bates LS, Waldren RP, Teare ID. 1972. Rapid determination of free proline for
water-stress studies. Plant and Soil 39: 205–207.Bernoux M, Ve T, Williams S, Warren C, Hatters D, Valkov E, Zhang X, Ellis
JG, Kobe B, Dodds PN. 2011. Structural and functional analysis of a plant
resistance protein TIR domain reveals interfaces for self-association, signaling,
and autoregulation. Cell Host & Microbe 9: 200–211.Bittner-Eddy PD, Beynon JL. 2001. The Arabidopsis downy mildew resistance
gene, RPP13-Nd, functions independently of NDR1 and EDS1 and does not
require the accumulation of salicylic acid.Molecular Plant–Microbe Interactions14: 416–421.
Bomblies K, Lempe J, Epple P, Warthmann N, Lanz C, Dangl JL, Weigel D.
2007. Autoimmune response as a mechanism for a Dobzhansky–Muller-type
incompatibility syndrome in plants. PLoS Biology 5: e236.Borhan MH, Holub EB, Beynon JL, Rozwadowski K, Rimmer SR. 2004. The
Arabidopsis TIR-NB-LRR gene RAC1 confers resistance to Albugo candida(white rust) and is dependent on EDS1 but not PAD4.Molecular Plant–Microbe Interactions 17: 711–719.
Boter M, Amigues B, Peart J, Breuer C, Kadota Y, Casais C, Moore G,
Kleanthous C, Ochsenbein F, Shirasu K et al. 2007. Structural and functional
analysis of SGT1 reveals that its interaction with HSP90 is required for the
accumulation of Rx, an R protein involved in plant immunity. Plant Cell 19:3791–3804.
Bowling SA, Clarke JD, Liu Y, Klessig DF, Dong X. 1997. The cpr5mutant of
Arabidopsis expresses both NPR1-dependent and NPR1-independent
resistance. Plant Cell 9: 1573–1584.Burch-Smith TM, Schiff M, Caplan JL, Tsao J, Czymmek K, Dinesh-Kumar SP.
2007. A novel role for the TIR domain in association with pathogen-derived
elicitors. PLoS Biology 5: e68.Century KS, Holub EB, Staskawicz BJ. 1995. NDR1, a locus of Arabidopsisthaliana that is required for disease resistance to both a bacterial and a fungalpathogen. Proceedings of the National Academy of Sciences, USA 92: 6597–6601.
Chen H, Zou Y, Shang Y, Lin H, Wang Y, Cai R, Tang X, Zhou JM. 2008.
Firefly luciferase complementation imaging assay for protein–proteininteractions in plants. Plant Physiology 146: 368–376.
Cheng YT, Germain H, Wiermer M, Bi D, Xu F, Garcia AV, Wirthmueller L,
Despres C, Parker JE, Zhang Y et al. 2009. Nuclear pore complex component
MOS7/Nup88 is required for innate immunity and nuclear accumulation of
defense regulators in Arabidopsis. Plant Cell 21: 2503–2516.Clough SJ, Bent AF. 1998. Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. PlantJournal 16: 735–743.
Feys BJ, Moisan LJ, Newman MA, Parker JE. 2001. Direct interaction between
the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4. EMBOJournal 20: 5400–5411.
Goritschnig S, Weihmann T, Zhang Y, Fobert P, McCourt P, Li X. 2008. A
novel role for protein farnesylation in plant innate immunity. Plant Physiology148: 348–357.
Goritschnig S, Zhang Y, Li X. 2007. The ubiquitin pathway is required for
innate immunity in Arabidopsis. Plant Journal 49: 540–551.Guy CL. 1990. Cold acclimation and freezing stress tolerance: role of protein
metabolism. Annual Review of Plant Physiology and Plant Molecular Biology 41:187–223.
Heidrich K, Wirthmueller L, Tasset C, Pouzet C, Deslandes L, Parker JE. 2011.
Arabidopsis EDS1 connects pathogen effector recognition to cell
compartment-specific immune responses. Science 334: 1401–1404.Hua J. 2013.Modulation of plant immunity by light, circadian rhythm, and
temperature. Current Opinion in Plant Biology 16: 406–413.Hua J, Grisafi P, Cheng SH, Fink GR. 2001. Plant growth homeostasis is
controlled by the Arabidopsis BON1 and BAP1 genes. Genes & Development15: 2263–2272.
Huang X, Li J, Bao F, Zhang X, Yang S. 2010. A gain-of-function mutation in
the Arabidopsis disease resistance gene RPP4 confers sensitivity to lowtemperature. Plant Physiology 154: 796–809.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 13

Hubert DA, Tornero P, Belkhadir Y, Krishna P, Takahashi A, Shirasu K,
Dangl JL. 2003. Cytosolic HSP90 associates with and modulates the
Arabidopsis RPM1 disease resistance protein. EMBO Journal 22: 5679–5689.Ichimura K, Casais C, Peck SC, Shinozaki K, Shirasu K. 2006.MEKK1 is
required for MPK4 activation and regulates tissue-specific and
temperature-dependent cell death in Arabidopsis. Journal of BiologicalChemistry 281: 36969–36976.
Jones JD, Dangl JL. 2006. The plant immune system. Nature 444: 323–329.Kim HS, Delaney TP. 2002. Arabidopsis SON1 is an F-box protein that
regulates a novel induced defense response independent of both salicylic acid
and systemic acquired resistance. Plant Cell 14: 1469–1482.Knoth C, Ringler J, Dangl JL, Eulgem T. 2007. Arabidopsis WRKY70 is
required for full RPP4-mediated disease resistance and basal defense against
Hyaloperonospora parasitica.Molecular Plant–Microbe Interactions 20: 120–128.Krishna P, Gloor G. 2001. The Hsp90 family of proteins in Arabidopsis thaliana.Cell Stress & Chaperones 6: 238–246.
Lee H, Guo Y, Ohta M, Xiong L, Stevenson B, Zhu JK. 2002. LOS2, a geneticlocus required for cold-responsive gene transcription encodes a bi-functional
enolase. EMBO Journal 21: 2692–2702.Li X, Clarke JD, Zhang Y, Dong X. 2001. Activation of an EDS1-mediated
R-gene pathway in the snc1mutant leads to constitutive, NPR1-independent
pathogen resistance.Molecular Plant–Microbe Interactions 14: 1131–1139.Li Y, Tessaro MJ, Li X, Zhang Y. 2010. Regulation of the expression of plant
resistance gene SNC1 by a protein with a conserved BAT2 domain. PlantPhysiology 153: 1425–1434.
Liu Y, Burch-Smith T, Schiff M, Feng S, Dinesh-Kumar SP. 2004.Molecular
chaperone Hsp90 associates with resistance protein N and its signaling proteins
SGT1 and Rar1 to modulate an innate immune response in plants. Journal ofBiological Chemistry 279: 2101–2108.
Maekawa T, Cheng W, Spiridon LN, Toller A, Lukasik E, Saijo Y, Liu P, Shen
QH, Micluta MA, Somssich IE et al. 2011. Coiled-coil domain-dependent
homodimerization of intracellular barley immune receptors defines a minimal
functional module for triggering cell death. Cell Host & Microbe 9: 187–199.Malamy J, Hennig J, Klessig DF. 1992. Temperature-dependent induction of
salicylic acid and its conjugates during the resistance response to tobacco
mosaic virus infection. Plant Cell 4: 359–366.McDowell JM, Cuzick A, Can C, Beynon J, Dangl JL, Holub EB. 2000. Downy
mildew (Peronospora parasitica) resistance genes in Arabidopsis vary in
functional requirements for NDR1, EDS1, NPR1 and salicylic acid
accumulation. Plant Journal 22: 523–529.Moffett P, Farnham G, Peart J, Baulcombe DC. 2002. Interaction between
domains of a plant NBS-LRR protein in disease resistance-related cell death.
EMBO Journal 21: 4511–4519.Monaghan J, Xu F, Gao M, Zhao Q, Palma K, Long C, Chen S, Zhang Y, Li X.
2009. Two Prp19-like U-box proteins in the MOS4-associated complex play
redundant roles in plant innate immunity. PLoS Pathogens 5: e1000526.Ni M, Cui D, Einstein J, Narasimhulu S, Vergara CE, Gelvin SB. 1995.
Strength and tissue specificity of chimeric promoters derived from the octopine
and mannopine synthase genes. Plant Journal 7: 661–676.Palma K, Zhang Y, Li X. 2005. An importin a homolog, MOS6, plays an
important role in plant innate immunity. Current Biology 15: 1129–1135.Palma K, Zhao Q, Cheng YT, Bi D, Monaghan J, Cheng W, Zhang Y, Li X. 2007.
Regulation of plant innate immunity by three proteins in a complex conserved
across the plant and animal kingdoms. Genes & Development 21: 1484–1493.Parker JE, Holub EB, Frost LN, Falk A, Gunn ND, Daniels MJ. 1996.
Characterization of eds1, a mutation in Arabidopsis suppressing resistance to
Peronospora parasitica specified by several different RPP genes. Plant Cell 8:2033–2046.
Prasinos C, Krampis K, Samakovli D, Hatzopoulos P. 2005. Tight regulation of
expression of two Arabidopsis cytosolic Hsp90 genes during embryo
development. Journal of Experimental Botany 56: 633–644.Rairdan GJ, Moffett P. 2006. Distinct domains in the ARC region of the potato
resistance protein Rx mediate LRR binding and inhibition of activation. PlantCell 18: 2082–2093.
Samakovli D, Thanou A, Valmas C, Hatzopoulos P. 2007. Hsp90
canalizes developmental perturbation. Journal of Experimental Botany 58:
3513–3524.
Seo PJ, Kim MJ, Park JY, Kim SY, Jeon J, Lee YH, Kim J, Park CM. 2010.
Cold activation of a plasma membrane-tethered NAC transcription factor
induces a pathogen resistance response in Arabidopsis. Plant Journal 61:661–671.
Shen QH, Saijo Y, Mauch S, Biskup C, Bieri S, Keller B, Seki H, Ulker B,
Somssich IE, Schulze-Lefert P. 2007. Nuclear activity of MLA immune
receptors links isolate-specific and basal disease-resistance responses. Science315: 1098–1103.
Swiderski MR, Birker D, Jones JD. 2009. The TIR domain of TIR-NB-LRR
resistance proteins is a signaling domain involved in cell death induction.
Molecular Plant–Microbe Interactions 22: 157–165.Takahashi A, Casais C, Ichimura K, Shirasu K. 2003. HSP90 interacts with
RAR1 and SGT1 and is essential for RPS2-mediated disease resistance in
Arabidopsis. Proceedings of the National Academy of Sciences, USA 100:
11777–11782.Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB. 1997. Subcellular
localization of H2O2 in plants. H2O2 accumulation in papillae and
hypersensitive response during the barley–powdery mildew interaction. PlantJournal 11: 1187–1194.
Tsutsui T, Kato W, Asada Y, Sako K, Sato T, Sonoda Y, Kidokoro S,
Yamaguchi-Shinozaki K, Tamaoki M, Arakawa K et al. 2009. DEAR1, a
transcriptional repressor of DREB protein that mediates plant defense and
freezing stress responses in Arabidopsis. Journal of Plant Research 122:633–643.
Upchurch RG, Ramirez ME. 2011. Effects of temperature during soybean seed
development on defense-related gene expression and fungal pathogen
accumulation. Biotechnology Letters 33: 2397–2404.Wagner S, Rietz S, Parker JE, Niefind K. 2011. Crystallization and preliminary
crystallographic analysis of Arabidopsis thaliana EDS1, a key component of
plant immunity, in complex with its signalling partner SAG101. ActaCrystallographica Section F, Structural biology and CrystallizationCommunications 67: 245–248.
Wang Y, Zhang Y, Wang Z, Zhang X, Yang S. 2013. A missense mutation in
CHS1, a TIR-NB protein, induces chilling sensitivity in Arabidopsis. PlantJournal 75: 553–565.
Wirthmueller L, Zhang Y, Jones JD, Parker JE. 2007. Nuclear accumulation of
the Arabidopsis immune receptor RPS4 is necessary for triggering
EDS1-dependent defense. Current Biology 17: 2023–2029.Xu S, Zhang Z, Jing B, Gannon P, Ding J, Xu F, Li X, Zhang Y. 2011.
Transportin-SR is required for proper splicing of resistance genes and plant
immunity. PLoS Genetics 7: e1002159.Yang H, Shi Y, Liu J, Guo L, Zhang X, Yang S. 2010. A mutant CHS3 protein
with TIR-NB-LRR-LIM domains modulates growth, cell death and freezing
tolerance in a temperature-dependent manner in Arabidopsis. Plant Journal 63:283–296.
Yang S, Hua J. 2004. A haplotype-specific resistance gene regulated by
BONZAI1 mediates temperature-dependent growth control in Arabidopsis.
Plant Cell 16: 1060–1071.Yi H, Richards EJ. 2007. A cluster of disease resistance genes in Arabidopsis is
coordinately regulated by transcriptional activation and RNA silencing. PlantCell 19: 2929–2939.
Zhai Z, Sooksa-nguan T, Vatamaniuk OK. 2009. Establishing RNA interference
as a reverse-genetic approach for gene functional analysis in protoplasts. PlantPhysiology 149: 642–652.
Zhang Y, Cheng YT, Bi D, Palma K, Li X. 2005.MOS2, a protein containing
G-patch and KOWmotifs, is essential for innate immunity in Arabidopsisthaliana. Current Biology 15: 1936–1942.
Zhang Y, Dorey S, Swiderski M, Jones JD. 2004. Expression of RPS4 in tobacco
induces an AvrRps4-independent HR that requires EDS1, SGT1 and HSP90.
Plant Journal 40: 213–224.Zhang Y, Goritschnig S, Dong X, Li X. 2003. A gain-of-function mutation in a
plant disease resistance gene leads to constitutive activation of downstream
signal transduction pathways in suppressor of npr1-1, constitutive 1. Plant Cell15: 2636–2646.
Zhang Y, Li X. 2005. A putative nucleoporin 96 is required for both basal
defense and constitutive resistance responses mediated by suppressor of npr1-1,constitutive 1. Plant Cell 17: 1306–1316.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist14

Zhu S, Jeong RD, Venugopal SC, Lapchyk L, Navarre D, Kachroo A, Kachroo
P. 2011. SAG101 forms a ternary complex with EDS1 and PAD4 and is
required for resistance signaling against turnip crinkle virus. PLoS Pathogens 7:e1002318.
Zhu Y, Qian W, Hua J. 2010. Temperature modulates plant defense responses
through NB-LRR proteins. PLoS Pathogens 6: e1000844.Zhu Z, Xu F, Zhang Y, Cheng YT, Wiermer M, Li X. 2010. Arabidopsis
resistance protein SNC1 activates immune responses through association with a
transcriptional corepressor. Proceedings of the National Academy of Sciences, USA107: 13960–13965.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Expression of HSP90 and different domains of RPP4protein in Nicotiana benthamiana leaves.
Fig. S2 The effect of temperature on rpp4 protein levels inArabidopsis protoplast.
Fig. S3 The effect of hsp90 on rpp4 protein levels in Arabidopsisprotoplast.
Fig. S4 Characterization of the intragenic suppressors of rpp4-1dArabidopsis mutant.
Fig. S5 The mutated positions of the intragenic suppressors ofrpp4-1d in RPP4 protein of Arabidopsis.
Fig. S6 RPP4 expression in rpp4-r25, rpp4-r26, rpp4-r27 andTIR-overexpressing Arabidopsis plants revealed by semi-quantita-tive reverse transcription-polymerase chain reaction (RT-PCR).
Fig. S7 Phenotypes of the rpp4-1d mos and rpp4-1d wrky70 dou-ble Arabidopsis mutants at 16°C.
Table S1 Gene-specific primers used in this study
Table S2 Comparison of morphological phenotypes of hsp90Arabidopsis mutants
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist is an electronic (online-only) journal owned by the New Phytologist Trust, a not-for-profit organization dedicatedto the promotion of plant science, facilitating projects from symposia to free access for our Tansley reviews.
Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged. We are committed to rapid processing, from online submission through to publication ‘as ready’ via Early View – our average timeto decision is <25 days. There are no page or colour charges and a PDF version will be provided for each article.
The journal is available online at Wiley Online Library. Visit www.newphytologist.com to search the articles and register for tableof contents email alerts.
If you have any questions, do get in touch with Central Office ([email protected]) or, if it is more convenient,our USA Office ([email protected])
For submission instructions, subscription and all the latest information visit www.newphytologist.com
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 15