antarctic glaciation recorded in early miocene new zealand foraminifera
TRANSCRIPT

Marine Micropaleontology 92–93 (2012) 52–60
Contents lists available at SciVerse ScienceDirect
Marine Micropaleontology
j ourna l homepage: www.e lsev ie r .com/ locate /marmic ro
Antarctic glaciation recorded in Early Miocene New Zealand foraminifera
Kenichi Fukuda a,1, Daniel B. Thomas b,2, Russell D. Frew a, R. Ewan Fordyce b,⁎a Department of Chemistry, University of Otago, P.O. Box 56, Dunedin 9054, New Zealandb Department of Geology, University of Otago, P.O. Box 56, Dunedin 9054, New Zealand
⁎ Corresponding author. Tel.: +64 3 479 7510; fax: +E-mail addresses: [email protected] (K. Fuk
(D.B. Thomas), [email protected] (R.D. Frew), ew(R.E. Fordyce).
1 Present address: Nakashima Propeller Co. Ltd., DeveOkayama, Japan.
2 Present address: Department of Vertebrate Zoology,Natural History, Smithsonian Institution, P.O. Box 37012
0377-8398/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.marmicro.2012.05.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 December 2010Received in revised form 9 May 2012Accepted 12 May 2012Available online 29 May 2012
Keywords:foraminiferageochemistryMg/Ca thermometerpalaeoclimatepalaeotemperature
New Zealand sedimentary sequences are important repositories of southern temperate palaeoenvironmentaldata, as may be interpreted from biogenic chemical signals preserved in marine microfossils. Calibration cur-ves for Mg/Ca ratios versus water temperatures were established using modern benthic foraminifera,Notorotalia and Cibicides. Notorotalia is a long-ranged endemic benthic genus with a good record in shelf sed-iments, while Cibicides allows comparisons with similar studies elsewhere. The resulting correlations were T(°C)=ln(Mg/Ca [mmol/mol] /1.64)×10.89 for Cibicides spp., and T (°C)=ln(Mg/Ca [mmol/mol] /0.44)×5.71for Notorotalia spp. Well-preserved Early Miocene Notorotalia and Cibicides were collected for paired Mg/Caand δ18O analysis from a 3.6 m section of the Mount Harris Formation spanning an estimated 60 ka and dat-ing from about 17.7 Ma (Globoconella zealandica zone, roughly middle Burdigalian, Early Miocene) within thelocal Altonian Stage (15.9–18.7 Ma). Mg/Ca bottom-water palaeotemperature estimates from Cibicides andNotorotalia gave concordant results: 13.3±1.0 °C for Notorotalia spinosa, 15.5±3.0 °C for Cibicides spp. Esti-mates of oxygen isotopic composition for Altonian sea water (δ18Opalaeo-sw) were −0.4±0.4‰, suggestingthe presence of small ice sheets on Antarctica. The method used to generate such results has far reaching im-plications for reconstructing δ18Opalaeo-sw, and should allow Antarctic ice volume history to be finely resolvedfrom New Zealand sequences.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
Oxygen isotope measurements from calcareous marine inverte-brates (δ18Ocarbonate, hereafter δ18Oc) have been widely employed as aproxy for palaeotemperature (e.g. Miller et al., 1987; Billups andSchrag, 2002; Hollis et al., 2009). The δ18Oc depends on the water tem-perature during carbonate formation and also on the δ18O of the waterin which the carbonate was formed. The δ18Oc composition of marineinvertebrates is dependent on salinity, and hence global ice volume,and currently binds reliable palaeotemperature estimates to indepen-dent estimates of polar ice volume (e.g. Shackleton, 1967). The dual sig-nals of salinity and ambient temperature contained as δ18Oc withinbiogenic carbonates have historically proven inseparable without inde-pendent proxies (e.g. alkenone saturation, faunal abundance; Elderfieldand Ganssen, 2000). One recently-favoured approach for establishingthe ambient temperature of growth in foraminifera is the assessment
64 3 479 7527.uda), [email protected]@otago.ac.nz
lopment Group, R&D Division,
MRC-116, National Museum of, Washington, D.C. 20013, USA.
rights reserved.
of theMg/Ca ratiowithin test carbonate: incorporation ofMg into the bio-genic calcite lattice is influenced by ambient temperature during calcifica-tion (Nürnberg et al., 1996; Rosenthal et al., 2000). The relationshipbetween Mg concentration of a foraminiferal test and temperature isnot universal across all species (Toyofuku et al., 2000; Dissard et al.,2010), thus requiring interpretations of fossil data to be grounded bystudies of modern representatives of particular fossil lineages.
Manyof the extant foraminifera frommodern-dayNewZealand havea significant fossil record (e.g. Hornibrook et al., 1989). The Cenozoic bio-stratigraphy of New Zealand is richly resolved (Hornibrook et al., 1989;Morgans et al., 1999; Graham et al., 2000; Cooper, 2004), with excellentdocumentation of foraminiferal localities. The New Zealand Cenozoicmarine record,which accumulated inmid-temperate southern latitudes,is readily accessible in onland outcrops of thin Palaeogene strata that ac-cumulated in a passive margin setting, andmuch thicker Neogene stratathat reflect development of an active plate boundary. Due to such attri-butes, New Zealand amongst southern landmasses is a Rosetta stonefor understanding Southern Hemisphere and Antarctic-related climatesignals (Zachos et al., 2008; Field et al., 2009) resulting from the break-up of Gondwana. As elsewhere, interpretation of the New Zealandpalaeoclimate record would be greatly assisted by the ability to teasetemperature from salinity signals stored in calcareous microfossils.Here we evaluate the efficacy of recovering temperature and salinitysignals from modern and related fossil foraminifera using a pairedMg/Ca and δ18Oc approach.

53K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
2. Materials and methods
2.1. Geologic setting
Modern foraminifera were collected from South Pacific and SouthernOcean sediments (localities detailed in Section 3.1; Fig. 1). Fossil forami-nifera were extracted from an outcrop of the Mount Harris Formation atPukeuri, North Otago, New Zealand (44°46′S, 171°01′E, or NZMS 260 ref-erence J41/544733; Fig. 1). TheMount Harris Formation is a soft, massive,light yellow-brown calcareous sandy siltstone to silty sandstone that ac-cumulated in a quiet setting below the stormwave base, probably on theouter shelf (discussed below). All samples collected for the present studyincluded the planktonic foraminiferaGloboconella zealandica, a zonal spe-cies with a first appearance datum in Chron C5En (18.06–18.52 Ma;Morgans et al., 2002; Gradstein et al., 2004) that marks the middle ofthe New Zealand Altonian Stage (Early Miocene, 15.9–18.7 Ma: Hollis etal., 2010). The Altonian Stage spans the later Burdigalian and earlierLanghian Stages (Cooper, 2004). Further, two Lentipecten sp. (scallop —
Pectinidae) shell samples within the Pukeuri sequence had 87Sr/86Srcompositions of 0.708625±0.000011 (shell 10 cm immediately belowforaminiferal sample 4) and 0.708633±0.000010 (shell level with fora-miniferal sample 7, about 50 cm along bedding plane) (D.A. Teagle andM. Cooper, pers. comm.), corresponding to 17.74 Ma and 17.65 Ma, re-spectively, and each rounding to 17.7 Ma (McArthur et al., 2001).
2.2. Samples
Modern benthic foraminifera specimens representing tropical to sub-polar latitudes were obtained from sediments within Fijian and NewZealand waters (Fig. 1). Specimens of Cibicides were collected fromKadavu Passage (18°11′S, 178°25′E, 135 m below sea level, BSL), ThreeKings Plateau (34°20′S, 172°30′E, 100 m BSL), Challenger Plateau(42°14′S, 169°30′E, 1125 m BSL), Taieri Bight, Otago (46°10′S, 170°23′E,66 m BSL), Enderby Island (50°50′S, 166°30′E, 115 m BSL) and CampbellIsland (52°32′S, 169°10′E, 15 mBSL). Samples ofNotorotaliawere collect-ed from Three Kings Plateau (34°20′S, 172°30′E, 100 m BSL), two siteswithin Blueskin Bay, Otago (45°44′S, 170°41′E, 20 m BSL and 45°46′S,171°05′E, 207 m BSL), Enderby Island (50°30′S, 166°19′E, 2–3 m BSL)and Auckland Island (50°32′S, 166°12′E, 2–3 m BSL). Kadavu Passagesamples were collected during the 2nd Joint National Fisheries Universi-ty–University of the South Pacific Fisheries and Oceanography Research
Fig. 1. Locality map for specimens collected and analysed in this study. A) Modern samplesShelf (Ot) and Auckland (Au), Enderby (En) and Campbell (Ca) Islands. Fossil samples werenorth of Oamaru at 45.0°S, and was located at 43.3°S during the middle of the local Altonia
Cruise (3–7 December 1997) by the Koyo-Maru (commercial vessel)using a Van Veen grab sampler. Otago Shelf samples were collectedas grab samples by the R/VMunida (19 September 2001) (Universityof Otago research vessel) and a 6 m alloy runabout (05 November2003). Samples from Three Kings and Challenger Plateaus were col-lected by the R/V Tangaroa (20 June 1978) (NIWA National Instituteof Water and Atmospheric Research Ltd. research vessel) andWestbay (6–10 November 1995) (commercial deep-sea trawler) usingan anchor dredge. Shallow water samples from Auckland, Campbell andEnderby Islands were provided by J. Guise (University of Canterbury),and were supplemented by samples collected in the early 1900s andhoused at Canterbury Museum, New Zealand (Chapman, 1909). Sea sur-face temperatures (SST) or bottom water temperatures (BWT) for theKadavu Passage, Challenger Plateau, Otago Shelf and Auckland Islandsites were collated from earlier studies (Heath, 1984; Pickering andSuda, 2003; Van Hale, 2003). Temperature for the Three Kings Plateausample site was estimated from oxygen isotopes derived from carbonate(unpublished thesis: Fukuda, 2002), with data for the Campbell andEnderby Island sites taken from the National Institute of Water and At-mosphere (NIWA) archive.
Fossil foraminifera were collected from a 3.6 m thick sequence of theMount Harris Formation, with 2 cm slices taken at 33 cm intervals (12 fo-raminiferal sample horizons and 1 Sr sample horizon, numbered as J40/f278 to J40/f 290 in the Geological Society of New Zealand Fossil RecordFile). The Mount Harris Formation at the Pukeuri locality yields well-preserved foraminifera, commonlywith translucent (glassy) tests that re-tain micron-level detail under scanning electron microscopy, with prom-ise of limited diagenetic alteration. Based on sedimentation ratesdiscussed for the Mount Harris Formation (62.5 mMa−1; e.g. Morganset al., 1999, using the nameBluecliffs Silt), the sampled interval at Pukeuriis expected to span approximately 60 ka. The present studyprovides a de-tailed description of a thin sequence spanning a 60 ka interval about17.7 Ma ago.
2.3. Modern sample extraction
Decanted oceanic sediments containing modern foraminifera weresaturated with Rose Bengal stain (BDH Chemicals) solution (2 g L−1 eth-anol) for 12 h to distinguish between recent living and relict foraminifera(Walton, 1952). Samples were thoroughly dried at 40 °C, allowing pink(protoplasm-stained) specimens of Cibicides corticatus, Cibicides dispars,
were collected from Fiji (Fi), Three Kings Plateau (Th), Challenger Plateau (Ch), Otagocollected from Pukeuri (Pu), North Otago, New Zealand. B) Pukeuri is currently locatedn Stage (15.9–18.7 Ma) (King, 2000).

54 K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
Cibicides marlboroughensis, Notorotalia depressa, Notorotalia finlayi,Notorotalia hornibrooki, Notorotalia inornata, Notorotalia olsoni andNotorotalia zealandica to be collected and stored for later analysis.
2.4. Fossil extraction
The Mount Harris Formation sediment samples (~100–300 g) weredried at 60 °C overnight, weighed, and soaked in a dispersion solutioncontaining ~10 g analytical grade sodium hexametaphosphate (NaPO3)6(BDH Chemicals) dissolved in ~500ml distilled water. Sediment and dis-persion solutionwere transferred to a stainless steel pot and dilutedwithexcess distilled water before being boiled for 30 min. Disaggregated sam-pleswere thenwashed (distilledwater) through a 63 μmstainless steel orbrass sieve. Collected sediment (>63 μmfraction)was then dried at 60 °Covernight and subsequently weighed, allowing the mass of the elutedmud fraction to bedetermined.Well preserved (not-infilled,minimal sur-face alteration) Notorotalia spinosa and Cibicides spp. were picked fromthe >63 μm fraction and stored for later analysis.
2.5. Siliceous sand and carbonate weight percent
Subsamples of the Mount Harris Formation sediment (~100–300 g)were dried,weighed and boiled prior to sieving to remove themud frac-tion (following the fossil extraction method above). Samples were thenoven dried (60 °C) for approximately 12 h before again being weighed,and were then soaked in excess 0.075 M HNO3 to dissolve carbonate.The residue was filtered, washed, dried and reweighed. Percentages ofboth siliceous sand and calcareous material were calculated relative tothe initial dry weight of each subsample.
2.6. Planktonic fraction
Between 50 and 100 g of the prepared >63 μm fraction of eachMount Harris Formation sediment sample was resieved using a150 μm mesh (e.g. Morgans et al., 2002). The >150 μm fraction wasthen split repeatedly until a few hundred (desirably >100 and b300)complete foraminifera remained. All foraminifera were then pickedfrom each sample using a Zeiss DR or SV11 binocular microscope, andwere transferred to slides for counting and identification.
2.7. Mg/Ca measurement
Prior to analysis, modern and fossil specimens were cleaned as fol-lows. Individual specimens were crushed between cleaned glass plates,with ~30 μg of crushed sample transferred into a 0.5 ml plastic vial withdoubly distilled water (Milli-Q®). Each sample was sonicated withinthe plastic vial for 1 min; supernatant water was then removed by vac-uumpipette and replacedwithMilli-Q®, before a secondminute of son-ication. Samples were rinsed with Milli-Q® and sonicated three times,followed by two sequences of rinse and sonication with methanol,and a final rinse and sonication with Milli-Q®. Approximately 2 mlalkaline peroxide solution (0.1% H2O2 in 0.1 M NaOH) were added toeach sample, which were then boiled for 10 min, and sonicated for30 s. Alkaline peroxide solution was replaced with Milli-Q® for a setof four rinses. Each sample was then subjected to a dilute acid leach(0.25 ml 0.001 M HNO3; 30 s with sonication) before two final rinseswith Milli-Q®. The level of contamination and cleaning efficacy ofeach step was evaluated through concurrent analyses of a marble labo-ratory standard. To track the fidelity of foraminiferal carbonate, repre-sentative N. spinosa specimens were imaged before and after the acidcleaned glass plate step using a Cambridge 360 scanning electronmicroscope (SEM) (Department of Anatomy, University of Otago).
Cleaned samples were dissolved in 0.4 ml 0.075 M HNO3. Mg/Caratios were determined using inductively coupled plasma opticalemission spectrometry (ICP-OES, Thermo Jarrell Ash) (Departmentof Chemistry, University of Otago). Single specimens of Notorotalia
generally sufficed for analysis, but 2 specimens of Cibicides spp.were required to quantify Mg. Mg/Ca ratios were calculated usingonly data with a precision (relative standard deviation) better than5%.
2.8. Stable isotope measurements
Oxygen isotopic composition of test carbonate (δ18Oc)was establishedformodern Cibicides sp. andNotorotalia spp., aswell asN. spinosa from theMount Harris Formation. Samples were cleaned prior to analysis: speci-mens were transferred into plastic vials with Milli-Q® and sonicated for1 min, followed by sonication inmethanol. Sampleswere further sonicat-ed in 12.5% NaClO to remove organic material and rinsed by being soni-cated in Milli-Q®. Samples were then oven-dried (40 °C) before beingcrushed. Between 700 μg and 1 mg of crushed foraminiferal powderwas reacted with 104% H3PO4 at 25 °C, with the evolved CO2 gascryogenically-purified and analysed using a Europa Scientific 20–20 iso-tope ratio mass spectrometer in dual-inlet mode. Oxygen and carbon iso-tope values were reported relative to Vienna Peedee Belemnite (VPDB)standardviaNational Bureau of Standards (NBS)19 anda laboratory stan-dard (δ18O=−2.45‰). A correction to the oxygen isotopic compositionof seawater (δ18Osw;−0.27‰) was used for converting Vienna StandardMean Ocean Water (VSMOW) to VPDB (Hut, 1987). Palaeoseawater iso-topic compositions (δ18Opalaeo-sw) are here reported relative to VSMOW.
3. Results
3.1. Mg/Ca measurements
Mg/Ca ratios were measured in modern Notorotalia spp. from foursample localities (Table 1): four specimens fromEnderby Island (2.1±0.4 mmol/mol), five from Auckland Island (2.3±0.6 mmol/mol), 16from Otago Shelf (2.7±0.3 mmol/mol) and 10 from Three Kings Pla-teau (6.7±0.9 mmol/mol) (mean±95% confidence interval; onlineSupplementary material). Mg/Ca ratios were also measured in mod-ern Cibicides spp. from six localities (Table 1): five from Challenger Pla-teau (2.1±0.2 mmol/mol), five from Enderby Island (2.6±0.28 mmol/mol), four from Campbell Island (3.9±1.3 mmol/mol), six from OtagoShelf (5.3±0.4 mmol/mol), six from Three Kings Plateau (6.3±1.3 mmol/mol) and five from Kadavu Passage (12.9±1.3 mmol/mol)(mean±95% confidence interval; online Supplementary material).
The Mg/Ca ratios of 53 N. spinosa and 23 Cibicides sp. specimensfrom the Mount Harris Formation at Pukeuri, North Otago were alsomeasured (Fig. 2; online Supplementary material). N. spinosa gaverelatively consistent Mg/Ca ratios across all 12 sampling horizons,ranging between 2.5±0.2 mmol/mol (n=4, horizon 10) and 3.8±0.2 mmol/mol (n=5, horizon 8), with an average for all sample horizonsof 3.3±0.2 mmol/mol. Mg/Ca ratios from Cibicides spp. demonstratedconsiderable variation both within and between sample horizons, withan average of 5.0±1.1 mmol/mol from a range of 3.3±1.0 mmol/mol(n=2, horizon 2) to 8.4 (n=1, horizon 1) mmol/mol.
Samples were cleaned prior to Mg/Ca analysis, and cleaning efficacywas checked using scanning electronmicroscope (SEM) images collectedfrom fossil Notorotalia spp. before and after the weak acid leach. Imagesshowed that most secondary carbonates were removed. The Mg/Caratio of a marble laboratory standard did not change with subsequentcleaning steps. Further, Rose Bengal stain did not alter the Mg/Ca ratioof foraminifera (unstained Notorotalia sp.=2.7 mmol/mol σ=0.3,n=4; stained Notorotalia sp.=2.63mmol/mol, n=1). Elderfield et al.(2006) also noted that Rose Bengal stain does not affect the Mg/Ca ratio.
3.2. Stable isotope measurements
Oxygen isotopic ratios (δ18Oc) for modern Cibicides sp. andNotorotalia spp., as well as fossil N. spinosa, were determined in

Table 1Sample and locality data for modern Cibicides and Notorotalia.
Locality Latitude (°S) Longitude (°E) Species N Depth (m) Annual mean water temperature (°C)
CibicidesChallenger Plateau 42°14′ 169°30′ Cibicides sp. 1 1125 4.7Challenger Plateau 42°14′ 169°30′ C. corticans 1 1125 4.7Challenger Plateau 42°14′ 169°30′ C. dispars 2 1125 4.7Challenger Plateau 42°14′ 169°30′ C. marlboroughensis 1 1125 4.716 km north of Enderby Is. 50°50′ 166°30′ C. dispars 4 115 5.5Campbell Island 52°32′ 169°10′ C. dispars 4 15 7.5Taieri Bight, Otago 46°10′ 170°23′ C. corticans 7 66 10.5Three Kings Plateau 34°20′ 172°30′ C. marlboroughensis 6 100 15.0Kadavu Passage, Fiji 18°11′ 178°25′ C. dispars 5 135 23.0
NotorotaliaEnderby Island 50°30′ 166°19′ Notorotalia sp. 4 2–3 8.5Auckland Island 50°32′ 166°12′ Notorotalia sp. 5 2–3 8.5Otago Shelf 45°44′ 170°41′ Notorotalia sp. 10 20 11.6Otago Shelf 45°46′ 171°05′ N. zealandica 6 207 10.1Three Kings Plateau 34°20′ 172°30′ N. hornibrooki 2 100 15.0Three Kings Plateau 34°20′ 172°30′ N. inornata 1 100 15.0Three Kings Plateau 34°20′ 172°30′ N. olsoni 7 100 15.0
55K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
combination with Mg/Ca analyses. Four Cibicides specimens from theThree Kings Plateau sampling site exhibited a mean δ18Oc value of0.6±0.3‰ (mean±95% confidence interval;‰VSMOW). Four spec-imens of Notorotalia sp. from the Blueskin Bay locality along theOtago Shelf had δ18Oc values of 1.5±0.2‰, and two Notorotalia sp.specimens from Auckland Island exhibited values of 1.38±0.03‰.Early-stage diagenesis was assessed by calculating the oxygen isoto-pic composition of sea water (δ18Osw) from ambient growth temper-ature and oxygen isotopic composition of test carbonate (δ18Oc), andcomparing the calculated and expected VSMOW values (e.g. Horibeand Oba, 1972). All δ18Osw values were within the expected rangefor VSMOW.
The oxygen isotopic compositions of N. spinosa from Mt. HarrisFormation ranged from 0.2±0.1‰ (n=2, horizon 4) to 0.8‰(n=1, horizon 12), and averaged 0.5±0.2‰ (n=23, mean±95%
Fig. 2. Mg/Ca ratios for Cibicides and Notorotalia at 12 sample horizons from Pukeuri,North Otago, New Zealand (60 ka span around 17.7 Ma). Lentipecten sp. (scallop —
Pectinidae) shell was used to establish the two absolute dates (Sr isotopemeasurements).
confidence interval; ‰VSMOW) (Fig. 3; online Supplementary mate-rial). Analytical precisions (1 sd) determined by replicate analyses ofa marble laboratory standard were±0.04‰.
3.3. Environmental indices
Weight of sand and carbonate fractions within the Mount HarrisFormation were established as percentages for the 12 sample hori-zons at Pukeuri, North Otago (Fig. 4). Sand weight percent (wt.%)ranged from 30 to 57 wt.% and averaged 40±8 wt.% (Fig. 4). Theamount of carbonate within the sediment was relatively constant, av-eraging 1.6±0.9%. Sand fraction was predominantly angular quartz(very fine sand) with less abundant microfossils (foraminifera).
The percent of planktonic foraminifera in the >150 μm fraction ofeach sample averaged about 30% and ranged between 25.7% and 36.1%(Fig. 4). G. zealandica, which is diagnostic for the middle Altonian stage(Morgans et al., 2002), occurred as large but rare specimens in all 12
Fig. 3. Oxygen isotopic composition of Notorotalia spinosa collected from 12 sample ho-rizons at Pukeuri, North Otago, New Zealand (60 ka span around 17.7 Ma).
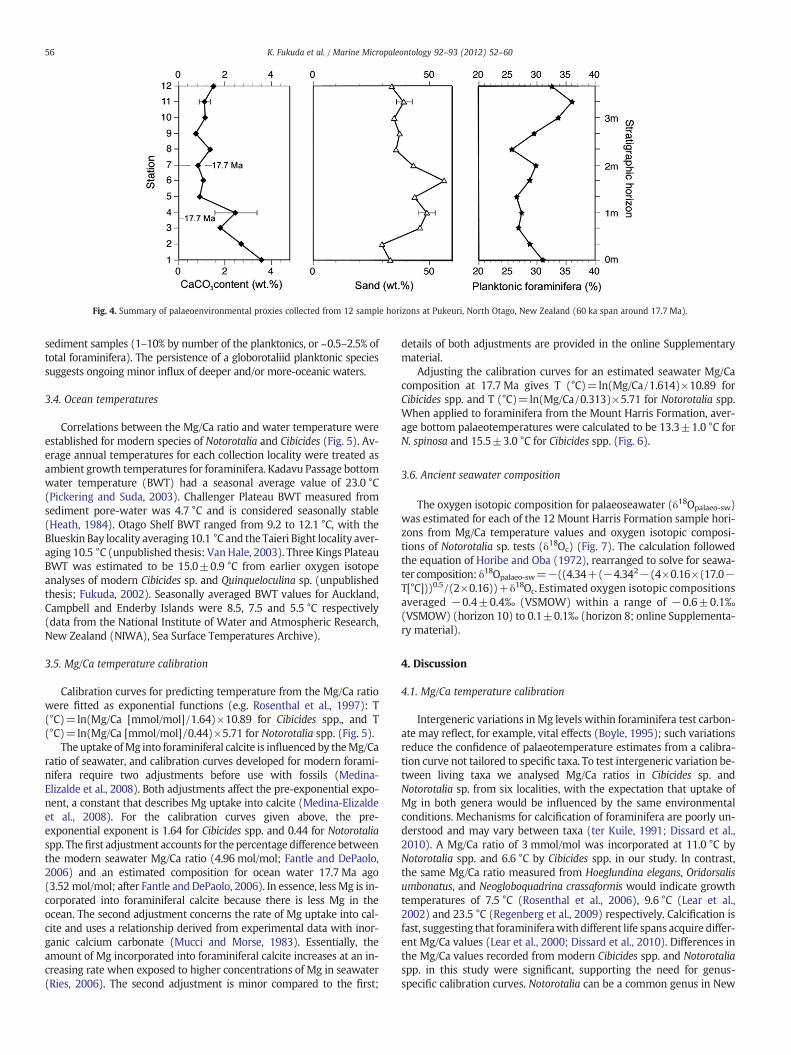
Fig. 4. Summary of palaeoenvironmental proxies collected from 12 sample horizons at Pukeuri, North Otago, New Zealand (60 ka span around 17.7 Ma).
56 K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
sediment samples (1–10% by number of the planktonics, or ~0.5–2.5% oftotal foraminifera). The persistence of a globorotaliid planktonic speciessuggests ongoing minor influx of deeper and/or more-oceanic waters.
3.4. Ocean temperatures
Correlations between the Mg/Ca ratio and water temperature wereestablished for modern species of Notorotalia and Cibicides (Fig. 5). Av-erage annual temperatures for each collection locality were treated asambient growth temperatures for foraminifera. Kadavu Passage bottomwater temperature (BWT) had a seasonal average value of 23.0 °C(Pickering and Suda, 2003). Challenger Plateau BWT measured fromsediment pore-water was 4.7 °C and is considered seasonally stable(Heath, 1984). Otago Shelf BWT ranged from 9.2 to 12.1 °C, with theBlueskin Bay locality averaging 10.1 °C and the Taieri Bight locality aver-aging 10.5 °C (unpublished thesis: VanHale, 2003). Three Kings PlateauBWT was estimated to be 15.0±0.9 °C from earlier oxygen isotopeanalyses of modern Cibicides sp. and Quinqueloculina sp. (unpublishedthesis; Fukuda, 2002). Seasonally averaged BWT values for Auckland,Campbell and Enderby Islands were 8.5, 7.5 and 5.5 °C respectively(data from the National Institute of Water and Atmospheric Research,New Zealand (NIWA), Sea Surface Temperatures Archive).
3.5. Mg/Ca temperature calibration
Calibration curves for predicting temperature from the Mg/Ca ratiowere fitted as exponential functions (e.g. Rosenthal et al., 1997): T(°C)=ln(Mg/Ca [mmol/mol]/1.64)×10.89 for Cibicides spp., and T(°C)=ln(Mg/Ca [mmol/mol]/0.44)×5.71 for Notorotalia spp. (Fig. 5).
The uptake ofMg into foraminiferal calcite is influenced by theMg/Caratio of seawater, and calibration curves developed for modern forami-nifera require two adjustments before use with fossils (Medina-Elizalde et al., 2008). Both adjustments affect the pre-exponential expo-nent, a constant that describes Mg uptake into calcite (Medina-Elizaldeet al., 2008). For the calibration curves given above, the pre-exponential exponent is 1.64 for Cibicides spp. and 0.44 for Notorotaliaspp. Thefirst adjustment accounts for the percentage differencebetweenthe modern seawater Mg/Ca ratio (4.96 mol/mol; Fantle and DePaolo,2006) and an estimated composition for ocean water 17.7 Ma ago(3.52 mol/mol; after Fantle and DePaolo, 2006). In essence, lessMg is in-corporated into foraminiferal calcite because there is less Mg in theocean. The second adjustment concerns the rate of Mg uptake into cal-cite and uses a relationship derived from experimental data with inor-ganic calcium carbonate (Mucci and Morse, 1983). Essentially, theamount of Mg incorporated into foraminiferal calcite increases at an in-creasing rate when exposed to higher concentrations of Mg in seawater(Ries, 2006). The second adjustment is minor compared to the first;
details of both adjustments are provided in the online Supplementarymaterial.
Adjusting the calibration curves for an estimated seawater Mg/Cacomposition at 17.7 Ma gives T (°C)=ln(Mg/Ca/1.614)×10.89 forCibicides spp. and T (°C)=ln(Mg/Ca/0.313)×5.71 for Notorotalia spp.When applied to foraminifera from the Mount Harris Formation, aver-age bottom palaeotemperatures were calculated to be 13.3±1.0 °C forN. spinosa and 15.5±3.0 °C for Cibicides spp. (Fig. 6).
3.6. Ancient seawater composition
The oxygen isotopic composition for palaeoseawater (δ18Opalaeo-sw)was estimated for each of the 12 Mount Harris Formation sample hori-zons from Mg/Ca temperature values and oxygen isotopic composi-tions of Notorotalia sp. tests (δ18Oc) (Fig. 7). The calculation followedthe equation of Horibe and Oba (1972), rearranged to solve for seawa-ter composition: δ18Opalaeo-sw=−((4.34+(−4.342−(4×0.16×(17.0−T[°C]))0.5/(2×0.16))+δ18Oc. Estimated oxygen isotopic compositionsaveraged −0.4±0.4‰ (VSMOW) within a range of −0.6±0.1‰(VSMOW) (horizon 10) to 0.1±0.1‰ (horizon 8; online Supplementa-ry material).
4. Discussion
4.1. Mg/Ca temperature calibration
Intergeneric variations in Mg levels within foraminifera test carbon-ate may reflect, for example, vital effects (Boyle, 1995); such variationsreduce the confidence of palaeotemperature estimates from a calibra-tion curve not tailored to specific taxa. To test intergeneric variation be-tween living taxa we analysed Mg/Ca ratios in Cibicides sp. andNotorotalia sp. from six localities, with the expectation that uptake ofMg in both genera would be influenced by the same environmentalconditions. Mechanisms for calcification of foraminifera are poorly un-derstood and may vary between taxa (ter Kuile, 1991; Dissard et al.,2010). A Mg/Ca ratio of 3 mmol/mol was incorporated at 11.0 °C byNotorotalia spp. and 6.6 °C by Cibicides spp. in our study. In contrast,the same Mg/Ca ratio measured from Hoeglundina elegans, Oridorsalisumbonatus, and Neogloboquadrina crassaformis would indicate growthtemperatures of 7.5 °C (Rosenthal et al., 2006), 9.6 °C (Lear et al.,2002) and 23.5 °C (Regenberg et al., 2009) respectively. Calcification isfast, suggesting that foraminiferawith different life spans acquire differ-ent Mg/Ca values (Lear et al., 2000; Dissard et al., 2010). Differences inthe Mg/Ca values recorded from modern Cibicides spp. and Notorotaliaspp. in this study were significant, supporting the need for genus-specific calibration curves. Notorotalia can be a common genus in New
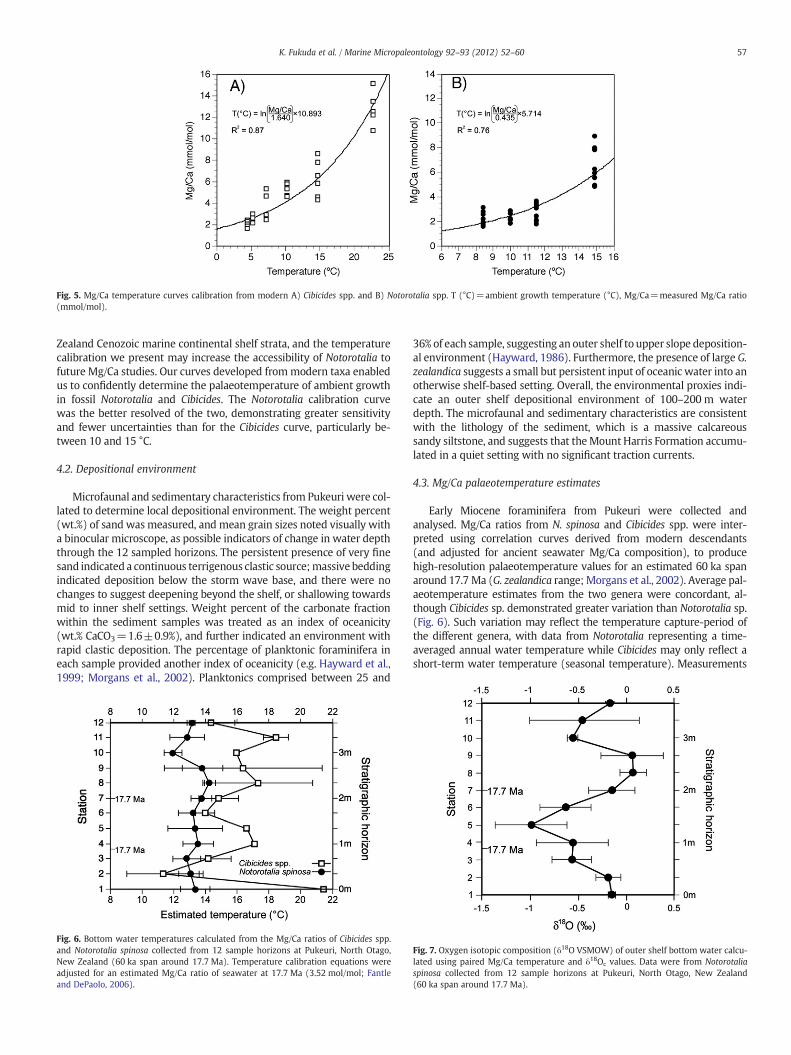
Fig. 5. Mg/Ca temperature curves calibration from modern A) Cibicides spp. and B) Notorotalia spp. T (°C)=ambient growth temperature (°C), Mg/Ca=measured Mg/Ca ratio(mmol/mol).
57K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
Zealand Cenozoic marine continental shelf strata, and the temperaturecalibration we present may increase the accessibility of Notorotalia tofuture Mg/Ca studies. Our curves developed frommodern taxa enabledus to confidently determine the palaeotemperature of ambient growthin fossil Notorotalia and Cibicides. The Notorotalia calibration curvewas the better resolved of the two, demonstrating greater sensitivityand fewer uncertainties than for the Cibicides curve, particularly be-tween 10 and 15 °C.
4.2. Depositional environment
Microfaunal and sedimentary characteristics from Pukeuri were col-lated to determine local depositional environment. The weight percent(wt.%) of sand was measured, and mean grain sizes noted visually witha binocular microscope, as possible indicators of change in water depththrough the 12 sampled horizons. The persistent presence of very finesand indicated a continuous terrigenous clastic source;massive beddingindicated deposition below the storm wave base, and there were nochanges to suggest deepening beyond the shelf, or shallowing towardsmid to inner shelf settings. Weight percent of the carbonate fractionwithin the sediment samples was treated as an index of oceanicity(wt.% CaCO3=1.6±0.9%), and further indicated an environment withrapid clastic deposition. The percentage of planktonic foraminifera ineach sample provided another index of oceanicity (e.g. Hayward et al.,1999; Morgans et al., 2002). Planktonics comprised between 25 and
Fig. 6. Bottom water temperatures calculated from the Mg/Ca ratios of Cibicides spp.and Notorotalia spinosa collected from 12 sample horizons at Pukeuri, North Otago,New Zealand (60 ka span around 17.7 Ma). Temperature calibration equations wereadjusted for an estimated Mg/Ca ratio of seawater at 17.7 Ma (3.52 mol/mol; Fantleand DePaolo, 2006).
36% of each sample, suggesting an outer shelf to upper slope deposition-al environment (Hayward, 1986). Furthermore, the presence of large G.zealandica suggests a small but persistent input of oceanic water into anotherwise shelf-based setting. Overall, the environmental proxies indi-cate an outer shelf depositional environment of 100–200 m waterdepth. The microfaunal and sedimentary characteristics are consistentwith the lithology of the sediment, which is a massive calcareoussandy siltstone, and suggests that theMount Harris Formation accumu-lated in a quiet setting with no significant traction currents.
4.3. Mg/Ca palaeotemperature estimates
Early Miocene foraminifera from Pukeuri were collected andanalysed. Mg/Ca ratios from N. spinosa and Cibicides spp. were inter-preted using correlation curves derived from modern descendants(and adjusted for ancient seawater Mg/Ca composition), to producehigh-resolution palaeotemperature values for an estimated 60 ka spanaround 17.7 Ma (G. zealandica range;Morgans et al., 2002). Average pal-aeotemperature estimates from the two genera were concordant, al-though Cibicides sp. demonstrated greater variation than Notorotalia sp.(Fig. 6). Such variation may reflect the temperature capture-period ofthe different genera, with data from Notorotalia representing a time-averaged annual water temperature while Cibicides may only reflect ashort-term water temperature (seasonal temperature). Measurements
Fig. 7. Oxygen isotopic composition (δ18O VSMOW) of outer shelf bottom water calcu-lated using paired Mg/Ca temperature and δ18Oc values. Data were from Notorotaliaspinosa collected from 12 sample horizons at Pukeuri, North Otago, New Zealand(60 ka span around 17.7 Ma).

58 K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
from single chambers of individual specimens of modern Notorotaliaspp. showed steady changes in Mg/Ca value, indicating calcite growthduring changing temperatures, and hence growth during multiple sea-sons (data not shown). In contrast, multiple Mg/Ca measurementsfrom individual, modern Cibicides spp. showed little variation (data notshown). The intergeneric differences observed here reinforce the needfor genus-specific Mg/Ca thermometric curves calibrated from moderndescendants.
Bottomwater palaeotemperature estimates for Pukeuri were approx-imately 14 °C at palaeolatitude 43.3°S (equivalent to modern day Christ-church, New Zealand, based on palaeogeographic reconstructions byKing, 2000). Our estimates are slightly lower than those previouslyreported for northern New Zealand during the Early Miocene: 17–22 °C(23.8 to 15.9 Ma, marine shelf; Devereux, 1967); 18–27 °C (~20 to~17 Ma, coral reef; Hayward, 1977); 15–18 °C (27.7 to 19.0 Ma, marineshelf; Nelson and Burns, 1982). Indeed, our Early Miocene temperatureis slightly warmer than a previous estimate for southern New Zealand(i.e. 10.4±0.3 °C, 21.7 to 19.0 Ma, marine shelf; Buening et al., 1998).Other New Zealand studies have generally identified the Altonian as atime of temperate to warm climate. Beu and Maxwell (1990) notedthat Altonian molluscs from the Mount Harris Formation resembled awarm-temperate fauna, and Hornibrook (1992) used foraminifera as-semblages to suggest that Altonian oceans around New Zealand werewarm to subtropical. Our results suggest that EarlyMioceneNewZealandouter shelf bottom temperatures were slightly warmer than at present.Further, our bottom water estimates for the southern Zealandia Shelfare within the range of contemporaneous sea surface temperatures esti-mated from Ocean Drilling Program (ODP) site 1171C (48°30′S, 149°07′E; Shevenell et al., 2004); these temperatures are substantially warmerthan contemporaneous pelagic bottom water temperatures estimatedfrom ODP sites 747 (55°S, 77°E; Billups and Schrag, 2003) and 1171C(~3 to 7 °C; Billups and Schrag, 2002; Shevenell et al., 2004). Our findingssuggest that the modern temperature gradient observed between thesouthern New Zealand Shelf (10 °C; this study) and Antarctic bottomwater (10 °C; Elderfield et al., 2010) was present during the Altonian,despite absolute temperatures being 3–4 °C cooler at present. Notethat our calculated temperatures are corrected for the Early MioceneMg/Ca seawater values summarised by Fantle and DePaolo (2006);different Mg/Ca values for ancient seawater could have a minor ef-fect on temperature values (e.g. up to 3 °C warmer; Fantle andDePaolo, 2006; Cramer et al., 2011). The seawater temperatures cal-culated by Billups and Schrag (2003) and Shevenell et al. (2004)were not corrected for an Early Miocene Mg/Ca seawater value.
The effect of diagenetic alteration is of utmost concern for interpreta-tion of signals from geochemical proxies. The growth of secondary car-bonate and dissolution of foraminiferal calcite may alter original Mg/Caratios (Lorens et al., 1977; Rosenthal and Boyle, 1993; Brown andElderfield, 1996; Erez, 2003). SEM images of surface and internal detailsconfirmed that the sampled fossils were clean and intact after thecleaning procedures (Fig. 8). Agreement between palaeotemperature es-timates from the 12 sample horizons gives confidence that the geochem-ical signals from the Mount Harris Formation foraminifera are primaryand not a result of alteration.
4.4. Ancient sea water
Mg/Ca temperature estimates and oxygen isotopic compositions ofN. spinosa test carbonate (δ18Oc) were used to calculate the oxygen iso-topic composition of seawater (δ18Opalaeo-sw) during the Altonian (e.g.Horibe and Oba, 1972). δ18Opalaeo-sw is an important parameter thathas provided insight into the fluctuation of continental ice volume,and the salinity and sea level of ancient oceans (e.g. Miller et al., 1987;Hollis et al., 2009). Sedimentological and micropalaeontological evi-dence suggest that theMount Harris Formation at Pukeuri accumulatedin anopenmarine shelf setting; there is no independent evidence of sig-nificant fresh water input from the gradually emerging but generally
low-relief Zealandia. Because major ocean currents comparable tothose of modern New Zealand were established before the Altonian(Nelson and Cooke, 2001), the water at Pukeuri was likely sourcedfrom the Southern Ocean, with a global oxygen isotopic signal little-influenced by major freshwater inputs within the 60 ka windowreported here. The palaeo-seawater isotopic compositions averaged−0.4±0.4‰ (VSMOW) throughout the studied sequence. Our resultsare consistent with δ18Opalaeo-sw values calculated from Altonian agedforaminifera at ODP sites 747 (Billups and Schrag, 2003) and 1171C(Shevenell et al., 2008). If the values here are considered to representthe global average for δ18Opalaeo-sw during the Early Miocene then thesignal implies reasonably ice-free conditions or the presence of onlysmall ice sheets (e.g. Kennett, 1986; Zachos et al., 2001, 2008). Palaeo-seawater values may represent a more local feature (e.g. localisedwater mass or current), but no other finely-sampled δ18Opalaeo-sw Mio-cene values for New Zealand are available for comparison. Note thatour palaeo-seawater isotopic compositions are influenced by an esti-mate of Early Miocene seawater Mg/Ca (i.e. Fantle and DePaolo,2006), and different estimates may give slightly different δ18Opalaeo-sw
values.Sedimentary and microfaunal proxies indicated that the pal-
aeoenvironmental setting was stable during relatively ice-free condi-tions, although small fluctuations between sample horizons 5 and8 (Anomaly 1) and between sample horizons 10 and 12 (Anomaly 2)may reflect increases in ice volume (Fig. 7). The increase in calculatedδ18Opalaeo-sw value (~0.7‰) of Anomaly 1 suggests a relative fall in sealevel and is supported by an increased sand fraction, reduction of theamount of carbonate and fewer planktonic foraminifera (Fig. 4). Thebrevity of the δ18Opalaeo-sw departure suggests that Anomaly 1 repre-sents a short period of rapid glacio-eustatic sea level change. In contrast,the percentage of planktonic foraminifera increased across Anomaly 2,indicating a decoupling or lag between δ18Opalaeo-sw and the relativeabundance of planktonic foraminifera. Irrespective, the similarity inδ18Opalaeo-sw excursions between the two anomalies suggests thatAnomaly 2 is also a response to a small glaciation event within theEarly Miocene.
5. Conclusions
Calibration curves were produced for benthic foraminifera by cor-relating Mg/Ca with known water temperatures. Intergeneric varia-tions were found between calibration curves developed for Cibicidesspp. and Notorotalia spp., with the Notorotalia calibration curvemore sensitive than the Cibicides curve between 10 and 15 °C. Themodern calibration curves were adjusted for ancient Mg/Ca seawatervalues and applied to fossil species of each genus from a 60 ka spanabout 17.7 Ma at Pukeuri, New Zealand. Palaeotemperature estimatesfrom the two different curves were concordant, although estimatesfrom N. spinosa (13.3±1.0 °C) were relatively constant while esti-mates from Cibicides (15.5±3.0 °C) were more varied. Both pal-aeotemperature estimates are consistent with warm-temperateoceans, indicating that Altonian bottom waters at the Pukeuri localitywere slightly warmer than the present day Otago Shelf. Oxygen isoto-pic composition was estimated for Altonian sea water (δ18Opalaeo-sw)using a paired Mg/Ca ratio and oxygen isotopic approach. δ18Opalaeo-sw
featured two small fluctuations that were hypothesised to representminor glaciations during otherwise relatively ice-free conditions.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.marmicro.2012.05.002.
Acknowledgements
We thank K. Swanson and J. Guise (University of Canterbury), N.Hiller (Canterbury Museum), B.A. Marshall (Museum of New Zealand),and O. Gussman (University of Otago) for providing modern materials.M. Uddstorm (NIWA) and B. Dickson (Portobello laboratory, University

Fig. 8. Scanning electron micrographs of fossil Notorotalia from horizon 2 of the Pukeuri study section. Images show foraminifera surfaces A) without cleaning, B) after sonicationand rinsing, and C) after all cleaning steps including acid leaching. The full suite of cleaning procedures was effective at removing secondary minerals.
59K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
of Otago) provided uswith seasonalwater temperature archive records.We thank R. Dewdney (Department of Geology, University of Otago) forhelping to prepare and pick samples.We thank B.W.Hayward, an anon-ymous reviewer, and the Editor for constructive comments. Supportfrom Department of Chemistry, Geology and Marine Science are grate-fully acknowledged. Financial support for this study was provided bya University of Otago Postgraduate Scholarship.
References
Beu, A.G., Maxwell, P.A., 1990. Cenozoic Mollusca of New Zealand. New Zealand Geo-logical Survey Paleontological Bulletin 58 (518 pp.).
Billups, K., Schrag, D.P., 2002. Paleotemperatures and ice volume of the past 27 Myrrevisited with paired Mg/Ca and 18O/16O measurements on benthic foraminifera.Paleoceanography 17, 3.1–3.11.
Billups, K., Schrag, D.P., 2003. Application of benthic foraminiferal Mg/Ca ratios to ques-tions of Cenozoic climate change. Earth and Planetary Science Letters 209,181–195.
Boyle, E.A., 1995. Limits on benthic foraminiferal chemical analyses as precise mea-sures of environmental properties. Journal of Foraminiferal Research 25, 4–13.
Brown, S.J., Elderfield, H., 1996. Variations in Mg/Ca and Sr/Ca ratios of planktonic fora-minifera caused by postdepositional dissolution: evidence of shallow Mg depen-dent dissolution. Paleoceanography 11, 543–551.
Buening, N., Carlson, S.J., Spero, H.J., Lee, D.E., 1998. Evidence for the Early Oligoceneformation of a proto-subtropical convergence from oxygen isotope records ofNew Zealand Paleogene brachiopods. Palaeogeography, Palaeoclimatology, Palaeo-ecology 138, 43–68.
Chapman, F., 1909. Report on the foraminifera from the subantarctic islands of NewZealand. In: Chilton, C. (Ed.), The subantarctic islands of New Zealand. Reports onthe geophysics, geology, zoology and botany of the islands lying to the south ofNew Zealand, based mainly on observations and collections made during an expe-dition in the Government Steamer “Hinemoa” (Captain J. Bollons) in November,1907. Philosophical Institute of Canterbury, Christchurch, pp. 312–371.
The New Zealand geological timescale. In: Cooper, R.A. (Ed.), Institute of Geological andNuclear Science Monograph 22 (284 pp.).
Cramer, B.S., Miller, K.G., Barrett, P.J., Wright, J.D., 2011. Late Cretaceous–Neogene trendsin deep ocean temperature and continental ice volume: reconciling records of ben-thic foraminiferal geochemistry (δ18O and Mg/Ca) with sea level history. Journal ofGeophysical Research 116, C12023, http://dx.doi.org/10.1029/2011JC007255.
Devereux, I., 1967. Oxygen isotope paleotemperature measurements on New ZealandTertiary fossils. New Zealand Journal of Science 10, 988–1011.
Dissard, D., Nehrke, G., Reichart, G.J., Bijma, J., 2010. The impact of salinity on the Mg/Caand Sr/Ca ratio in the benthic foraminifera Ammonia tepida: results from cultureexperiments. Biogeosciences 7, 81–93.
Elderfield, H., Ganssen, G., 2000. Past temperature and δ18O of surface ocean waters in-ferred from foraminiferal Mg/Ca ratios. Nature 405, 442–445.
Elderfield, H., Yu, J., Anand, P., Kiefer, T., Nyland, B., 2006. Calibrations for benthic fora-miniferal Mg/Ca paleothermometry and the carbonate ion hypothesis. Earth andPlanetary Science Letters 250, 633–649.
Elderfield, H., Greaves, M., Barker, S., Hall, I.R., Tripati, A., Ferretti, P., Crowhurst, S.,Booth, L., Daunt, C., 2010. A record of bottom water temperature and seawaterδ18O for the Southern Ocean over the past 440 kyr based on Mg/Ca of benthic fora-miniferal Uvigerina spp. Quaternary Science Reviews 29, 160–169.
Erez, J., 2003. The source of ions for biomineralization in foraminifera and their impli-cations for paleoceanographic proxies. In: Dove, P.M., De Yoreo, J.J., Weiner, S.(Eds.), Biomineralization. : Reviews in Mineralogy and Geochemistry, 54. Mineral-ogical Society of America, Washington, DC, pp. 115–149.
Fantle, M.S., DePaolo, D.J., 2006. Sr isotopes and pore fluid chemistry in carbonate sedi-ment of the Ontong Java Plateau: calcite recrystallization rates and evidence for arapid rise in seawaterMg over the last 10 million years. Geochimica et CosmochimicaActa 70, 3883–3904.
Field, B.D., Crundwell, M.P., Lyon, G.L., Mildenhall, D.C., Morgans, H.E.G., Ohneiser, C.,Wilson, G.S., Kennett, J.P., Chanier, F., 2009. Middle Miocene paleoclimate changeat Bryce Burn, southern New Zealand. New Zealand Journal of Geology and Geo-physics 52, 321–333.
Fukuda, K., 2002. Deducing the Cenozoic paleotemperature by using Mg/Ca ratios inbenthic foraminifera. MSc thesis, University of Otago, Dunedin.
Gradstein, F.M., Ogg, J.G., Smith, A.G., 2004. A Geologic Time Scale 2004. CambridgeUniversity Press, Cambridge. 610 pp.
Graham, I.J., Morgans, H.E.G., Waghorn, D.B., Trotter, J.A., Whitford, D.J., 2000. Stron-tium isotope stratigraphy of the Oligocene–Miocene Otekaike Limestone (Trig Zsection) in southern New Zealand: age of the Duntroonian/Waitakian stage bound-ary. New Zealand Journal of Geology and Geophysics 43, 335–347.
Hayward, B.W., 1977. Lower Miocene corals from the Waitakere Ranges, North Auck-land, New Zealand. Journal of the Royal Society of New Zealand 7, 99–111.
Hayward, B.W., 1986. A guide to paleoenvironmental assessment using New Zealand Ce-nozoic foraminiferal faunas. New Zealand Geological Survey Report 109 (73 pp.).
Hayward, B.W., Grenfell, H.R., Reid, C.M., Hayward, K.A., 1999. Recent New Zealandshallow water benthic foraminifera: taxonomy, ecologic distribution, biogeogra-phy and use in paleoenvironmental assessment. Institute of Geological and NuclearScience Monograph 21 (258 pp.).

60 K. Fukuda et al. / Marine Micropaleontology 92–93 (2012) 52–60
Heath, R.A., 1984. The depth of the mixed layer as an indicator of oceanic circulationaround New Zealand. New Zealand Journal of Marine and Freshwater Research18, 83–92.
Hollis, C., Handley, L., Crouch, E.M., Morgans, H.E.G., Baker, J.A., Creech, J., Collins, K.S.,Gibbs, S.J., Huber, M., Schouten, S., Zachos, J.C., Pancost, R.D., 2009. Tropical seatemperatures in the high-latitude South Pacific during the Eocene. Geology 37,99–102.
Hollis, C.J., Beu, A.G., Crampton, J.S., Crundwell, M.P., Morgans, H.E.G., Raine, J.I., Jones,C.M., Boyes, A.F., 2010. Calibration of the New Zealand Cretaceous–Cenozoic time-scale to GTS2004. GNS Science report 2010/43. 20 pp.
Horibe, Y., Oba, T., 1972. Temperature scales of aragonite–water and calcite–water sys-tems. Fossils 23/24, 69–74.
Hornibrook, N. de B., 1992. New Zealand Cenozoic marine paleoclimates: a reviewbased on the distribution of some shallow water and terrestrial biota. In: Tsuchi,R., Ingles, J.C. (Eds.), Pacific Neogene Environment, Evolution and Events. Universi-ty of Tokyo Press, Tokyo, pp. 83–106.
Hornibrook, N. de B., Brazier, R.C., Strong, C.P., 1989. Manual of New Zealand Permianto Pleistocene foraminiferal biostratigraphy. New Zealand Geological Survey Pale-ontological Bulletin 69, 45p.
Hut, G., 1987. Consultant's Group Meeting on Stable Isotope Reference Samples forGeochemical and Hydrological Investigations, September 16–18 1985, Vienna. In-ternational Atomic Energy Agency, Vienna. 43 pp.
Kennett, J.P., 1986. Miocene to early Pliocene oxygen and carbon isotope stratigraphyin the southwest Pacific, Deep Sea Drilling Project Leg 90. In: Kennett, J.P., vonder Borch, C.C. (Eds.), Initial Report of the Deep Sea Drilling Project Leg, 90. UnitedStates Government Printing Office, Washington, DC, pp. 1383–1411.
King, P.R., 2000. Tectonic reconstructions of New Zealand: 40 Ma to the Present. NewZealand Journal of Geology and Geophysics 43, 611–638.
Lear, C.H., Elderfield, H., Wilson, P.A., 2000. Cenozoic deep-sea temperatures and globalice volumes from Mg/Ca in benthic foraminiferal calcite. Science 287, 269–272.
Lear, C.H., Rosenthal, Y., Slowey, N., 2002. Benthic foraminiferal Mg/Ca-paleothermometry: arevised core-top calibration. Geochimica et Cosmochimica Acta 66, 3375–3387.
Lorens, R.B., Williams, D.F., Bender, M.L., 1977. The early nonstructural chemical dia-genesis of foraminiferal calcite. Journal of Sedimentary Petrology 47, 1602–1609.
McArthur, J.M., Howarth, R.J., Bailey, T.R., 2001. Strontium isotope stratigraphy: Lowessversion 3: best fit to the marine Sr-isotope curve for 0–509 Ma and accompanyinglook‐up table for deriving numerical age. Journal of Geology 109, 155–170.
Medina-Elizalde, M., Lea, D.W., Fantle, M.S., 2008. Implications of seawater Mg/Ca var-iability for Plio-Pleistocene tropical climate reconstruction. Earth and PlanetaryScience Letters 269, 584–594.
Miller, K.G., Fairbanks, R.G., Mountain, G.S., 1987. Tertiary oxygen isotope synthesis, sealevel history, and continental margin erosion. Paleoceanography 2, 1–19.
Morgans, H.E.G., Edwards, A.R., Scott, G.H., Graham, I.J., Kamp, P.J.J., Mumme, T.C.,Wilson, G.J., Wilson, G.S., 1999. Integrated stratigraphy of the Waitakian–Otaianstage boundary stratotype, Early Miocene, New Zealand. New Zealand Journal ofGeology and Geophysics 42, 581–614.
Morgans, H.E.G., Scott, G.H., Edwards, A.R., Graham, I.J., Mumme, T.C., Waghorn, D.B.,Wilson, G.S., 2002. Integrated stratigraphy of the lower Altonian (Early Miocene)sequence at Tangakaka Stream, East Cape, New Zealand. New Zealand Journal ofGeology and Geophysics 45, 145–173.
Mucci, A., Morse, J.W., 1983. The incorporation of Mg2+ and Sr2+ into calcite over-growths: influences of growth rate and solution composition. Geochimica etCosmochimica Acta 47, 217–233.
Nelson, C.S., Burns, D.A., 1982. Effect of sampling interval on the resolution of oxygenisotopic paleotemperature trends — an example from the New Zealand early Mio-cene. New Zealand Journal of Geology and Geophysics 25, 77–81.
Nelson, C.S., Cooke, P.J., 2001. History of oceanic front development in the New Zealandsector of the Southern Ocean during the Cenozoic— a synthesis. New Zealand Jour-nal of Geology and Geophysics 44, 535–553.
Nürnberg, D., Bijma, J., Hemleben, C., 1996. Assessing the reliability of magnesium inforaminiferal calcite as a proxy for water mass temperatures. Geochimica etCosmochimica Acta 60, 803–814.
Pickering, T.D., Suda, Y. (Eds.), 2003. Report of the 2nd Joint National Fisheries Univer-sity–University of the South Pacific Fisheries and Oceanography Research Cruise onboard Koyo-Maru, Kadavu Passage, December 1997. : Marine Studies technical re-port. University of the South Pacific. 69 pp.
Regenberg, M., Steph, S., Nürnberg, D., Tiedemann, R., Garbe-Schönberg, D., 2009. Cal-ibrating Mg/Ca ratios of multiple planktonic foraminiferal species with δ18O--calcification temperatures: paleothermometry for the upper water column. Earthand Planetary Science Letters 278, 324–336.
Ries, J.B., 2006. Mg fractionation in crustose coralline algae: geochemical, biological,and sedimentological implications of secular variation in the Mg/Ca ratio of seawa-ter. Geochimica et Cosmochimica Acta 70, 891–900.
Rosenthal, Y., Boyle, E.A., 1993. Factors controlling the fluoride content of planktonicforaminifera: an evaluation of its paleoceanographic applicability. Geochimica etCosmochimica Acta 57, 335–346.
Rosenthal, Y., Boyle, E.A., Slowey, N., 1997. Temperature control on the incorporation ofmagnesium, strontium, fluorine, and cadmium into benthic foraminiferal shellsfrom Little Bahama Bank: prospects for thermocline paleoceanography. Geo-chimica et Cosmochimica Acta 61, 3633–3643.
Rosenthal, Y., Lohmann, G.P., Sherrell, R.M., 2000. Incorporation and preservation of Mgin Globigerinoides sacculifer: implications for reconstructing the temperature and18O/16O of seawater. Paleoceanography 15, 135–145.
Rosenthal, Y., Lear, C.H., Oppo, D.W., Linsley, B.K., 2006. Temperature and carbonate ioneffects on Mg/Ca and Sr/Ca ratios in benthic foraminifera: the aragonitic speciesHoeglundina elegans. Paleoceanography 21, PA1007.
Shackleton, N., 1967. Oxygen isotope analyses and Pleistocene temperatures reassessed. Na-ture 215, 15–17.
Shevenell, A.E., Kennett, J.P., Lea, D.W., 2004. Middle Miocene Southern Ocean coolingand Antarctic cryosphere expansion. Science 305, 1766–1769.
Shevenell, A.E., Kennett, J.P., Lea, D.W., 2008. Middle Miocene ice sheet dynamics,deep-sea temperatures, and carbon cycling: a Southern Ocean perspective. Geo-chemistry, Geophysics, Geosystems 9, Q02006.
ter Kuile, B., 1991. Mechanisms for calcification and carbon cycling in algal symbiont-bearing foraminifera. In: Lee, J.J., Anderson, R. (Eds.), Biology of Foraminifera. Aca-demic Press, London, pp. 255–284.
Toyofuku, T., Kitazato, H., Kawahata, H., Tsuchiya, M., Nohara, M., 2000. Evaluation ofMg/Ca thermometry in foraminifera: comparison of experimental results and mea-surements in nature. Paleoceanography 15, 456–464.
Van Hale, R., 2003. The Stable Isotope Oceanography of the Otago Shelf. PhD thesis,University of Otago, Dunedin.
Walton, W.R., 1952. Techniques for recognition of living foraminifera. Contributionsfrom Cushman Foundation for Foraminiferal Research 3, 56–60.
Zachos, J.C., Pagani, M., Sloan, L., Thomas, E., Billups, K., 2001. Trends, rhythms, and ab-errations in global climate 65 Ma to Present. Science 292, 686–693.
Zachos, J.C., Dickens, G.R., Zeebe, R.E., 2008. An early Cenozoic perspective on green-house warming and carbon-cycle dynamics. Nature 451, 279–283.