and b. rapa reveals - hindawi publishing...
TRANSCRIPT

Research ArticleTranscriptome Analysis of InterspecificHybrid between Brassica napus and B rapa RevealsHeterosis for Oil Rape Improvement
Jinfang Zhang1 Guangrong Li2 Haojie Li1 Xiaobin Pu1 Jun Jiang1 Liang Chai1
Benchuan Zheng1 Cheng Cui1 Zujun Yang2 Yongqing Zhu3 and Liangcai Jiang1
1Crop Science Institute Sichuan Academy of Agricultural Sciences Chengdu Sichuan 6110066 China2School of Life Science and Technology University of Electronic Science and Technology of China Chengdu Sichuan 610054 China3Institute of Agro-Products Processing Science and Technology Sichuan Academy of Agricultural Sciences ChengduSichuan 6110066 China
Correspondence should be addressed to Liangcai Jiang jlcrape163com
Received 3 May 2015 Revised 3 August 2015 Accepted 5 August 2015
Academic Editor Graziano Pesole
Copyright copy 2015 Jinfang Zhang et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The hybrid between Brassica napus and B rapa displays obvious heterosis in both growth performance and stress tolerances Acomparative transcriptome analysis for B napus (AnAnCC genome) B rapa (ArAr genome) and its hybrid F1 (AnArC genome)was carried out to reveal the possiblemolecularmechanisms of heterosis at the gene expression level A total of 40320 nonredundantunigenes were identified using B rapa (AA genome) and B oleracea (CC genome) as reference genomes A total of 6816differentially expressed genes (DEGs)weremapped in theA andCgenomeswith 4946DEGs displayed nonadditively by comparingthe gene expression patterns among the three samples The coexistence of nonadditive DEGs including high-parent dominancelow-parent dominance overdominance and underdominance was observed in the gene action modes of F1 hybrid which werepotentially related to the heterosis The coexistence of multiple gene actions in the hybrid was observed and provided a list ofcandidate genes and pathways for heterosis The expression bias of transposable element-associated genes was also observed inthe hybrid compared to their parents The present study could be helpful for the better understanding of the determination andregulation of mechanisms of heterosis to aid Brassica improvement
1 Introduction
Brassica napus (rapeseed AACC 2119899 = 38) is one of the mainoil crops used for human consumption and is widely grownin China Canada Europe and Australia and increasinglygrown in South America [1] B napus is likely derived fromthe natural hybridization and genome doubling from its twodiploid parentsB rapa (genomeAA 2119899 = 20) andB oleracea(genome CC 2119899 = 18) along the Mediterranean coastline inSouthern Europe approximately 10000 years ago [2] How-ever there is evidence that the genetic diversity in rapeseedhas been continuously reduced by extensive breeding efforts[3] Approaches to increase the genetic variation of rapeseedhave beenmade by introducing the genetic components from
related species as B napus can easily cross with one of its an-cestral parentsB rapa to produce viable interspecies hybrids[4] Due to the high crossability betweenB napus andB rapaand low aneuploidy of their interspecific hybrids differentB rapa accessions have been widely hybridized to rapeseedbreeding program and novel agronomic traits from B rapahave been successfully transmitted into commercial B napusvarieties [5] Strong heteroses affecting both biomass andseed yield have been observed in hybrids derived from theseinterspecific crosses [6] However the genetic and molecularmechanism of heterosis in the interspecific hybrids has notbeen investigated
Heterosis is a prevalent phenomenon in evolution andbreeding process of plants [7] The hybrid offspring obtains
Hindawi Publishing CorporationInternational Journal of GenomicsVolume 2015 Article ID 230985 11 pageshttpdxdoiorg1011552015230985
2 International Journal of Genomics
the advantages in many agricultural and developmental traitsincluding biomass yield plant height vigor and stress toler-ances from their parents thereby improving the adaptationof the crop and subsequently increasing the global area sown[8] Although heterosis has been widely utilized the geneticand molecular basis of heterotic improvements remainsunclear Various genetic models have been proposed forexplaining heterosis and include dominance overdominanceand pseudo overdominance [9] Heterotic interactions inthe tetraploid B napus can be derived from two types ofallelic interactions namely those between the genomes ofthe two diploid progenitors as well as those between thetwo parental genomes making up a hybrid Gene expressionis potentially altered not only due to a dosage effect butalso with gene functional divergence [8] Epigenetic mod-ification in hybrids genomes can also account for hetero-sis [10] Emerging new technologies make it possible toinvestigate the DNA sequence RNA transcripts proteinsand metabolism products genome-wide allowing for newinsights into benefits of heterotic recombinations [7 9] Nextgeneration sequencing technology utilizing RNA-seq hasemerged as a powerful tool for investigating gene expressiondata at the whole genome level Both the gene transcriptand expression levels can be detected in RNA-seq whichcould provide new insights into themolecularmechanisms ofheterosis [11]
Chinese cabbage Brassica rapa (syn B campestris) con-tains two subspecies named pekinensis and chinensis Thesetwo distinct groups are used as leafy vegetables widely acrosseastern Asia and have much genetic variation In particularB rapa ssp chinensis has wide morphological variation andis found in China Korea and Japan [12] Recently a numberof genes responsible for developmental and stress toleranceswere isolated [13ndash15] showing thatB rapa ssp chinensis couldbe a novel resource for B napus improvement
In this study the transcriptomes of B rapa ssp chinen-sis Makino B napus and their interspecific hybrid weresequenced to investigate the molecular basis of heterosisGene expression profiles were used to identify the numberof differentially expressed genes and gene expression levelswere analyzed and compared among two parents and theirhybrid This data was used to give a better understandingof additive and nonadditive effects of genes and how thesecontribute to heterosis thus providing new insights into thegenetic and epigenetic mechanism of subgenomic heterosisin B napus
2 Materials and Methods
21 Sample Preparation and RNA Isolation Three lines weregrown in a green house at 22∘C under 12 hours of light and12 hours of darkness Leaves of the seedling were harvestedafter 4 weeks by cutting off the sixth leaf of each seedlingThe leaves of four plants were combined for RNA extractionSamples were immediately frozen in liquid nitrogen andstored at minus80∘C until RNA was extracted Total RNA of eachsample was isolated using RNAprep Pure Plant Kit (TiangenShanghai China) RNA quality was characterized initially
by NanoDrop ND1000 spectrophotometer (NanoDrop Tech-nologies Wilmington DE USA) and then further assessedby RIN (RNA Integrity Number) value (gt95) using Agilent2100 Bioanalyzer (Santa Clara CA USA) Equal quantitiesof high-quality RNA from each sample were pooled forcDNA synthesis Cytological observation of the F1 hybrid wasperformed as described by Zhan et al [16]
22 cDNA Library Construction for Illumina Sequencing ThecDNA library was constructed following the manufacturerrsquosinstructions of mRNA-Seq Sample Preparation Kit (catnumber RS-930-1001 Illumina Inc San Diego CA) Brieflythe poly(A)mRNAwas isolated from total RNA samples withMagnetic Oligo (dT) BeadsThemRNAwas then fragmentedinto small pieces using RNA fragmentation kit (Ambion)Using these short fragments as the templates the first cDNAstrand was synthesized using random hexamer primers andreverse transcriptase (Invitrogen) and the second-strandcDNA was synthesized using DNA polymerase I and RNaseH The cDNA fragments were purified using the QiaQuickPCR extraction kit (Qiagen) and resolved with EB buffer forend reparation and poly(A) addition The short fragmentswere then connected with sequencing adapters and theproducts were subsequently purified and amplified via PCRLibraries were prepared from a 400ndash500 bp size-selectedfraction following adapter ligation and agarose gel separationThe quality control analysis on the sample library wasperformed to quantify the DNA concentration and validatethe library After validation with an Eppendorf MastercyclerReal-Time PCR System the cDNA libraries were sequencedon the Illumina Hiseq 2000 platform with read length of2 times 100 bp The sequencing-derived raw image data weretransformedby base calling into sequence data using IlluminaPipeline Software v16 The raw sequencing reads have beensubmitted to NCBI Short Read Archive under the accessionnumber of SRR2131203 SRR2134440 and SRR2134444
23 Sequence Data Analysis and Expression Analysis Theraw reads were cleaned by removing adapter sequences low-quality sequences (reads with ambiguous bases ldquo119873rdquo) andreadswithmore than 10 119876 lt 20 bases To analyze transcriptabundance levels the uniquely mapped reads for a specificgene were counted by mapping reads to de novo assembleddistinct sequences using SOAP2 software [17] and the RPKM(Reads Per Kb per Million reads) values were computed asproposed byMortazavi et al [18] Gene transcript abundancedifferences were obtained fromRPKMvalues using amethodmodified from Audicrsquos proposal [19] Gene expression levelswere calculatedwith the RPKMmethodwhich is able to elim-inate the influence of different gene lengths and sequencingdiscrepancy within the calculation of gene expression
24 Statistics Statistical analysis of the phenotypic datawas performed with a Welch two-sample 119905-test in R (RDevelopment Core Team 2005) [20] Midparent heterosis(MPH) values were calculated using the formula MPH =(hminus (P1 + P2)2)((P1 + P2)2) lowast 100 + 100 (whereas h is thevalue of the hybrid and P1 and P2 are the values of the one
International Journal of Genomics 3
50 120583m
Figure 1 Root tip squash showing chromosomes from the F1 hybrid (chromosome number of 29)
(a) (b)
Figure 2 Seed germination (a) and 2-week-old seedlings (b) of P1 P2 and F1 germplasm
and the other parental line) To test for significance of MPHvalues the contrast h minus (P1 + P2)2 was used [21]
25 Quantitative RT-PCR Validation of Differentiation GeneExpression RNA-seq datawere further validated using quan-titative real-time PCR analysis (qRT-PCR) for a selectednumber of genes using gene-specific primer sets The plantleaves and growth condition for total RNA extraction usedfor qRT-PCR were the same as these for RNA-seq experi-ments Three independent RNA extractions per sample wereperformed for biological replicates The reverse transcribedinto cDNA using PrimeScritH RT reagent kit with gDNAEraser (Takara Dalian China) Primer pairs of 10 uni-genes were designed using Invitrogenrsquos (Carlsbad CA USA)OligoPerfect Designer software [22] Specificity of the primersets and their product length was verified by agarose gelelectrophoresis The qRT-PCR reaction mixture consisted ofthe SYBR Premix EX-Taq II Kit (TakaRa Japan) on iCycleriQ (BIO-RAD) for three repeats of each sample The thermalcycling conditions were as follows 95 uC 2min and 40 cyclesat 95∘C for 10 s for denaturation and 65∘C 20 s and 72∘C 30 sfor annealing and extension The expression of TIPS-41 withprimer sequences 51015840-TGAAGAGCAGATTGATTTGGCT-31015840and 51015840-ACACTCCATTGTC AGCCAGTT-31015840 was used asan internal control for normalization to compare the geneexpression level between the accessions The relative levels ofgene expression were calculated using the 2minusΔΔCt method
3 Results
31 Sample Preparation Sequencing and Data Filtering Thehybridization of B napus cv CWH-2 (P1) and B rapa ssp
chinensis cv Qianjin (P2) was made in 2010-2011 oilseedgrowing season Root-tip chromosome assessments of F1showed that all F1 plants had a chromosome number of29 (Figure 1) suggesting that they were hybrids The seedsrsquosizes of F1 hybrids were intermediate to those of the parentsbut the seed color resembled that of B napus (Figure 2(a))Root growth of the F1 over the first 24 h was similar toP1 Leaf morphology of the F1 was more similar to B rapathan to B napus (Figure 2) In 4-week-old seedlings theF1 plant displayed stronger roots than either of the parents(Figure 2(b)) while adult F1 plants displayed the similarheading time to B napus
To identify transcripts of P1 P2 and F1 three cDNAlibraries were constructed from their leaves and sequenced byIllumina paired-end sequencing Sequence data from each ofthe libraries consisted of 60Gb from B rapa (AA genome)52 Gb from B napus (AACC genome) and 67Gb from thehybrid (AAC genome) High-quality cleaned raw reads fromall of those data were aligned to reference sequences In total67951208 reads and 46207184 reads were mapped in theB rapa and B oleracea reference genomes respectively Inthe B rapa genome 38522 unigenes were assembled witha minimum scaffold size of 200 bp and a total length of51995312 bp with an average length of 1349 bp (Figure 3(a))The B oleracea genome had a total of 42009 unigeneswith a minimum scaffold size of 300 bp a total length of51691329 bp and an average length of 1230 bp (Figure 3(b))
32 Heterosis of Differential Gene Expression in B rapa Bnapus and Their Hybrid The numbers of expressed genein each genome varied slightly There were 29579 (718)27939 (679) and 29447 (717) from B rapa B napus
4 International Journal of Genomics
0
500
1000
1500
2000
2500
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(a)
0500
1000150020002500300035004000
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(b)
Figure 3 The size distribution of transcriptomic unigenes assembled by the B rapa (a) and B oleracea (b)
and the hybrid genome respectively The unique mappingrates were similar among three Brassica samples suggestingthat their genomes were quite alike Some genes had expres-sion that was germplasm specific The B rapa genome hadapproximately about 6 (1639) of genes that were uniquewhilst B napus and the hybrid had only 2 (420) and3 (951) unique sequences respectively There were 25985genes expressed in all three lines A total of 489 genes wereexpressed in both B rapa and B napus but silenced in thehybrid Gene expression profiles of the three lines revealedthat the number of expressed genes did not show obviousdifference in the three sets of material The mode of geneaction for the differentially expressed genes (DEGs) wasanalyzed About 2700 genes (1840 of 6816) exhibited anexpression pattern that was not distinguishable from additiv-ity while the other 7256 (4946 of 6816) of genes showednonadditive expression patterns (Table 1) The nonadditivenumber ofDEGs from the crosswas further classified into fivedistinct classes high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) underdominance(UDO) and partial-dominanceThere were 7 of transcripts(545) that were absent in their parents indicating that therewas transcriptional activation of new genes in hybrid F1There was also transcriptional silencing with 32 (218) ofparentally expressed gene not being expressed in the hybrid
There were 2894 unigenes that mapped to the A genome2107 that mapped to the C genome (Figure 4) and 593that were not assigned to either genome Chromosomaldistribution of DEG numbers in hybrids showed that low-parent domiance (LPD) genes were significantly accumulatedon chromosomes A06 and C07 (119875 lt 0001) while high-parent dominance (HPD) genes were on chromosomes A09and C08 (119875 lt 001) Gene expression variation causedby interspecific hybridization was not randomly distributedalong the chromosomesTherefore differentB napus-B rapachromosome introgression lines can be developed to the fur-ther study of heterosis pertaining to specific chromosomes
33 Contribution of A and C Subgenomes to Gene DifferentialExpression in the Hybrid There were 1386 significant differ-entially expressed genes between P1 and F1 samples 858genes had increased expression in the F1 and 528 genes
Table 1 Summary of dominance patterns of differentially expressedgenes
DGs Number PercentageAdditivity 1840 27Nonadditivity 4946 7256Others 30 044High-parent dominance(HPD) 353 714
Low-parent dominance(LPD) 191 386
Overdominance (ODO) 593 1199Underdominance (UDO) 193 39Positive partial-dominance(PPD) 1635 3306
Negativepartial-dominance (NPD) 1981 405
Total 6816Additivity F1 asymp 12(P1 + P2) nonadditivity F1 gt 12(P1 + P2) or F1 lt 12(P1 +P2) High-parent dominance (HPD) F1asymp P1gt P2 or F1asymp P2lt P1 low-parentdominance (LPD) F1 asymp P1 lt P2 or F1 asymp P2 lt P1 overdominance (ODO) F1gt P1 and F1 gt P2 underdominance (UDO) F1 lt P1 and F1 lt P2
had decreased expression There were 5077 significant dif-ferentially expressed genes between P2 and F1 samples with3241 genes having increased expression in the F1 and 1836genes being decreased in expression (Figure 5) Changesto expression from P2 were approximately three times thenumber of genes compared to from P1 There were moregenes with increased expression in the F1 (4098) than withdecreased expression (2464) when compared to expressionlevels in the respective parents We performed gene ontology(GO) enrichment analysis to test the functional categoriesfor DEGs among P1 P2 and F1 (Figure 6) by AgriGO onlinetool [23] and R statistical software [20] There were 1012 and 18 functional categories of these transcripts thatbelonged to the cell component (CC) molecular function(MF) and biological process (BP) respectively The GOanalysis showed that the composition of functional pathwaysassociated with genes showing differential expression was
International Journal of Genomics 5
0
20
40
60
80
100
120
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10
LPDHPD
UDOODO
(a)
0
20
40
60
80
100
120
140
C1 C2 C3 C4 C5 C6 C7 C8 C9
LPDHPD
UDOODO
(b)
Figure 4 Chromosomal distribution of nonadditive differentially expressed genes with high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) and underdominance (UDO) on the A genome (a) and C genome (b)
P1_versus_F1F1_versus_P2
P1_versus_P2
694
310
3765
308
971
402
366
(a)
3000
2000
1000
0
858
528
1836
3241
2024
3422
RegulationUpDown
F1_versus_P1 F1_versus_P2 P1_versus_P2
(b)
Figure 5 Total numbers of differentially expressed genes between P1 P2 and F1 by Venn diagram analyses (a) and statistics of up- ordownregulated genes (b)
similar when comparing changes from either P1 or P2 withF1
The biological process category was further subdividedinto functional classes in those DEGs with high-parent
dominance (HPD 119875 lt 05) based on gene ontology Table 2highlights the 18 gene ontology classes that were differentiallyexpressed in the F1 Combined with the results in Figure 6we found that the GOs such as regulation of transcription
6 International Journal of Genomics
Response to chitinRegulation of plant-type hypersensitive responseResponse to woundingRespiratory burst involved in defense responseProtein targeting to membraneEthylene biosynthetic processJasmonic acid mediated signaling pathwayDefense response to fungusRegulation of hydrogen peroxide metabolic processSystemic acquired resistance salicylic acid mediated signaling pathwayAbscisic acid-activated signaling pathwayMAPK cascadeIntracellular signal transductionIndoleacetic acid biosynthetic processResponse to jasmonic acidResponse to ethyleneHyperosmotic salinity responseNegative regulation of defense responsePositive regulation of flavonoid biosynthetic processResponse to water deprivationDefense response to bacteriumGlucosinolate catabolic processResponse to mechanical stimulusResponse to endoplasmic reticulum stressDetection of biotic stimulusResponse to herbivoreResponse to bacteriumResponse to cyclopentenoneJasmonic acid biosynthetic processResponse to fungus
1
08
06
04
02
005
0
F1_v
ersu
s_P2
P1_v
ersu
s_P2
P1_v
ersu
s_F1
Figure 6 The heat map from GO enrichment analysis of DEGs between the P1 P2 and F1 combinations Color scale represents 119875 values ofenrichment test
metabolic process defense response multicellular organ-ismal development and transporters were enriched in Agenomes transcripts The genes responsible for stress werestriking enriched in the differential expressed genes fromC genome It possibly implied that the hybrid expressedprotein-coding genes from A genomes of both B rapaand B napus potentially participated in metabolism anddevelopment while those from C genome of B napus werelargely involved in stress resistance
34 TransposonActive or Inactive inHybrid Compared to theB rapa genome there were 25 transposable elements (TE)like genes expressed in P1 and P2 while the 11 (44) genesdisplayed differential expression in F1 (Table 3) The EnSpmtype of transposon and the retrotransposon-like transcriptwere significantly enriched It is likely that the interspecific
hybrids significantly modify the transcription of transposon-like genes which may directly or indirectly influence geneexpression of other genes in the heterotic material
35 Validation of Transcriptome Data by qRT-PCR Toassess the accuracy of RNA-seq data ten differentiallyexpressed unigenes including stress responsive genes sec-ondary metabolism biosynthesis genes and epigenetic mod-ifying genes were selected Three genes belong to under-dominance (UDO) and seven genes are belonging to over-dominance (ODO) types which are represented for heterosisanalysis Primer pairs of 10 unigenes for qRT-PCR weredesigned and listed in Table 4 We tested the similaritybetween differential gene expression identified by transcrip-tome and those identified by qRT-PCR As shown in Figure 7the qRT-PCR revealed that 8 of 10 genes (except Bol022348
International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 International Journal of Genomics
the advantages in many agricultural and developmental traitsincluding biomass yield plant height vigor and stress toler-ances from their parents thereby improving the adaptationof the crop and subsequently increasing the global area sown[8] Although heterosis has been widely utilized the geneticand molecular basis of heterotic improvements remainsunclear Various genetic models have been proposed forexplaining heterosis and include dominance overdominanceand pseudo overdominance [9] Heterotic interactions inthe tetraploid B napus can be derived from two types ofallelic interactions namely those between the genomes ofthe two diploid progenitors as well as those between thetwo parental genomes making up a hybrid Gene expressionis potentially altered not only due to a dosage effect butalso with gene functional divergence [8] Epigenetic mod-ification in hybrids genomes can also account for hetero-sis [10] Emerging new technologies make it possible toinvestigate the DNA sequence RNA transcripts proteinsand metabolism products genome-wide allowing for newinsights into benefits of heterotic recombinations [7 9] Nextgeneration sequencing technology utilizing RNA-seq hasemerged as a powerful tool for investigating gene expressiondata at the whole genome level Both the gene transcriptand expression levels can be detected in RNA-seq whichcould provide new insights into themolecularmechanisms ofheterosis [11]
Chinese cabbage Brassica rapa (syn B campestris) con-tains two subspecies named pekinensis and chinensis Thesetwo distinct groups are used as leafy vegetables widely acrosseastern Asia and have much genetic variation In particularB rapa ssp chinensis has wide morphological variation andis found in China Korea and Japan [12] Recently a numberof genes responsible for developmental and stress toleranceswere isolated [13ndash15] showing thatB rapa ssp chinensis couldbe a novel resource for B napus improvement
In this study the transcriptomes of B rapa ssp chinen-sis Makino B napus and their interspecific hybrid weresequenced to investigate the molecular basis of heterosisGene expression profiles were used to identify the numberof differentially expressed genes and gene expression levelswere analyzed and compared among two parents and theirhybrid This data was used to give a better understandingof additive and nonadditive effects of genes and how thesecontribute to heterosis thus providing new insights into thegenetic and epigenetic mechanism of subgenomic heterosisin B napus
2 Materials and Methods
21 Sample Preparation and RNA Isolation Three lines weregrown in a green house at 22∘C under 12 hours of light and12 hours of darkness Leaves of the seedling were harvestedafter 4 weeks by cutting off the sixth leaf of each seedlingThe leaves of four plants were combined for RNA extractionSamples were immediately frozen in liquid nitrogen andstored at minus80∘C until RNA was extracted Total RNA of eachsample was isolated using RNAprep Pure Plant Kit (TiangenShanghai China) RNA quality was characterized initially
by NanoDrop ND1000 spectrophotometer (NanoDrop Tech-nologies Wilmington DE USA) and then further assessedby RIN (RNA Integrity Number) value (gt95) using Agilent2100 Bioanalyzer (Santa Clara CA USA) Equal quantitiesof high-quality RNA from each sample were pooled forcDNA synthesis Cytological observation of the F1 hybrid wasperformed as described by Zhan et al [16]
22 cDNA Library Construction for Illumina Sequencing ThecDNA library was constructed following the manufacturerrsquosinstructions of mRNA-Seq Sample Preparation Kit (catnumber RS-930-1001 Illumina Inc San Diego CA) Brieflythe poly(A)mRNAwas isolated from total RNA samples withMagnetic Oligo (dT) BeadsThemRNAwas then fragmentedinto small pieces using RNA fragmentation kit (Ambion)Using these short fragments as the templates the first cDNAstrand was synthesized using random hexamer primers andreverse transcriptase (Invitrogen) and the second-strandcDNA was synthesized using DNA polymerase I and RNaseH The cDNA fragments were purified using the QiaQuickPCR extraction kit (Qiagen) and resolved with EB buffer forend reparation and poly(A) addition The short fragmentswere then connected with sequencing adapters and theproducts were subsequently purified and amplified via PCRLibraries were prepared from a 400ndash500 bp size-selectedfraction following adapter ligation and agarose gel separationThe quality control analysis on the sample library wasperformed to quantify the DNA concentration and validatethe library After validation with an Eppendorf MastercyclerReal-Time PCR System the cDNA libraries were sequencedon the Illumina Hiseq 2000 platform with read length of2 times 100 bp The sequencing-derived raw image data weretransformedby base calling into sequence data using IlluminaPipeline Software v16 The raw sequencing reads have beensubmitted to NCBI Short Read Archive under the accessionnumber of SRR2131203 SRR2134440 and SRR2134444
23 Sequence Data Analysis and Expression Analysis Theraw reads were cleaned by removing adapter sequences low-quality sequences (reads with ambiguous bases ldquo119873rdquo) andreadswithmore than 10 119876 lt 20 bases To analyze transcriptabundance levels the uniquely mapped reads for a specificgene were counted by mapping reads to de novo assembleddistinct sequences using SOAP2 software [17] and the RPKM(Reads Per Kb per Million reads) values were computed asproposed byMortazavi et al [18] Gene transcript abundancedifferences were obtained fromRPKMvalues using amethodmodified from Audicrsquos proposal [19] Gene expression levelswere calculatedwith the RPKMmethodwhich is able to elim-inate the influence of different gene lengths and sequencingdiscrepancy within the calculation of gene expression
24 Statistics Statistical analysis of the phenotypic datawas performed with a Welch two-sample 119905-test in R (RDevelopment Core Team 2005) [20] Midparent heterosis(MPH) values were calculated using the formula MPH =(hminus (P1 + P2)2)((P1 + P2)2) lowast 100 + 100 (whereas h is thevalue of the hybrid and P1 and P2 are the values of the one
International Journal of Genomics 3
50 120583m
Figure 1 Root tip squash showing chromosomes from the F1 hybrid (chromosome number of 29)
(a) (b)
Figure 2 Seed germination (a) and 2-week-old seedlings (b) of P1 P2 and F1 germplasm
and the other parental line) To test for significance of MPHvalues the contrast h minus (P1 + P2)2 was used [21]
25 Quantitative RT-PCR Validation of Differentiation GeneExpression RNA-seq datawere further validated using quan-titative real-time PCR analysis (qRT-PCR) for a selectednumber of genes using gene-specific primer sets The plantleaves and growth condition for total RNA extraction usedfor qRT-PCR were the same as these for RNA-seq experi-ments Three independent RNA extractions per sample wereperformed for biological replicates The reverse transcribedinto cDNA using PrimeScritH RT reagent kit with gDNAEraser (Takara Dalian China) Primer pairs of 10 uni-genes were designed using Invitrogenrsquos (Carlsbad CA USA)OligoPerfect Designer software [22] Specificity of the primersets and their product length was verified by agarose gelelectrophoresis The qRT-PCR reaction mixture consisted ofthe SYBR Premix EX-Taq II Kit (TakaRa Japan) on iCycleriQ (BIO-RAD) for three repeats of each sample The thermalcycling conditions were as follows 95 uC 2min and 40 cyclesat 95∘C for 10 s for denaturation and 65∘C 20 s and 72∘C 30 sfor annealing and extension The expression of TIPS-41 withprimer sequences 51015840-TGAAGAGCAGATTGATTTGGCT-31015840and 51015840-ACACTCCATTGTC AGCCAGTT-31015840 was used asan internal control for normalization to compare the geneexpression level between the accessions The relative levels ofgene expression were calculated using the 2minusΔΔCt method
3 Results
31 Sample Preparation Sequencing and Data Filtering Thehybridization of B napus cv CWH-2 (P1) and B rapa ssp
chinensis cv Qianjin (P2) was made in 2010-2011 oilseedgrowing season Root-tip chromosome assessments of F1showed that all F1 plants had a chromosome number of29 (Figure 1) suggesting that they were hybrids The seedsrsquosizes of F1 hybrids were intermediate to those of the parentsbut the seed color resembled that of B napus (Figure 2(a))Root growth of the F1 over the first 24 h was similar toP1 Leaf morphology of the F1 was more similar to B rapathan to B napus (Figure 2) In 4-week-old seedlings theF1 plant displayed stronger roots than either of the parents(Figure 2(b)) while adult F1 plants displayed the similarheading time to B napus
To identify transcripts of P1 P2 and F1 three cDNAlibraries were constructed from their leaves and sequenced byIllumina paired-end sequencing Sequence data from each ofthe libraries consisted of 60Gb from B rapa (AA genome)52 Gb from B napus (AACC genome) and 67Gb from thehybrid (AAC genome) High-quality cleaned raw reads fromall of those data were aligned to reference sequences In total67951208 reads and 46207184 reads were mapped in theB rapa and B oleracea reference genomes respectively Inthe B rapa genome 38522 unigenes were assembled witha minimum scaffold size of 200 bp and a total length of51995312 bp with an average length of 1349 bp (Figure 3(a))The B oleracea genome had a total of 42009 unigeneswith a minimum scaffold size of 300 bp a total length of51691329 bp and an average length of 1230 bp (Figure 3(b))
32 Heterosis of Differential Gene Expression in B rapa Bnapus and Their Hybrid The numbers of expressed genein each genome varied slightly There were 29579 (718)27939 (679) and 29447 (717) from B rapa B napus
4 International Journal of Genomics
0
500
1000
1500
2000
2500
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(a)
0500
1000150020002500300035004000
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(b)
Figure 3 The size distribution of transcriptomic unigenes assembled by the B rapa (a) and B oleracea (b)
and the hybrid genome respectively The unique mappingrates were similar among three Brassica samples suggestingthat their genomes were quite alike Some genes had expres-sion that was germplasm specific The B rapa genome hadapproximately about 6 (1639) of genes that were uniquewhilst B napus and the hybrid had only 2 (420) and3 (951) unique sequences respectively There were 25985genes expressed in all three lines A total of 489 genes wereexpressed in both B rapa and B napus but silenced in thehybrid Gene expression profiles of the three lines revealedthat the number of expressed genes did not show obviousdifference in the three sets of material The mode of geneaction for the differentially expressed genes (DEGs) wasanalyzed About 2700 genes (1840 of 6816) exhibited anexpression pattern that was not distinguishable from additiv-ity while the other 7256 (4946 of 6816) of genes showednonadditive expression patterns (Table 1) The nonadditivenumber ofDEGs from the crosswas further classified into fivedistinct classes high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) underdominance(UDO) and partial-dominanceThere were 7 of transcripts(545) that were absent in their parents indicating that therewas transcriptional activation of new genes in hybrid F1There was also transcriptional silencing with 32 (218) ofparentally expressed gene not being expressed in the hybrid
There were 2894 unigenes that mapped to the A genome2107 that mapped to the C genome (Figure 4) and 593that were not assigned to either genome Chromosomaldistribution of DEG numbers in hybrids showed that low-parent domiance (LPD) genes were significantly accumulatedon chromosomes A06 and C07 (119875 lt 0001) while high-parent dominance (HPD) genes were on chromosomes A09and C08 (119875 lt 001) Gene expression variation causedby interspecific hybridization was not randomly distributedalong the chromosomesTherefore differentB napus-B rapachromosome introgression lines can be developed to the fur-ther study of heterosis pertaining to specific chromosomes
33 Contribution of A and C Subgenomes to Gene DifferentialExpression in the Hybrid There were 1386 significant differ-entially expressed genes between P1 and F1 samples 858genes had increased expression in the F1 and 528 genes
Table 1 Summary of dominance patterns of differentially expressedgenes
DGs Number PercentageAdditivity 1840 27Nonadditivity 4946 7256Others 30 044High-parent dominance(HPD) 353 714
Low-parent dominance(LPD) 191 386
Overdominance (ODO) 593 1199Underdominance (UDO) 193 39Positive partial-dominance(PPD) 1635 3306
Negativepartial-dominance (NPD) 1981 405
Total 6816Additivity F1 asymp 12(P1 + P2) nonadditivity F1 gt 12(P1 + P2) or F1 lt 12(P1 +P2) High-parent dominance (HPD) F1asymp P1gt P2 or F1asymp P2lt P1 low-parentdominance (LPD) F1 asymp P1 lt P2 or F1 asymp P2 lt P1 overdominance (ODO) F1gt P1 and F1 gt P2 underdominance (UDO) F1 lt P1 and F1 lt P2
had decreased expression There were 5077 significant dif-ferentially expressed genes between P2 and F1 samples with3241 genes having increased expression in the F1 and 1836genes being decreased in expression (Figure 5) Changesto expression from P2 were approximately three times thenumber of genes compared to from P1 There were moregenes with increased expression in the F1 (4098) than withdecreased expression (2464) when compared to expressionlevels in the respective parents We performed gene ontology(GO) enrichment analysis to test the functional categoriesfor DEGs among P1 P2 and F1 (Figure 6) by AgriGO onlinetool [23] and R statistical software [20] There were 1012 and 18 functional categories of these transcripts thatbelonged to the cell component (CC) molecular function(MF) and biological process (BP) respectively The GOanalysis showed that the composition of functional pathwaysassociated with genes showing differential expression was
International Journal of Genomics 5
0
20
40
60
80
100
120
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10
LPDHPD
UDOODO
(a)
0
20
40
60
80
100
120
140
C1 C2 C3 C4 C5 C6 C7 C8 C9
LPDHPD
UDOODO
(b)
Figure 4 Chromosomal distribution of nonadditive differentially expressed genes with high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) and underdominance (UDO) on the A genome (a) and C genome (b)
P1_versus_F1F1_versus_P2
P1_versus_P2
694
310
3765
308
971
402
366
(a)
3000
2000
1000
0
858
528
1836
3241
2024
3422
RegulationUpDown
F1_versus_P1 F1_versus_P2 P1_versus_P2
(b)
Figure 5 Total numbers of differentially expressed genes between P1 P2 and F1 by Venn diagram analyses (a) and statistics of up- ordownregulated genes (b)
similar when comparing changes from either P1 or P2 withF1
The biological process category was further subdividedinto functional classes in those DEGs with high-parent
dominance (HPD 119875 lt 05) based on gene ontology Table 2highlights the 18 gene ontology classes that were differentiallyexpressed in the F1 Combined with the results in Figure 6we found that the GOs such as regulation of transcription
6 International Journal of Genomics
Response to chitinRegulation of plant-type hypersensitive responseResponse to woundingRespiratory burst involved in defense responseProtein targeting to membraneEthylene biosynthetic processJasmonic acid mediated signaling pathwayDefense response to fungusRegulation of hydrogen peroxide metabolic processSystemic acquired resistance salicylic acid mediated signaling pathwayAbscisic acid-activated signaling pathwayMAPK cascadeIntracellular signal transductionIndoleacetic acid biosynthetic processResponse to jasmonic acidResponse to ethyleneHyperosmotic salinity responseNegative regulation of defense responsePositive regulation of flavonoid biosynthetic processResponse to water deprivationDefense response to bacteriumGlucosinolate catabolic processResponse to mechanical stimulusResponse to endoplasmic reticulum stressDetection of biotic stimulusResponse to herbivoreResponse to bacteriumResponse to cyclopentenoneJasmonic acid biosynthetic processResponse to fungus
1
08
06
04
02
005
0
F1_v
ersu
s_P2
P1_v
ersu
s_P2
P1_v
ersu
s_F1
Figure 6 The heat map from GO enrichment analysis of DEGs between the P1 P2 and F1 combinations Color scale represents 119875 values ofenrichment test
metabolic process defense response multicellular organ-ismal development and transporters were enriched in Agenomes transcripts The genes responsible for stress werestriking enriched in the differential expressed genes fromC genome It possibly implied that the hybrid expressedprotein-coding genes from A genomes of both B rapaand B napus potentially participated in metabolism anddevelopment while those from C genome of B napus werelargely involved in stress resistance
34 TransposonActive or Inactive inHybrid Compared to theB rapa genome there were 25 transposable elements (TE)like genes expressed in P1 and P2 while the 11 (44) genesdisplayed differential expression in F1 (Table 3) The EnSpmtype of transposon and the retrotransposon-like transcriptwere significantly enriched It is likely that the interspecific
hybrids significantly modify the transcription of transposon-like genes which may directly or indirectly influence geneexpression of other genes in the heterotic material
35 Validation of Transcriptome Data by qRT-PCR Toassess the accuracy of RNA-seq data ten differentiallyexpressed unigenes including stress responsive genes sec-ondary metabolism biosynthesis genes and epigenetic mod-ifying genes were selected Three genes belong to under-dominance (UDO) and seven genes are belonging to over-dominance (ODO) types which are represented for heterosisanalysis Primer pairs of 10 unigenes for qRT-PCR weredesigned and listed in Table 4 We tested the similaritybetween differential gene expression identified by transcrip-tome and those identified by qRT-PCR As shown in Figure 7the qRT-PCR revealed that 8 of 10 genes (except Bol022348
International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
International Journal of Genomics 3
50 120583m
Figure 1 Root tip squash showing chromosomes from the F1 hybrid (chromosome number of 29)
(a) (b)
Figure 2 Seed germination (a) and 2-week-old seedlings (b) of P1 P2 and F1 germplasm
and the other parental line) To test for significance of MPHvalues the contrast h minus (P1 + P2)2 was used [21]
25 Quantitative RT-PCR Validation of Differentiation GeneExpression RNA-seq datawere further validated using quan-titative real-time PCR analysis (qRT-PCR) for a selectednumber of genes using gene-specific primer sets The plantleaves and growth condition for total RNA extraction usedfor qRT-PCR were the same as these for RNA-seq experi-ments Three independent RNA extractions per sample wereperformed for biological replicates The reverse transcribedinto cDNA using PrimeScritH RT reagent kit with gDNAEraser (Takara Dalian China) Primer pairs of 10 uni-genes were designed using Invitrogenrsquos (Carlsbad CA USA)OligoPerfect Designer software [22] Specificity of the primersets and their product length was verified by agarose gelelectrophoresis The qRT-PCR reaction mixture consisted ofthe SYBR Premix EX-Taq II Kit (TakaRa Japan) on iCycleriQ (BIO-RAD) for three repeats of each sample The thermalcycling conditions were as follows 95 uC 2min and 40 cyclesat 95∘C for 10 s for denaturation and 65∘C 20 s and 72∘C 30 sfor annealing and extension The expression of TIPS-41 withprimer sequences 51015840-TGAAGAGCAGATTGATTTGGCT-31015840and 51015840-ACACTCCATTGTC AGCCAGTT-31015840 was used asan internal control for normalization to compare the geneexpression level between the accessions The relative levels ofgene expression were calculated using the 2minusΔΔCt method
3 Results
31 Sample Preparation Sequencing and Data Filtering Thehybridization of B napus cv CWH-2 (P1) and B rapa ssp
chinensis cv Qianjin (P2) was made in 2010-2011 oilseedgrowing season Root-tip chromosome assessments of F1showed that all F1 plants had a chromosome number of29 (Figure 1) suggesting that they were hybrids The seedsrsquosizes of F1 hybrids were intermediate to those of the parentsbut the seed color resembled that of B napus (Figure 2(a))Root growth of the F1 over the first 24 h was similar toP1 Leaf morphology of the F1 was more similar to B rapathan to B napus (Figure 2) In 4-week-old seedlings theF1 plant displayed stronger roots than either of the parents(Figure 2(b)) while adult F1 plants displayed the similarheading time to B napus
To identify transcripts of P1 P2 and F1 three cDNAlibraries were constructed from their leaves and sequenced byIllumina paired-end sequencing Sequence data from each ofthe libraries consisted of 60Gb from B rapa (AA genome)52 Gb from B napus (AACC genome) and 67Gb from thehybrid (AAC genome) High-quality cleaned raw reads fromall of those data were aligned to reference sequences In total67951208 reads and 46207184 reads were mapped in theB rapa and B oleracea reference genomes respectively Inthe B rapa genome 38522 unigenes were assembled witha minimum scaffold size of 200 bp and a total length of51995312 bp with an average length of 1349 bp (Figure 3(a))The B oleracea genome had a total of 42009 unigeneswith a minimum scaffold size of 300 bp a total length of51691329 bp and an average length of 1230 bp (Figure 3(b))
32 Heterosis of Differential Gene Expression in B rapa Bnapus and Their Hybrid The numbers of expressed genein each genome varied slightly There were 29579 (718)27939 (679) and 29447 (717) from B rapa B napus
4 International Journal of Genomics
0
500
1000
1500
2000
2500
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(a)
0500
1000150020002500300035004000
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(b)
Figure 3 The size distribution of transcriptomic unigenes assembled by the B rapa (a) and B oleracea (b)
and the hybrid genome respectively The unique mappingrates were similar among three Brassica samples suggestingthat their genomes were quite alike Some genes had expres-sion that was germplasm specific The B rapa genome hadapproximately about 6 (1639) of genes that were uniquewhilst B napus and the hybrid had only 2 (420) and3 (951) unique sequences respectively There were 25985genes expressed in all three lines A total of 489 genes wereexpressed in both B rapa and B napus but silenced in thehybrid Gene expression profiles of the three lines revealedthat the number of expressed genes did not show obviousdifference in the three sets of material The mode of geneaction for the differentially expressed genes (DEGs) wasanalyzed About 2700 genes (1840 of 6816) exhibited anexpression pattern that was not distinguishable from additiv-ity while the other 7256 (4946 of 6816) of genes showednonadditive expression patterns (Table 1) The nonadditivenumber ofDEGs from the crosswas further classified into fivedistinct classes high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) underdominance(UDO) and partial-dominanceThere were 7 of transcripts(545) that were absent in their parents indicating that therewas transcriptional activation of new genes in hybrid F1There was also transcriptional silencing with 32 (218) ofparentally expressed gene not being expressed in the hybrid
There were 2894 unigenes that mapped to the A genome2107 that mapped to the C genome (Figure 4) and 593that were not assigned to either genome Chromosomaldistribution of DEG numbers in hybrids showed that low-parent domiance (LPD) genes were significantly accumulatedon chromosomes A06 and C07 (119875 lt 0001) while high-parent dominance (HPD) genes were on chromosomes A09and C08 (119875 lt 001) Gene expression variation causedby interspecific hybridization was not randomly distributedalong the chromosomesTherefore differentB napus-B rapachromosome introgression lines can be developed to the fur-ther study of heterosis pertaining to specific chromosomes
33 Contribution of A and C Subgenomes to Gene DifferentialExpression in the Hybrid There were 1386 significant differ-entially expressed genes between P1 and F1 samples 858genes had increased expression in the F1 and 528 genes
Table 1 Summary of dominance patterns of differentially expressedgenes
DGs Number PercentageAdditivity 1840 27Nonadditivity 4946 7256Others 30 044High-parent dominance(HPD) 353 714
Low-parent dominance(LPD) 191 386
Overdominance (ODO) 593 1199Underdominance (UDO) 193 39Positive partial-dominance(PPD) 1635 3306
Negativepartial-dominance (NPD) 1981 405
Total 6816Additivity F1 asymp 12(P1 + P2) nonadditivity F1 gt 12(P1 + P2) or F1 lt 12(P1 +P2) High-parent dominance (HPD) F1asymp P1gt P2 or F1asymp P2lt P1 low-parentdominance (LPD) F1 asymp P1 lt P2 or F1 asymp P2 lt P1 overdominance (ODO) F1gt P1 and F1 gt P2 underdominance (UDO) F1 lt P1 and F1 lt P2
had decreased expression There were 5077 significant dif-ferentially expressed genes between P2 and F1 samples with3241 genes having increased expression in the F1 and 1836genes being decreased in expression (Figure 5) Changesto expression from P2 were approximately three times thenumber of genes compared to from P1 There were moregenes with increased expression in the F1 (4098) than withdecreased expression (2464) when compared to expressionlevels in the respective parents We performed gene ontology(GO) enrichment analysis to test the functional categoriesfor DEGs among P1 P2 and F1 (Figure 6) by AgriGO onlinetool [23] and R statistical software [20] There were 1012 and 18 functional categories of these transcripts thatbelonged to the cell component (CC) molecular function(MF) and biological process (BP) respectively The GOanalysis showed that the composition of functional pathwaysassociated with genes showing differential expression was
International Journal of Genomics 5
0
20
40
60
80
100
120
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10
LPDHPD
UDOODO
(a)
0
20
40
60
80
100
120
140
C1 C2 C3 C4 C5 C6 C7 C8 C9
LPDHPD
UDOODO
(b)
Figure 4 Chromosomal distribution of nonadditive differentially expressed genes with high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) and underdominance (UDO) on the A genome (a) and C genome (b)
P1_versus_F1F1_versus_P2
P1_versus_P2
694
310
3765
308
971
402
366
(a)
3000
2000
1000
0
858
528
1836
3241
2024
3422
RegulationUpDown
F1_versus_P1 F1_versus_P2 P1_versus_P2
(b)
Figure 5 Total numbers of differentially expressed genes between P1 P2 and F1 by Venn diagram analyses (a) and statistics of up- ordownregulated genes (b)
similar when comparing changes from either P1 or P2 withF1
The biological process category was further subdividedinto functional classes in those DEGs with high-parent
dominance (HPD 119875 lt 05) based on gene ontology Table 2highlights the 18 gene ontology classes that were differentiallyexpressed in the F1 Combined with the results in Figure 6we found that the GOs such as regulation of transcription
6 International Journal of Genomics
Response to chitinRegulation of plant-type hypersensitive responseResponse to woundingRespiratory burst involved in defense responseProtein targeting to membraneEthylene biosynthetic processJasmonic acid mediated signaling pathwayDefense response to fungusRegulation of hydrogen peroxide metabolic processSystemic acquired resistance salicylic acid mediated signaling pathwayAbscisic acid-activated signaling pathwayMAPK cascadeIntracellular signal transductionIndoleacetic acid biosynthetic processResponse to jasmonic acidResponse to ethyleneHyperosmotic salinity responseNegative regulation of defense responsePositive regulation of flavonoid biosynthetic processResponse to water deprivationDefense response to bacteriumGlucosinolate catabolic processResponse to mechanical stimulusResponse to endoplasmic reticulum stressDetection of biotic stimulusResponse to herbivoreResponse to bacteriumResponse to cyclopentenoneJasmonic acid biosynthetic processResponse to fungus
1
08
06
04
02
005
0
F1_v
ersu
s_P2
P1_v
ersu
s_P2
P1_v
ersu
s_F1
Figure 6 The heat map from GO enrichment analysis of DEGs between the P1 P2 and F1 combinations Color scale represents 119875 values ofenrichment test
metabolic process defense response multicellular organ-ismal development and transporters were enriched in Agenomes transcripts The genes responsible for stress werestriking enriched in the differential expressed genes fromC genome It possibly implied that the hybrid expressedprotein-coding genes from A genomes of both B rapaand B napus potentially participated in metabolism anddevelopment while those from C genome of B napus werelargely involved in stress resistance
34 TransposonActive or Inactive inHybrid Compared to theB rapa genome there were 25 transposable elements (TE)like genes expressed in P1 and P2 while the 11 (44) genesdisplayed differential expression in F1 (Table 3) The EnSpmtype of transposon and the retrotransposon-like transcriptwere significantly enriched It is likely that the interspecific
hybrids significantly modify the transcription of transposon-like genes which may directly or indirectly influence geneexpression of other genes in the heterotic material
35 Validation of Transcriptome Data by qRT-PCR Toassess the accuracy of RNA-seq data ten differentiallyexpressed unigenes including stress responsive genes sec-ondary metabolism biosynthesis genes and epigenetic mod-ifying genes were selected Three genes belong to under-dominance (UDO) and seven genes are belonging to over-dominance (ODO) types which are represented for heterosisanalysis Primer pairs of 10 unigenes for qRT-PCR weredesigned and listed in Table 4 We tested the similaritybetween differential gene expression identified by transcrip-tome and those identified by qRT-PCR As shown in Figure 7the qRT-PCR revealed that 8 of 10 genes (except Bol022348
International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 International Journal of Genomics
0
500
1000
1500
2000
2500
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(a)
0500
1000150020002500300035004000
3000+
2900sim3000
2800sim2900
2700sim2800
2600sim2700
2500sim2600
2400sim2500
2300sim2400
2200sim2300
2100sim2200
2000sim2100
1900sim2000
1800sim1900
1700sim1800
1600sim1700
1500sim1600
1400sim1500
1300sim1400
1200sim1300
1100sim1200
1000sim1100
900sim1000
800sim900
700sim800
600sim700
500sim600
400sim500
300sim400
(b)
Figure 3 The size distribution of transcriptomic unigenes assembled by the B rapa (a) and B oleracea (b)
and the hybrid genome respectively The unique mappingrates were similar among three Brassica samples suggestingthat their genomes were quite alike Some genes had expres-sion that was germplasm specific The B rapa genome hadapproximately about 6 (1639) of genes that were uniquewhilst B napus and the hybrid had only 2 (420) and3 (951) unique sequences respectively There were 25985genes expressed in all three lines A total of 489 genes wereexpressed in both B rapa and B napus but silenced in thehybrid Gene expression profiles of the three lines revealedthat the number of expressed genes did not show obviousdifference in the three sets of material The mode of geneaction for the differentially expressed genes (DEGs) wasanalyzed About 2700 genes (1840 of 6816) exhibited anexpression pattern that was not distinguishable from additiv-ity while the other 7256 (4946 of 6816) of genes showednonadditive expression patterns (Table 1) The nonadditivenumber ofDEGs from the crosswas further classified into fivedistinct classes high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) underdominance(UDO) and partial-dominanceThere were 7 of transcripts(545) that were absent in their parents indicating that therewas transcriptional activation of new genes in hybrid F1There was also transcriptional silencing with 32 (218) ofparentally expressed gene not being expressed in the hybrid
There were 2894 unigenes that mapped to the A genome2107 that mapped to the C genome (Figure 4) and 593that were not assigned to either genome Chromosomaldistribution of DEG numbers in hybrids showed that low-parent domiance (LPD) genes were significantly accumulatedon chromosomes A06 and C07 (119875 lt 0001) while high-parent dominance (HPD) genes were on chromosomes A09and C08 (119875 lt 001) Gene expression variation causedby interspecific hybridization was not randomly distributedalong the chromosomesTherefore differentB napus-B rapachromosome introgression lines can be developed to the fur-ther study of heterosis pertaining to specific chromosomes
33 Contribution of A and C Subgenomes to Gene DifferentialExpression in the Hybrid There were 1386 significant differ-entially expressed genes between P1 and F1 samples 858genes had increased expression in the F1 and 528 genes
Table 1 Summary of dominance patterns of differentially expressedgenes
DGs Number PercentageAdditivity 1840 27Nonadditivity 4946 7256Others 30 044High-parent dominance(HPD) 353 714
Low-parent dominance(LPD) 191 386
Overdominance (ODO) 593 1199Underdominance (UDO) 193 39Positive partial-dominance(PPD) 1635 3306
Negativepartial-dominance (NPD) 1981 405
Total 6816Additivity F1 asymp 12(P1 + P2) nonadditivity F1 gt 12(P1 + P2) or F1 lt 12(P1 +P2) High-parent dominance (HPD) F1asymp P1gt P2 or F1asymp P2lt P1 low-parentdominance (LPD) F1 asymp P1 lt P2 or F1 asymp P2 lt P1 overdominance (ODO) F1gt P1 and F1 gt P2 underdominance (UDO) F1 lt P1 and F1 lt P2
had decreased expression There were 5077 significant dif-ferentially expressed genes between P2 and F1 samples with3241 genes having increased expression in the F1 and 1836genes being decreased in expression (Figure 5) Changesto expression from P2 were approximately three times thenumber of genes compared to from P1 There were moregenes with increased expression in the F1 (4098) than withdecreased expression (2464) when compared to expressionlevels in the respective parents We performed gene ontology(GO) enrichment analysis to test the functional categoriesfor DEGs among P1 P2 and F1 (Figure 6) by AgriGO onlinetool [23] and R statistical software [20] There were 1012 and 18 functional categories of these transcripts thatbelonged to the cell component (CC) molecular function(MF) and biological process (BP) respectively The GOanalysis showed that the composition of functional pathwaysassociated with genes showing differential expression was
International Journal of Genomics 5
0
20
40
60
80
100
120
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10
LPDHPD
UDOODO
(a)
0
20
40
60
80
100
120
140
C1 C2 C3 C4 C5 C6 C7 C8 C9
LPDHPD
UDOODO
(b)
Figure 4 Chromosomal distribution of nonadditive differentially expressed genes with high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) and underdominance (UDO) on the A genome (a) and C genome (b)
P1_versus_F1F1_versus_P2
P1_versus_P2
694
310
3765
308
971
402
366
(a)
3000
2000
1000
0
858
528
1836
3241
2024
3422
RegulationUpDown
F1_versus_P1 F1_versus_P2 P1_versus_P2
(b)
Figure 5 Total numbers of differentially expressed genes between P1 P2 and F1 by Venn diagram analyses (a) and statistics of up- ordownregulated genes (b)
similar when comparing changes from either P1 or P2 withF1
The biological process category was further subdividedinto functional classes in those DEGs with high-parent
dominance (HPD 119875 lt 05) based on gene ontology Table 2highlights the 18 gene ontology classes that were differentiallyexpressed in the F1 Combined with the results in Figure 6we found that the GOs such as regulation of transcription
6 International Journal of Genomics
Response to chitinRegulation of plant-type hypersensitive responseResponse to woundingRespiratory burst involved in defense responseProtein targeting to membraneEthylene biosynthetic processJasmonic acid mediated signaling pathwayDefense response to fungusRegulation of hydrogen peroxide metabolic processSystemic acquired resistance salicylic acid mediated signaling pathwayAbscisic acid-activated signaling pathwayMAPK cascadeIntracellular signal transductionIndoleacetic acid biosynthetic processResponse to jasmonic acidResponse to ethyleneHyperosmotic salinity responseNegative regulation of defense responsePositive regulation of flavonoid biosynthetic processResponse to water deprivationDefense response to bacteriumGlucosinolate catabolic processResponse to mechanical stimulusResponse to endoplasmic reticulum stressDetection of biotic stimulusResponse to herbivoreResponse to bacteriumResponse to cyclopentenoneJasmonic acid biosynthetic processResponse to fungus
1
08
06
04
02
005
0
F1_v
ersu
s_P2
P1_v
ersu
s_P2
P1_v
ersu
s_F1
Figure 6 The heat map from GO enrichment analysis of DEGs between the P1 P2 and F1 combinations Color scale represents 119875 values ofenrichment test
metabolic process defense response multicellular organ-ismal development and transporters were enriched in Agenomes transcripts The genes responsible for stress werestriking enriched in the differential expressed genes fromC genome It possibly implied that the hybrid expressedprotein-coding genes from A genomes of both B rapaand B napus potentially participated in metabolism anddevelopment while those from C genome of B napus werelargely involved in stress resistance
34 TransposonActive or Inactive inHybrid Compared to theB rapa genome there were 25 transposable elements (TE)like genes expressed in P1 and P2 while the 11 (44) genesdisplayed differential expression in F1 (Table 3) The EnSpmtype of transposon and the retrotransposon-like transcriptwere significantly enriched It is likely that the interspecific
hybrids significantly modify the transcription of transposon-like genes which may directly or indirectly influence geneexpression of other genes in the heterotic material
35 Validation of Transcriptome Data by qRT-PCR Toassess the accuracy of RNA-seq data ten differentiallyexpressed unigenes including stress responsive genes sec-ondary metabolism biosynthesis genes and epigenetic mod-ifying genes were selected Three genes belong to under-dominance (UDO) and seven genes are belonging to over-dominance (ODO) types which are represented for heterosisanalysis Primer pairs of 10 unigenes for qRT-PCR weredesigned and listed in Table 4 We tested the similaritybetween differential gene expression identified by transcrip-tome and those identified by qRT-PCR As shown in Figure 7the qRT-PCR revealed that 8 of 10 genes (except Bol022348
International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
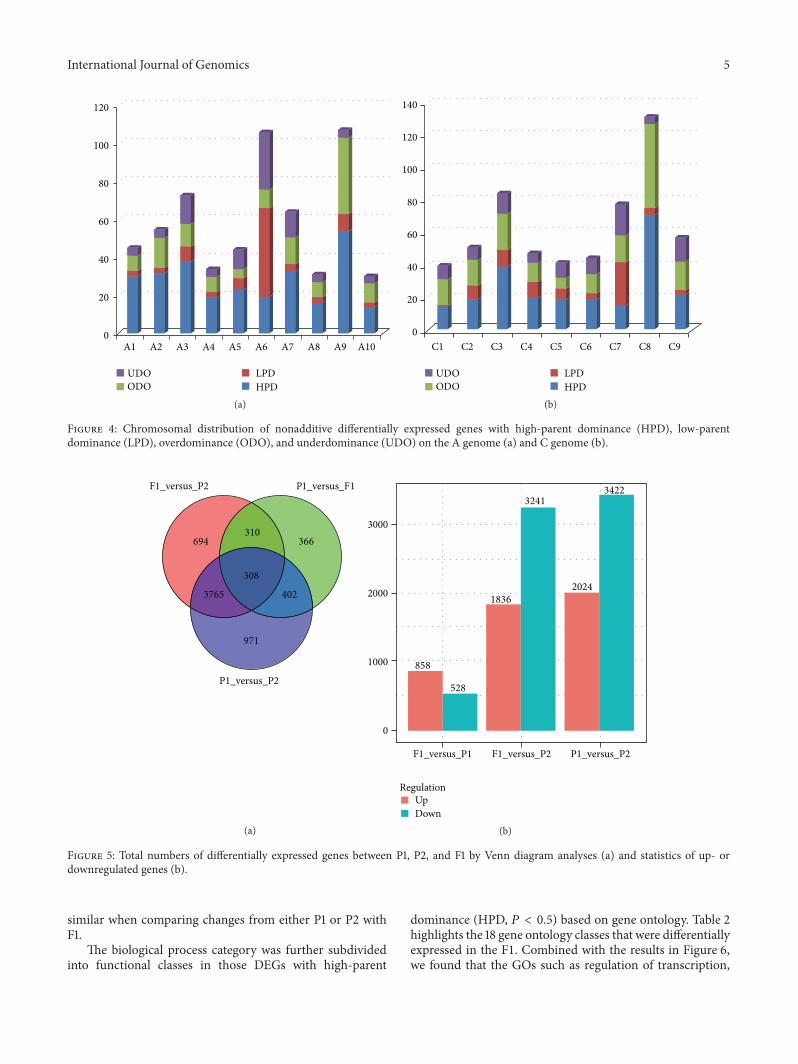
International Journal of Genomics 5
0
20
40
60
80
100
120
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10
LPDHPD
UDOODO
(a)
0
20
40
60
80
100
120
140
C1 C2 C3 C4 C5 C6 C7 C8 C9
LPDHPD
UDOODO
(b)
Figure 4 Chromosomal distribution of nonadditive differentially expressed genes with high-parent dominance (HPD) low-parentdominance (LPD) overdominance (ODO) and underdominance (UDO) on the A genome (a) and C genome (b)
P1_versus_F1F1_versus_P2
P1_versus_P2
694
310
3765
308
971
402
366
(a)
3000
2000
1000
0
858
528
1836
3241
2024
3422
RegulationUpDown
F1_versus_P1 F1_versus_P2 P1_versus_P2
(b)
Figure 5 Total numbers of differentially expressed genes between P1 P2 and F1 by Venn diagram analyses (a) and statistics of up- ordownregulated genes (b)
similar when comparing changes from either P1 or P2 withF1
The biological process category was further subdividedinto functional classes in those DEGs with high-parent
dominance (HPD 119875 lt 05) based on gene ontology Table 2highlights the 18 gene ontology classes that were differentiallyexpressed in the F1 Combined with the results in Figure 6we found that the GOs such as regulation of transcription
6 International Journal of Genomics
Response to chitinRegulation of plant-type hypersensitive responseResponse to woundingRespiratory burst involved in defense responseProtein targeting to membraneEthylene biosynthetic processJasmonic acid mediated signaling pathwayDefense response to fungusRegulation of hydrogen peroxide metabolic processSystemic acquired resistance salicylic acid mediated signaling pathwayAbscisic acid-activated signaling pathwayMAPK cascadeIntracellular signal transductionIndoleacetic acid biosynthetic processResponse to jasmonic acidResponse to ethyleneHyperosmotic salinity responseNegative regulation of defense responsePositive regulation of flavonoid biosynthetic processResponse to water deprivationDefense response to bacteriumGlucosinolate catabolic processResponse to mechanical stimulusResponse to endoplasmic reticulum stressDetection of biotic stimulusResponse to herbivoreResponse to bacteriumResponse to cyclopentenoneJasmonic acid biosynthetic processResponse to fungus
1
08
06
04
02
005
0
F1_v
ersu
s_P2
P1_v
ersu
s_P2
P1_v
ersu
s_F1
Figure 6 The heat map from GO enrichment analysis of DEGs between the P1 P2 and F1 combinations Color scale represents 119875 values ofenrichment test
metabolic process defense response multicellular organ-ismal development and transporters were enriched in Agenomes transcripts The genes responsible for stress werestriking enriched in the differential expressed genes fromC genome It possibly implied that the hybrid expressedprotein-coding genes from A genomes of both B rapaand B napus potentially participated in metabolism anddevelopment while those from C genome of B napus werelargely involved in stress resistance
34 TransposonActive or Inactive inHybrid Compared to theB rapa genome there were 25 transposable elements (TE)like genes expressed in P1 and P2 while the 11 (44) genesdisplayed differential expression in F1 (Table 3) The EnSpmtype of transposon and the retrotransposon-like transcriptwere significantly enriched It is likely that the interspecific
hybrids significantly modify the transcription of transposon-like genes which may directly or indirectly influence geneexpression of other genes in the heterotic material
35 Validation of Transcriptome Data by qRT-PCR Toassess the accuracy of RNA-seq data ten differentiallyexpressed unigenes including stress responsive genes sec-ondary metabolism biosynthesis genes and epigenetic mod-ifying genes were selected Three genes belong to under-dominance (UDO) and seven genes are belonging to over-dominance (ODO) types which are represented for heterosisanalysis Primer pairs of 10 unigenes for qRT-PCR weredesigned and listed in Table 4 We tested the similaritybetween differential gene expression identified by transcrip-tome and those identified by qRT-PCR As shown in Figure 7the qRT-PCR revealed that 8 of 10 genes (except Bol022348
International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 International Journal of Genomics
Response to chitinRegulation of plant-type hypersensitive responseResponse to woundingRespiratory burst involved in defense responseProtein targeting to membraneEthylene biosynthetic processJasmonic acid mediated signaling pathwayDefense response to fungusRegulation of hydrogen peroxide metabolic processSystemic acquired resistance salicylic acid mediated signaling pathwayAbscisic acid-activated signaling pathwayMAPK cascadeIntracellular signal transductionIndoleacetic acid biosynthetic processResponse to jasmonic acidResponse to ethyleneHyperosmotic salinity responseNegative regulation of defense responsePositive regulation of flavonoid biosynthetic processResponse to water deprivationDefense response to bacteriumGlucosinolate catabolic processResponse to mechanical stimulusResponse to endoplasmic reticulum stressDetection of biotic stimulusResponse to herbivoreResponse to bacteriumResponse to cyclopentenoneJasmonic acid biosynthetic processResponse to fungus
1
08
06
04
02
005
0
F1_v
ersu
s_P2
P1_v
ersu
s_P2
P1_v
ersu
s_F1
Figure 6 The heat map from GO enrichment analysis of DEGs between the P1 P2 and F1 combinations Color scale represents 119875 values ofenrichment test
metabolic process defense response multicellular organ-ismal development and transporters were enriched in Agenomes transcripts The genes responsible for stress werestriking enriched in the differential expressed genes fromC genome It possibly implied that the hybrid expressedprotein-coding genes from A genomes of both B rapaand B napus potentially participated in metabolism anddevelopment while those from C genome of B napus werelargely involved in stress resistance
34 TransposonActive or Inactive inHybrid Compared to theB rapa genome there were 25 transposable elements (TE)like genes expressed in P1 and P2 while the 11 (44) genesdisplayed differential expression in F1 (Table 3) The EnSpmtype of transposon and the retrotransposon-like transcriptwere significantly enriched It is likely that the interspecific
hybrids significantly modify the transcription of transposon-like genes which may directly or indirectly influence geneexpression of other genes in the heterotic material
35 Validation of Transcriptome Data by qRT-PCR Toassess the accuracy of RNA-seq data ten differentiallyexpressed unigenes including stress responsive genes sec-ondary metabolism biosynthesis genes and epigenetic mod-ifying genes were selected Three genes belong to under-dominance (UDO) and seven genes are belonging to over-dominance (ODO) types which are represented for heterosisanalysis Primer pairs of 10 unigenes for qRT-PCR weredesigned and listed in Table 4 We tested the similaritybetween differential gene expression identified by transcrip-tome and those identified by qRT-PCR As shown in Figure 7the qRT-PCR revealed that 8 of 10 genes (except Bol022348
International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 7
Table 2 Significant gene ontology terms of high-parent dominance(HPD) in the biological process category from A and C genomes
Gene ID Gene O termsBra028791 Regulation of transcription (GO0006355)Bra005111 Metabolic process (GO0008152)Bra032185 Defense response (GO0002679)Bra010724 Multicellular organismal development (GO0007275)Bra007111 Metabolic process (GO0010408)Bra031515 Single-organism transport (GO0044765)Bra031678 Transport (GO0006810)Bra027171 Transmembrane transport (GO0006855)Bra024033 Peroxidase activity (GO0004601)Bra002550 Chromosome segregation (GO0007059)
Bol015697 Response to molecule of bacterial origin(GO0002237)
Bol032712 Defense response to insect (GO0002213)Bol014351 Defense response to bacterium (GO0042742)Bol045822 Defense response (GO0002679)Bol034224 Response to wounding (GO0009611)Bol038369 Single-organism process (GO0044699)Bol045822 Regulation of transcription (GO0006355)
Bol015697 Response to molecule of bacterial origin(GO0002237)
and Bol026880) that showed the differential gene expressionlevel agreed well with the expression patterns of DGE dataHence the qRT-PCR results showed general agreement withtheir transcript abundance changes determined by RNA-seqwhich suggested the reliability of the transcriptomic profilingdata among the three samples
4 Discussion
Interspecific hybridization is an important approach toobtaining and utilizing novel agronomic traits from relatedspecies for crop improvement Previous researches revealedstrong heterosis between B napus and B rapa on biomassand seed yield [5ndash7] Recently Li et al [24] reported thatB napus-like individuals in the F3 and F4 generations frominterspecific hybridization between B napus and B oleraceashowed diverse genetic variation relative to current B napusand strong heterotic potential We selected a number ofbackcrossed offspring derived from the F1 by being crossedwith P1 (B napus) and new traits such as wrinkle leaves sixsepals which different from their parents were observed inthe progenies In particular we identified several lines whichdisplayed high yield up to 4500 kg per hectare in BC1F2population It suggested that the novel lines from interspecifichybridization between B napus and B rapa with high yieldgood quality can be potentially useful for oilseeds productionin the near future
Intraspecific hybrids of Arabidopsis thaliana [25 26]rice [27ndash29] and maize [30 31] revealed that there areboth additive and nonadditive effects of gene expression
in hybrids and heterosis is strongly influenced by geneticdifferences between two parents A transcriptome approachhas been utilized to investigate the heterosis of intraspecifichybrids ofA thaliana ricemaize [29 30]Our research of thetranscriptomes from B napus and B rapa demonstrated thegenetic effect of interspecies heterosis at the gene expressionlevel More genes specific to certain pathways in furtherexperiments are needed to fully validate the heterotic asso-ciation between the gene expression patterns and the targetagronomic traits In the present study we found that bothadditive and dominance effects contributed to interspeciesheterosis between B napus and B rapa Analysis of thenumber of DEGs in hybrid and parental germplasm revealedthat the nonadditive effect contributed more than the addi-tive effect Previous research reveals interspecies heterosisbetweenB napus andB rapa through genetic analysis but themain focus was on phenotypic variation [6 32] Our studyprovided evidence of differential gene expression and shednew insights on the molecular mechanism for this heterosisin the interspecific hybrids
Brassica species constitute an elite system for investi-gating the changes in genomic structure and functionaldivergence of duplicate genes in the process of hybridizationand polyploidization among the diploid species and theiramphidiploids [32 33] Subgenomes in Brassica species aredesignated with superscripts with An and Ar representing Agenomes from B napus and B rapa respectively [33] Wehave shown intersubgenomic heterosis between Ar and An
genomes supporting work by others that show that suchheterosis can be useful in creating genetic diversity andbe used in breeding Studies indicated that the midparentheterosis contributes intersubgenomic heterosis [8 34 35]Therefore the utilization of intersubgenomic heterosis canbe creating genetic diversity and breeding In the presentstudy we found that the nonadditive DEGs from the Agenome in the hybrids mainly participated in metabolismand development while those from the C genome werelargely involved in stress resistance It is likely that theneofunctionalization of subgenome contributes the heterosisor the genomic adaptation in hybrid Recently Li et al[36] investigated the total homoeolog expression level ofhexaploid wheat (AABBDD) and compared it to its ancestralparents T turgidum (AABB) and Ae tauschii (DD) findingthat genes expressed from the AABB potentially participatedin development and those from Ae tauschii (DD) were morelikely involved in adaptation Differential contributions fromthe parental sources may also be occurring in the Brassicahybrids and this may hint at the molecular mechanismsbehind heterosis Transposable elements (TEs) have majoreffects on different plant genomes and can significantlycontribute differences in genome size of over 1000-fold [37]Recent sequencing of the B rapa and B oleracea genomesrevealed that the amplification of TEs is one of the mainfactors inducing the difference in genome size [37] Theoverall TEs in either Brassica genomes are expressed at verylow levels and the expression levels of different TE categoriesand families vary among different organs [38] Recently thevariations in TE-associated sequences during the processof allopolyploidisation were detected between Brassica rapa
8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 International Journal of Genomics
Table 3 The differential expressed transposon-like genes in hybrid F1
Gene ID Regulation Chromosome Similar to transposonBrapanewgene1068 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene871 Inactive A07 Putative transposon protein (Arabidopsis thaliana)Brapanewgene1142 Inactive A09 Retrotransposon-like protein (Arabidopsis thaliana)Brapanewgene957 Inactive A08 Retrotransposon (Arabidopsis thaliana)Bra040302 Up A08 Putative transposon protein (Arabidopsis thaliana)Brapanewgene307 Up A03 EnSpm transposon protein (Arabidopsis thaliana)Brapanewgene726 Up A06 EnSpm-like transposon protein (Arabidopsis thaliana)Bra034711 Active A05 hAT transposon superfamily protein (Arabidopsis thaliana)Bra027867 Active A09 Pol polyprotein transposon element Bs1 (Zea mays)Brapanewgene8 Active A01 Tam1 transposon protein (Arabidopsis thaliana)Bra035011 Active A07 Putative transposon protein (Arabidopsis thaliana)Bol019342 Inactive C09 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol041429 Up C07 Putative Tam3-like transposon protein (Zea mays)Bol035479 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol005562 Down C03 EnSpm-related transposon protein (Brassica oleracea)Bol019765 Down C09 EnSpm-related transposon protein (Brassica oleracea)Bol006373 Down C07 Transposon protein-like (Arabidopsis thaliana)Bol035958 Up C02 Transposon-like ORF (Brassica oleracea)Bol030703 Active C03 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol030374 Active C09 EnSpm-related transposon protein (Brassica oleracea)Bol004880 Active C04 Retrotransposon Tto1 DNA (Nicotiana tabacum)Bol043208 Active C07 Similar to retrotransposon (Arabidopsis thaliana)
Table 4 The qPCR primers for selected genes
Unigenes Primers Biological process NonadditivityBra007087 F TGCAGCGCTTGATTTACCT Oxidation-reduction
process HPDBra007087 R GCAAACTCCAGAGCTATGTBra020376 F AGGTCATTCTGGTGAGCCACA Chaperones HPDBra020376 R TGGAGACTTTGGAAGGATACTBra031210 F ACGAGGCTCAGTCTCGTGGT Response to water
deprivation ODOBra031210 R TCCGCTGCGGTATCCACCABra031666 F TGCCAAGGACAACAACTTGGACT Lignin biosynthetic
process HPDBra031666 R AGTTGGTTGTAGGACTGGTCCABol016209 F TAACCTACCAGAAGCACGGT Response to abscisic
acid ODOBol016209 R ACAACTTCAACGGTGCACGACTBol022348 F AGGTACCTTACGAGTCTCGT Response to karrikin HPDBol022348 R CACGAACCTGATAGAAGCTCGTBol026880 F TGTAAGGCTACGAAGGGACAT Defense response to
fungus HPDBol026880 R TCCAACATATCCATATGTCCGTBol026959 F TAGCTCTTCCTCTTCAAGCGAT Defense response to
fungus ODOBol026959 R TCCTTCTTCCTCTTCTCACCABol041012 F TGACACATTGTGGATGGAACT Cellular hyperosmotic
response ODOBol041012 R GGCATACCAATCATTGGAACTBol044348 F TACTACTGGACCTTTGGTGCT Calcium ion
transport HPDBol044348 R GATCTTCATCTGAAGGTCACT
International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 9
20
15
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Bra007087
P1 P2 F1 P1 P2 F1
P1 P2 F1 P1 P2 F1
P1 P2 F1
15
P1 P2 F1
P1 P2 F1
P1 P2 F1
10
05
00
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bra020376
Bra0316663
2
1
0
Bol022348
140
120
100
80
60
10
8
6
4
20
15
10
05
00
Bra0312108
6
4
2
0
Bol026880
6
4
2
0
Bol026959
140
120
100
60
58
56
54
52
50
8
6
4
2
0
Bol041012
Figure 7 Continued
10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 International Journal of Genomics
P1 P2 F1 P1 P2 F1
Relat
ive e
xpre
ssio
n
Relat
ive e
xpre
ssio
n
15
10
05
00
Bol0443485
4
3
2
1
0
Bol016209
Figure 7 Genes with expression levels validated by qRT-PCR
(AA) andB oleracea (CC) aswell as in successive generationsof self-pollinated progeny [39] Our study revealed thatthe expression activities of TE categories or families wereclearly modified in the hybrids between the two Brassicaspecies and different types of TEs showed different patternsof variation during the process of hybridization Thereforethe association of activation and inactivation of TEs andof TE related genes expression in relation to heterotic geneexpression needs to be further investigated
5 Conclusions
Development of interspecific hybrids has been widelyexploited for the heterosis breeding of Brassica crops Theinterspecific hybrid between Brassica napus and B rapadisplayed obvious heterosis In the present study the com-parative transcriptome analysis for the parents B napus andB rapa and its hybrid F1 was sequenced using the platform ofIlluminaSolexa with de novo assemblageThe nonredundantunigenes were identified using B rapa (A genome) and Boleracea (C genome) as reference genomesThenonadditivelydifferentially expressed genes were potentially related tothe heterosis The results indicated that the differentiallyexpressed genes in hybrids from the A genome mainlyparticipated in metabolism and development while thosefrom the C genome were largely involved in stress resistanceThe coexistence of multiple gene actions and transposableelement-associated genes regulations in the hybrid wereobserved The present study could be helpful for the betterunderstanding of the mechanisms of heterosis and a list ofcandidate genes will be used for future Brassica breeding
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank the Earmarked Fund for Modern Agro-industry Technology Research System of China the National
High Technology Research and Development Program ldquo863rdquoof China (2011AA10A104) and National Science and Tech-nology Supporting Plan of China (2011BAD35B04) TheMinistry of agriculture experimental observation of theupper reaches of the Yangtze River Oil Crop Science Sta-tion (09203020) and Sichuan Crop Breeding Community(2011NZ0098-5 2011NZ0098-16 and 2011JYGC04-013) forthe financial support
References
[1] J A Foley N Ramankutty K A Brauman et al ldquoSolutions fora cultivated planetrdquoNature vol 478 no 7369 pp 337ndash342 2011
[2] U Nagaharu ldquoGenome analysis in Brassica with special refer-ence to the experimental formation of B napus and peculiarmode of fertilizationrdquo Japanese Journal of Botany vol 7 pp 389ndash452 1935
[3] A Abbadi andG Leckband ldquoRapeseed breeding for oil contentquality and sustainabilityrdquo European Journal of Lipid Scienceand Technology vol 113 no 10 pp 1198ndash1206 2011
[4] T Jesske B Olberg A Schierholt and H C Becker ldquoResynthe-sized lines from domesticated and wild Brassica taxa and theirhybrids with B napus L genetic diversity and hybrid yieldrdquoTheoretical and Applied Genetics vol 126 no 4 pp 1053ndash10652013
[5] H L Liu Genetics and Breeding in Rapeseed Chinese Agricul-tural Universities Press Beijing China 2000
[6] R Liu W Qian and J Meng ldquoAssociation of RFLP markersand biomass heterosis in trigenomic hybrids of oilseed rape(Brassica napus times B campestris)rdquo Theoretical and AppliedGenetics vol 105 no 6-7 pp 1050ndash1057 2002
[7] D Fu M Xiao A Hayward et al ldquoWhat is crop heterosis newinsights into an old topicrdquo Journal of Applied Genetics vol 56no 1 pp 1ndash13 2014
[8] J A Birchler D L Auger and N C Riddle ldquoIn search of themolecular basis of heterosisrdquo Plant Cell vol 15 no 10 pp 2236ndash2239 2003
[9] Z J Chen ldquoGenomic and epigenetic insights into themolecularbases of heterosisrdquo Nature Reviews Genetics vol 14 no 7 pp471ndash482 2013
[10] N R Hofmann ldquoA global view of hybrid vigor DNA methyla-tion small RNAs and gene expressionrdquo Plant Cell vol 24 no3 article 841 2012
International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 11
[11] R Zhai Y Feng H Wang et al ldquoTranscriptome analysis of riceroot heterosis by RNA-Seqrdquo BMC Genomics vol 14 article 192013
[12] S Yu F Zhang X Wang et al ldquoGenetic diversity and marker-trait associations in a collection of Pak-choi (Brassica rapa Lssp chinensis Makino) Accessionsrdquo Genes and Genomics vol32 no 5 pp 419ndash428 2014
[13] X L Zhang and L G Zhang ldquoMolecular cloning and expres-sion of the male sterility-related CtYABBY1 gene in floweringChinese cabbage (Brassica campestris L ssp chinensis varparachinensis)rdquo Genetics and Molecular Research vol 13 no 2pp 4336ndash4347 2014
[14] N Wang F Li B Chen et al ldquoGenome-wide investigation ofgenetic changes during modern breeding of Brassica napusrdquoTheoretical and Applied Genetics vol 127 no 8 pp 1817ndash18292014
[15] S Lin H Dong F Zhang et al ldquoBcMF8 a putative ara-binogalactan protein-encoding gene contributes to pollen walldevelopment aperture formation and pollen tube growth inBrassica campestrisrdquo Annals of Botany vol 113 no 5 pp 777ndash788 2014
[16] H Zhan X Zhang G Li et al ldquoMolecular characterization ofa new wheat-Thinopyrum intermedium translocation line withresistance to powdery mildew and stripe rustrdquo InternationalJournal of Molecular Sciences vol 16 no 1 pp 2162ndash2173 2015
[17] R Li C Yu Y Li et al ldquoSOAP2 an improved ultrafast tool forshort read alignmentrdquo Bioinformatics vol 25 no 15 pp 1966ndash1967 2009
[18] A Mortazavi B A Williams K McCue L Schaeffer and BWold ldquoMapping and quantifying mammalian transcriptomesby RNA-Seqrdquo Nature Methods vol 5 no 7 pp 621ndash628 2008
[19] S Audic and J-M Claverie ldquoThe significance of digital geneexpression profilesrdquo Genome Research vol 7 no 10 pp 986ndash995 1997
[20] RC Team R A Language and Environment for StatisticalComputing R Foundation for Statistical Computing ViennaAustria 2013
[21] H Ding C Qin X Luo et al ldquoHeterosis in early maize earinflorescence development a genome-wide transcription anal-ysis for two maize inbred lines and their hybridrdquo InternationalJournal of Molecular Sciences vol 15 no 8 pp 13892ndash139152014
[22] Custom Primers-OligoPerfect Designer Invitrogen CarlsbadCalif USA 2006
[23] Z Du X Zhou Y Ling Z Zhang and Z Su ldquoAgriGO a GOanalysis toolkit for the agricultural communityrdquo Nucleic AcidsResearch vol 38 no 2 pp W64ndashW70 2010
[24] Q Li Q Zhou J Mei et al ldquoImprovement of Brassica napus viainterspecific hybridization between B napus and B oleraceardquoMolecular Breeding vol 34 no 4 pp 1955ndash1963 2014
[25] J Wang L Tian H-S Lee et al ldquoGenome-wide non-additivegene regulation in Arabidopsis allotetraploidsrdquo Genetics vol172 no 1 pp 507ndash517 2006
[26] R Fujimoto J M Taylor S Shirasawa W J Peacock andE S Dennis ldquoHeterosis of Arabidopsis hybrids between C24and Col is associated with increased photosynthesis capacityrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 109 no 18 pp 7109ndash7114 2012
[27] X Ge W Chen S Song WWang S Hu and J Yu ldquoTranscrip-tomic profiling of mature embryo from an elite super-hybridrice LYP9 and its parental linesrdquo BMC Plant Biology vol 8article 114 2008
[28] G Wei Y Tao G Liu et al ldquoA transcriptomic analysis ofsuperhybrid rice LYP9 and its parentsrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 106 no 19 pp 7695ndash7701 2009
[29] G-S Song H-L Zhai Y-G Peng et al ldquoComparative tran-scriptional profiling and preliminary study on heterosis mech-anism of super-hybrid ricerdquo Molecular Plant vol 3 no 6 pp1012ndash1025 2010
[30] N Hoecker B Keller H-P Piepho and F HochholdingerldquoManifestation of heterosis during early maize (Zea mays L)root developmentrdquoTheoretical andAppliedGenetics vol 112 no3 pp 421ndash429 2006
[31] A Thiemann J Fu T A Schrag A E Melchinger M Frischand S Scholten ldquoCorrelation between parental transcriptomeand field data for the characterization of heterosis in Zea maysLrdquoTheoretical and Applied Genetics vol 120 no 2 pp 401ndash4132010
[32] W Qian R Liu and J Meng ldquoGenetic effects on biomass yieldin interspecific hybrids between Brassica napus and B rapardquoEuphytica vol 134 no 1 pp 9ndash15 2003
[33] F Cheng T Mandakova J Wu Q Xie M A Lysak andX Wang ldquoDeciphering the diploid ancestral genome of themesohexaploid Brassica rapardquo Plant Cell vol 25 no 5 pp 1541ndash1554 2013
[34] J Zou J Zhu S Huang et al ldquoBroadening the avenue ofintersubgenomic heterosis in oilseed Brassicardquo Theoretical andApplied Genetics vol 120 no 2 pp 283ndash290 2010
[35] W Qian X Chen D Fu J Zou and J Meng ldquoIntersubgenomicheterosis in seed yield potential observed in a new type ofBrassica napus introgressed with partial Brassica rapa genomerdquoTheoretical and Applied Genetics vol 110 no 7 pp 1187ndash11942005
[36] A Li D Liu JWu et al ldquomRNAand small RNA transcriptomesreveal insights into dynamic homoeolog regulation of allopoly-ploid heterosis in nascent hexaploid wheatrdquo Plant Cell vol 26no 5 pp 1878ndash1900 2014
[37] X Wang H Wang J Wang et al ldquoThe genome of themesopolyploid crop species Brassica rapardquoNature Genetics vol43 no 10 pp 1035ndash1039 2011
[38] M X Zhao B Zhang S L Liu and J Ma ldquoTransposonexpression and potential effects on gene regulation of Brassicarapa andB oleracea genomesrdquoHereditas vol 35 no 8 pp 1014ndash1022 2013
[39] Z An Z Tang BMa et al ldquoTransposon variation by order dur-ing allopolyploidisation between Brassica oleracea and Brassicarapardquo Plant Biology vol 16 no 4 pp 825ndash835 2014
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology