anatomy and physiology of saccadic burst neurons in the alert squirrel monkey. ii. inhibitory burst...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 249~358-380 (1986)
Anatomy and Physiology of Saccadic Burst Neurons in the Alert Squirrel Monkey.
11. Inhibitory Burst Neurons
A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA Department of Neuroscience, The Albert Einstein College of Medicine, New York,
New York 10461
ABSTRACT Electrophysiological and intracellular labelling studies in the cat have
identified a population of saccadic burst neurons in the medullary reticular formation that have an inhibitory, monosynaptic projection to the contralat- era1 abducens nucleus. In the present study, intraaxonal recording and injection of horseradish peroxidase were used to identify and characterize the corresponding population of inhibitory burst neurons (IBNs) in the alert squirrel monkey. Squirrel monkey IBNs are located in the reticular forma- tion ventral and caudal to the abducens nucleus and project contralaterally to the abducens. Additional contralateral projections are present to the vestibular nuclei, the nucleus prepositus, and the pontine and medullary reticular formation rostra1 and caudal to the abducens. All neurons fire a burst of spikes during saccades and are silent during fixation. In most neurons the burst begins 5-15 msec before saccade onset. The number of spikes in the saccadic burst is linearly related to the amplitude of the component of the saccade in the neuron's on-direction. Linear relationships also exist between burst duration and saccade duration and between firing frequency and instantaneous eye velocity. For all neurons, the on-direction is in the ipsilateral hemifield, with a vertical component that may be either upward or downward. Neurons with projections to the vertically related descending and superior vestibular nuclei tend to have on-directions with larger vertical components than neurons that lack these projections. These results, together with those on excitatory burst neurons reported in the preceding paper, demonstrate a reciprocal organization of burst neuron input to the abducens in the monkey similar to that found in the cat and indicate a major role for these neurons in generating the oculomotor activity in motoneurons as well as in other classes of premotor neurons.
Key words: saccades, reticular neurons, abducens nucleus, intracellular staining, horseradish peroxidase
Saccadic eye movements are generated by a burst of fir- ing in motoneurons innervating agonist extraocular mus- cles and a pause in firing in antagonist motoneurons (Robinson, '70; Schiller, '70; Fuchs and Luschei, '70; Henn and Cohen, '73; Goldstein, '83). Electrophysiological studies in the cat suggest that this saccadic modulation of motoneu- ron firing is produced by direct synaptic input from sepa- rate populations of inhibitory and excitatory saccadic burst neurons, which exhibit a burst of firing during saccades and are otherwise silent (Hikosaka and Kawakami, '77; Hikosaka et al., '78b; Igusa et al., '80).
Saccadic burst neurons in the reticular formation caudal to the abducens nucleus exert an inhibitory synaptic action
0 1986 ALAN R. LISS, INC.
Accepted January 29, 1986.
Address reprint requests to A. Strassman at his present address, Neurology Service, Massachusetts General Hospital, Boston, MA 02114.
S.M. Highstein's present address is Department of Otolaryngol- ogy, Washington University School of Medicine, St. Louis, MO 63110.
R.A. McCrea's present address is Committee on Neurobiology, Department of Pharmacological and Physiological Sciences, Uni- versity of Chicago, Chicago, IL 60637.

INHIBITORY BURST NEURONS 359
on contralateral abducens motoneurons, as shown by mi- crostimulation-evoked inhibitory postsynaptic potentials (IPSPs) in abducens motoneurons, and by spike-triggered averaging of abducens field potential and motoneuron membrane potential (Hikosaka and Kawakami, '77; Hiko- saka et al., '78b). Mapping of low-threshold antidromic stimulation sites (Hikosaka et al., '80) and intracellular staining with horseradish peroxidase (HRP) (Yoshida et al., '82) demonstrate dense projections of these inhibitory burst neurons (IBNs) to the abducens as well as projections to the contralateral vestibular nuclei, the nucleus prepositus hy- poglossi, and the pontomedullary reticular formation.
Although IBNs in the cat have been well characterized, no information is presently available on the anatomy or physiology of identified IBNs in the monkey. In the present study, intraaxonal recording and injection of HRP is used to characterize the anatomy and physiology of IBNs in the squirrel monkey. These data, considered together with the findings on excitatory burst neurons in the squirrel monkey described in the preceding paper (Strassman et al., '861, provide the first direct information on the organization of the premotor circuitry underlying the generation of sac- cades in the primate.
METHODS All methods were described in detail in the preceding
paper (Strassman et al., '86). Adult male squirrel monkeys were surgically prepared for chronic recording of eye posi- tion and neuronal activity. Following recovery from sur- gery, neuronal discharge was recorded intraaxonally in alert monkeys during spontaneous saccades. Physiologi- cally characterized neurons were labelled intracellularly by iontophoretic injection of HRP. Monkeys were perfused 24 hours after intracellular labelling, and brain tissue was processed for visualization of HRP-labelled neurons.
Physiological analysis Neuronal firing and eye position channels on the tape
were digitized and analyzed by using a PDP 1173 computer. An analysis of on-direction was carried out for each neuron as described in the preceding paper (Strassman et al., '86). Figure 1 illustrates the on-direction analysis for one IBN, based on its activity during 49 spontaneous saccades. (This neuron is anatomically illustrated in Fig. 12.) In A-D, the number of spikes in a saccadic burst is plotted on the ordinate versus amplitude of the saccade component in a given direction on the abscissa, for four different directions (0", go", 180", 270", where 0" = ipsilateral, 90" = up). Figure 1E shows a plot of the correlation coefficient (filled circles) and slope (open circles) on the ordinate versus the reference direction used for the regression analysis on the abscissa. The filled arrowhead indicates the direction with the highest correlation coefficient (68"; correlation coeffi- cient = .91, slope = 2.1 spikeddegree); the open arrowhead indicates the direction with the highest slope (115 "; corre- lation coefficient = .75, slope = 2.7 spikeddegree). For the neuron shown in Figure 1, the correlation coefficient for the direction with the highest slope is not within .08 of the maximum correlation coefficient, and so 68" was chosen as the on-direction using the criteria described previously (Strassman et al., '86).
An on-direction was calculated for every neuron. The accuracy of the on-direction determination depends on the number of saccades recorded for the neuron. The descrip- tion of on-direction characteristics in the Results (Fig. 4) is
BC BP cx DVN INT LC LVN ML MLF MVN NRTP P PH RB RF RO SO SOL SVN VLVN V PR v SP vl VI N VII VII N X XI1
Abbreviations
brachium conjunctivum brachium pontis external cuneate nucleus descending vestibular nucleus nucleus intercalatus locus coeruleus lateral vestibular nucleus medial lemniscus medial longitudinal fasciculus medial vestibular nucleus nucleus reticularis tegmenti pontis pyramidal tract nucleus prepositus hypoglossi restiform body reticular formation nucleus of Roller superior olive nucleus of the solitary tract superior vestibular nucleus ventral lateral vestibular nucleus principal sensory trigeminal nucleus spinal trigeminal nucleus abducens nucleus abducens nerve facial nucleus facial nerve dorsal motor nucleus of the vagus hypoglossal nucleus
based on 17 IBNs, for which an average of 47 saccades were recorded per neuron. The remaining two IBNs were re- corded during 10-15 saccades and were not included in Figure 4.
Anatomical analysis Anatomical reconstructions and measurements of intra-
cellularly labelled neurons were made by using a camera lucida.
The horizontal maps of Figures 17 and 18A,B are in- tended as qualitative summaries of the projections of EBNs and IBNs. Such maps were first obtained for each neuron and then combined to produce the population maps shown in the figures. For each neuron, each axonal branch was examined on all (coronal) sections on which it appeared for the presence of boutons. For each section on which boutons were present, their distance from the midline (+ 50 pm) and the cytoarchitectural region in which they were located were noted. The locations of all sites containing boutons were mapped for each neuron on a horizontal drawing of the brainstem superimposed on a grid of 80- pm squares. The mediolateral coordinate on the map was determined from the distance from the midline, and the rostrocaudal coordinate was determined from the number of the section. The rostra1 and caudal borders of the abducens nucleus were used as standard reference points for rostrocaudal position, and the lateral borders of the abducens, the pre- positus, and the medial vestibular nucleus were used as mediolateral reference points. Each 80- pm square to which a neuron distributed boutons was marked with a single dot, regardless of the number of boutons; a relatively greater projection of a given neuron to a particular region is re- flected in a larger number of squares to which it contributes a dot. Gradations in the number of boutons in a single 80- pm square were not represented in order to minimize the effect of variability in the intensity of staining on the rela- tive contribution of each neuron to the population map. Each dot in Figures 17 and 18 typically represents 20-50 boutons.

360 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
! .
1 - k , @ - " 1.;-i--- o l d -
0 10 20 0 10 20 0 I 0 20 0 l o 20
A
qUPv . .- down
E. B
L-2.0 270' 0. 90" 180' 270'
DIRECTION
Fig, 1. Determination of on-direction for one inhibitory burst neuron (IBN). A-D. Plots of the number of spikes in the burst (ordinate) versus the amplitude of the saccade in a given direction (abscissa), for four directions (0",90",180",270"). r = correlation coefficient, b = slope. A total of 49 saccades were recorded for this neuron: For each plot, only saccades within 90" of the direction being tested are shown. E. Plot of correlation coefficient (filled circles) and slope (open circles) on the ordinate, versus the reference direction on the abscissa, for the same neuron as in A-D. Filled and open arrowheads indicate the directions with the highest correlation coefficient and slope, respectively.
RESULTS This report describes 19 IBNs, physiologically identified
by their firing during spontaneous saccades, and anatomi- cally classified on the basis of their soma location and their contralateral projection to the abducens nucleus (Hikosaka and Kawakami, '77; Hikosaka et al., '78b, '80) as demon- strated by intracellular staining. Only those burst neurons for which the soma could be located (usually by retrograde labelling following an axonal site of HRP injection) are included in this study.
Physiology Figure 2 shows the firing of an IBN during two saccades
in which the horizontal component is ipsilateral (directed toward the side on which the soma is located) (Fig. 2A) and two saccades in which it is contralateral (2B). The neuron fires a burst of spikes beginning before the onset of ipsilat- era1 saccades (Fig. 2A), and is silent or fires a small number of spikes after the onset of contralateral saccades (Fig. 2B), regardless of whether the saccade has an upward or down- ward vertical component. All the neurons in this study fire exclusively during saccades and are silent during fixation.
All of the neurons fire preferentially for saccades in a particular direction, described as the on-direction for the neuron (see Methods). Figure 3 shows a plot of the number of spikes in the burst (ordinate) versus the amplitude of the component of the saccade in the on-direction (abscissa) for the neuron illustrated in Figure 2. The on-direction calcu- lated for the neuron in Figure 3 is 10. The regression
qUPv - / [ 1 0 m v
down __;__I.. -- 30 ms
Fig. 2. IBN discharge during saccadic eye movements. Each of the panels in A and B shows horizontal (H) eye position, vertical (V) eye position, and intraaxonal recording of membrane potential. A. Ipsilateral saccades. B. Contralateral saccades.
L_
r =0.82 b.1.5 rpikerideg
%1
.. . I
.. - . . . . . . . . ... . . . . . . . . . . .- . .
I 1 ;2" 10" k 6" 4' 2" 0' 2' 4" 6" 8' 10" 1;'
contralateral AMPLITUDE iprilateral
Fig. 3. Relationship between the number of spikes in the burst and the amplitude of the saccade in the on-direction, for the neuron shown in Figure 2. The number of spikes is plotted on the ordinate versus the amplitude of the on-direction component on the abscissa he., modtheta); see Methods). The on-direction calculated for this neuron is 10" (not illustrated). The regression line was calculated by using only saccades with a component in the on-direction (i.e., using only points to the right of the ordinate).
analysis for this plot yields a slope of 1.5 spikeddegree and a correlation coefficient of 0.82. Only saccades with a com- ponent in the on-direction (the points to the right of the ordinate) were used in the regression analysis. The mean value of the slope for the population is 1.9 f 1.4 spiked degree; the mean correlation coefficient is 0.79 k 0.23. These values are similar to those calculated for the EBNs in the preceding paper (Strassman et al., '86).
Most of the neurons fire a burst during saccades perpen- dicular to the on-direction (saccades for which the on-direc- tion component is 0); this is indicated by a positive intercept
INHIBITORY BURST NEURONS 361
IPS1 LATERAL
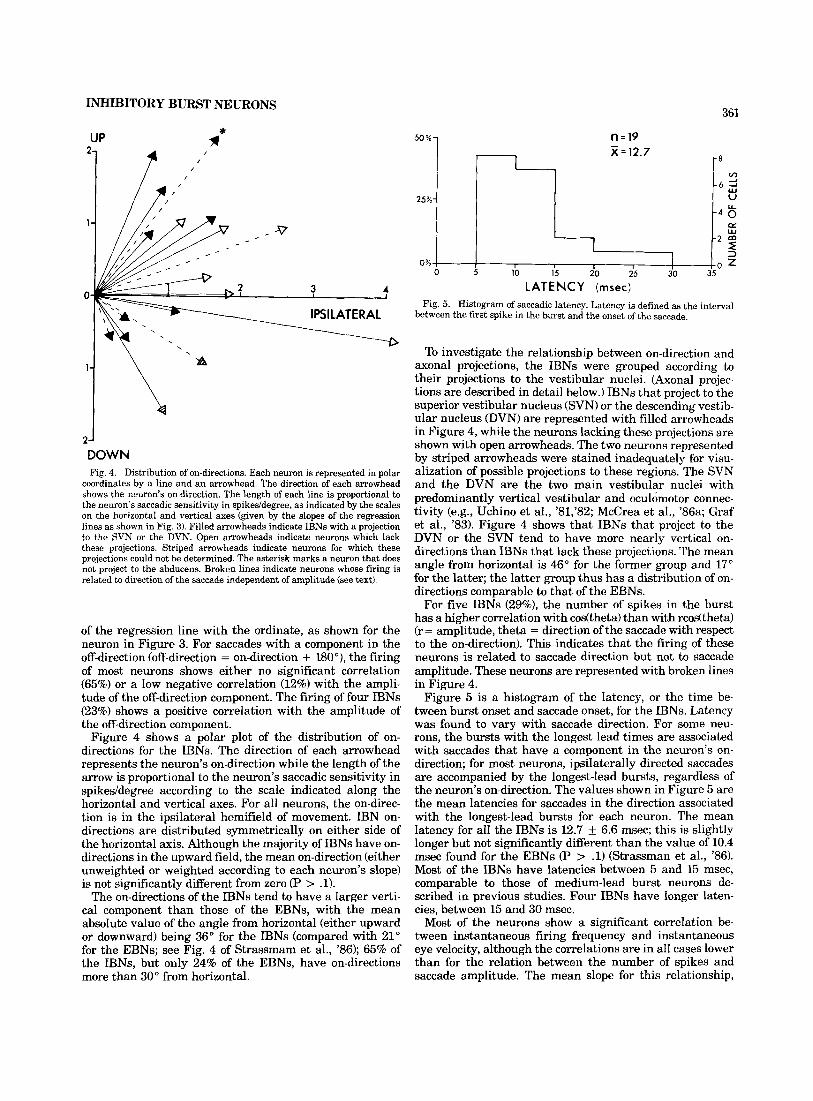
DOWN Fig. 4. Distribution of on-directions. Each neuron is represented in polar
coordinates by a line and an arrowhead. The direction of each arrowhead shows the neuron's on-direction. The length of each line is proportional to the neuron's saccadic sensitivity in spikeddegree, as indicated by the scales on the horizontal and vertical axes (given by the slopes of the regression lines as shown in Fig. 3). Filled arrowheads indicate IBNs with a projection to the SVN or the DVN. Open arrowheads indicate neurons which lack these projections. Striped arrowheads indicate neurons for which these projections could not be determined. The asterisk marks a neuron that does not project to the abducens. Broken lines indicate neurons whose firing is related to direction of the saccade independent of amplitude (see text).
of the regression line with the ordinate, as shown for the neuron in Figure 3. For saccades with a component in the off-direction (off-direction = on-direction + 180°), the firing of most neurons shows either no significant correlation (65%) or a low negative correlation (12%) with the ampli- tude of the off-direction component. The firing of four IBNs (23%) shows a positive correlation with the amplitude of the off-direction component.
Figure 4 shows a polar plot of the distribution of on- directions for the IBNs. The direction of each arrowhead represents the neuron's on-direction while the length of the arrow is proportional to the neuron's saccadic sensitivity in spikeddegree according to the scale indicated along the horizontal and vertical axes. For all neurons, the on-direc- tion is in the ipsilateral hemifield of movement. IBN on- directions are distributed symmetrically on either side of the horizontal axis. Although the majority of IBNs have on- directions in the upward field, the mean on-direction (either unweighted or weighted according to each neuron's slope) is not significantly different from zero (P > .1).
The on-directions of the IBNs tend to have a larger verti- cal component than those of the EBNs, with the mean absolute value of the angle from horizontal (either upward or downward) being 36" for the IBNs (compared with 21" for the EBNs; see Fig. 4 of Strassmam et al., '86); 65% of the IBNs, but only 24% of the EBNs, have on-directions more than 30" from horizontal.
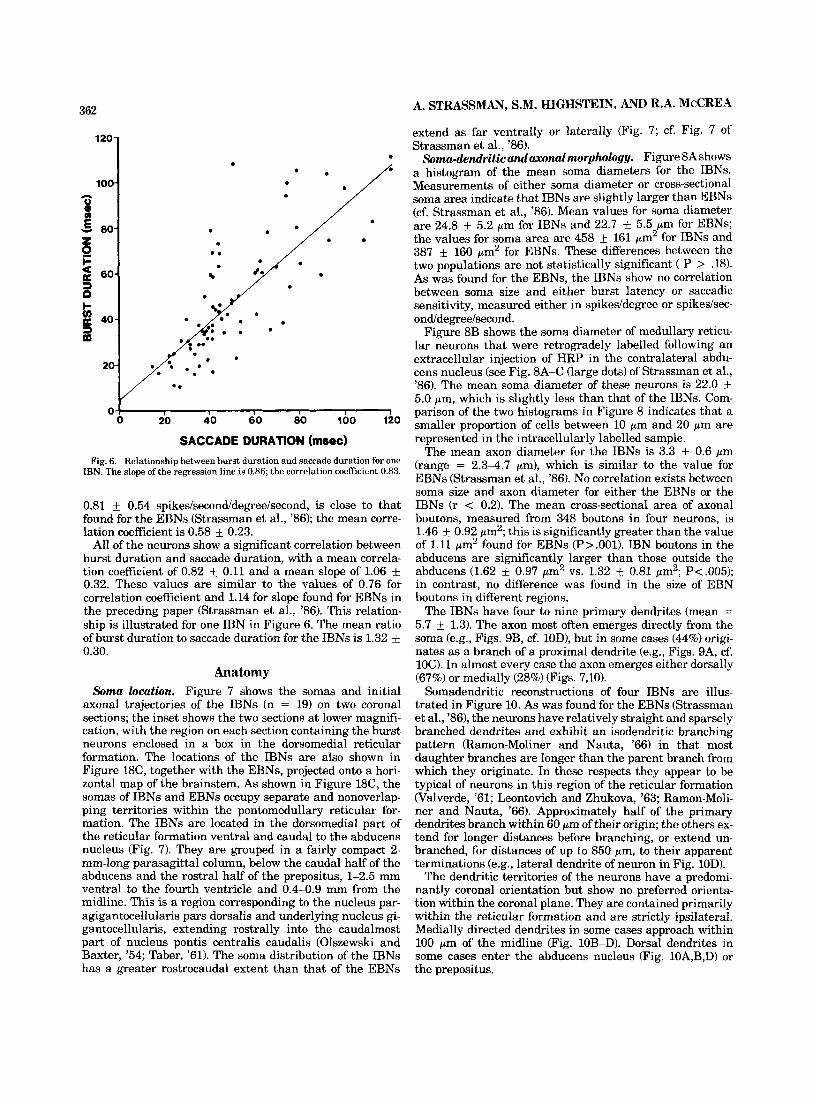
n=19 x = 12.7
LATE N C Y (msec) Fig. 5. Histogram of saccadic latency. Latency is defined as the interval
between the first spike in the burst and the onset of the saccade.
To investigate the relationship between on-direction and axonal projections, the IBNs were grouped according to their projections to the vestibular nuclei. (Axonal projec- tions are described in detail below.) IBNs that project to the superior vestibular nucleus (SVN) or the descending vestib- ular nucleus (DVN) are represented with filled arrowheads in Figure 4, while the neurons lacking these projections are shown with open arrowheads. The two neurons represented by striped arrowheads were stained inadequately for visu- alization of possible projections to these regions. The SVN and the DVN are the two main vestibular nuclei with predominantly vertical vestibular and oculomotor connec- tivity (e.g., Uchino et al., '81,'82; McCrea et al., '86a; Graf et al., '83). Figure 4 shows that IBNs that project to the DVN or the SVN tend to have more nearly vertical on- directions than IBNs that lack these projections. The mean angle from horizontal is 46" for the former group and 17" for the latter; the latter group thus has a distribution of on- directions comparable to that of the EBNs.
For five IBNs (29%), the number of spikes in the burst has a higher correlation with codtheta) than with rcodtheta) (r = amplitude, theta = direction of the saccade with respect to the on-direction). This indicates that the firing of these neurons is related to saccade direction but not to saccade amplitude. These neurons are represented with broken lines in Figure 4.
Figure 5 is a histogram of the latency, or the time be- tween burst onset and saccade onset, for the IBNs. Latency was found to vary with saccade direction. For some neu- rons, the bursts with the longest lead times are associated with saccades that have a component in the neuron's on- direction; for most neurons, ipsilaterally directed saccades are accompanied by the longest-lead bursts, regardless of the neuron's on-direction. The values shown in Figure 5 are the mean latencies for saccades in the direction associated with the longest-lead bursts for each neuron. The mean latency for all the lBNs is 12.7 k 6.6 msec; this is slightly longer but not signifkantly different than the value of 10.4 msec found for the EBNs (P 7 .l) (Strassman et al., '86). Most of the IBNs have latencies between 5 and 15 msec, comparable to those of medium-lead burst neurons de- scribed in previous studies. Four IBNs have longer laten- cies, between 15 and 30 msec.
Most of the neurons show a significant correlation be- tween instantaneous firing frequency and instantaneous eye velocity, although the correlations are in all cases lower than for the relation between the number of spikes and saccade amplitude. The mean slope for this relationship,
362
leu-
100-
00-
60-
40 -
20-
0
A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
extend as far ventrally or laterally (Fig. 7; cf. Fig. 7 of Strassman et al., '86).
Soma-dendn'ticandaxonal morphology. Figure 8A shows a histogram of the mean soma diameters for the IBNs. Measurements of either soma diameter or cross-sectional soma area indicate that IBNs are slightly larger than EBNs (cf. Strassman et al., '86). Mean values for soma diameter are 24.8 & 5.2 pm for IBNs and 22.7 5.5 pm for EBNs; the values for soma area are 458 k 161 pm2 for IBNs and 387 & 160 pm2 for EBNs. These differences between the two populations are not statistically significant ( P > .18). As was found for the EBNs, the IBNs show no correlation between soma size and either burst latency or saccadic sensitivity, measured either in spikeddegree or spikedsec- ond/degree/second.
Figure 8B shows the soma diameter of medullary reticu- lar neurons that were retrogradely labelled following an extracellular injection of HRP in the contralateral abdu- cens nucleus (see Fig. 8A-C (large dots) of Strassman et al., '86). The mean soma diameter of these neurons is 22.0 k 5.0 pm, which is slightly less than that of the IBNs. Com- parison of the two histograms in Figure 8 indicates that a smaller proportion of cells between 10 pm and 20 pm are represented in the intracellularly labelled sample.
The mean axon diameter for the IBNs is 3.3 k 0.6 pm (range = 2.3-4.7 pm), which is similar to the value for EBNs (Strassman et al., '86). No correlation exists between soma size and axon diameter for either the EBNs or the IBNs (r < 0.2). The mean cross-sectional area of axonal boutons, measured from 348 boutons in four neurons, is 1.46 k 0.92 pm2; this is significantly greater than the value of 1.11 ,urn2 found for EBNs (P>.OOl). IBN boutons in the abducens are significantly larger than those outside the abducens (1.62 k 0.97 pm2 vs. 1.32 k 0.81 pm2; P<.OO5); in contrast, no difference was found in the size of EBN boutons in different regions.
The IBNs have four to nine primary dendrites (mean = 5.7 k 1.3). The axon most often emerges directly from the soma (e.g., Figs. 9B, cf. lOD), but in some cases (44%) origi- nates as a branch of a proximal dendrite (e.g., Figs. 9A, cf. 1OC). In almost every case the axon emerges either dorsally (67%) or medially (28%) (Figs. 7,101.
Somadendritic reconstructions of four IBNs are illus- trated in Figure 10. As was found for the EBNs (Strassman et al., '861, the neurons have relatively straight and sparsely branched dendrites and exhibit an isodendritic branching pattern (Ramon-Moliner and Nauta, '66) in that most daughter branches are longer than the parent branch from which they originate. In these respects they appear to be typical of neurons in this region of the reticular formation (Valverde, '61; Leontovich and Zhukova, '63; Ramon-Moli- ner and Nauta, '66). Approximately half of the primary dendrites branch within 60 pm of their origin; the others ex- tend for longer distances before branching, or extend un- branched, for distances of up to 850 pm, to their apparent terminations (e.g., lateral dendrite of neuron in Fig. 10D).
The dendritic territories of the neurons have a predomi- nantly coronal orientation but show no preferred orienta- tion within the coronal plane. They are contained primarily within the reticular formation and are strictly ipsilateral. Medially directed dendrites in some cases approach within 100 pm of the midline (Fig. 10B-D). Dorsal dendrites in some cases enter the abducens nucleus (Fig. 10A,B,D) or the prepositus.
*
I I I I I 0 20 40 60 00 100 120
.
SACCADE DURATION (msec) Fig. 6. Relationship between burst duration and saccade duration for one
IBN. The slope of the regression line is 0.86; the correlation coefficient 0.83.
0.81 +_ 0.54 spikes/second/degree/second, is close to that found for the EBNs (Strassman et al., '86); the mean corre- lation coefficient is 0.58 & 0.23.
All of the neurons show a significant correlation between burst duration and saccade duration, with a mean correla- tion coefficient of 0.82 k 0.11 and a mean slope of 1.06 & 0.32. These values are similar to the values of 0.76 for correlation coefficient and 1.14 for slope found for EBNs in the preceding paper (Strassman et al., '86). This relation- ship is illustrated for one IBN in Figure 6. The mean ratio of burst duration to saccade duration for the IBNs is 1.32 k 0.30.
Anatomy Soma location. Figure 7 shows the somas and initial
axonal trajectories of the IBNs (n = 19) on two coronal sections; the inset shows the two sections at lower magnifi- cation, with the region on each section containing the burst neurons enclosed in a box in the dorsomedial reticular formation. The locations of the IBNs are also shown in Figure 18C, together with the EBNs, projected onto a hori- zontal map of the brainstem. As shown in Figure 18C, the somas of IBNs and EBNs occupy separate and nonoverlap- ping territories within the pontomedullary reticular for- mation. The IBNs are located in the dorsomedial part of the reticular formation ventral and caudal to the abducens nucleus (Fig. 7). They are grouped in a fairly compact 2- mm-long parasagittal column, below the caudal half of the abducens and the rostra1 half of the prepositus, 1-2.5 mm ventral to the fourth ventricle and 0.4-0.9 mm from the midline. This is a region corresponding to the nucleus par- agigantocellularis pars dorsalis and underlying nucleus gi- gantocellularis, extending rostrally into the caudalmost part of nucleus pontis centralis caudalis (Olszewski and Baxter, '54; Taber, '61). The soma distribution of the IBNs has a greater rostrocaudal extent than that of the EBNs

INHIBITORY BURST NEURONS
VI
363
I
Fig. 7. Soma locations and initial axonal trajectories of IBNs. IBNs are shown on two coronal sections at the level of and caudal to the ahducens. The inset shows the two sections at lower magnification with the areas containing the IBNs enclosed by a box in the dorsomedial reticular formation on each section. The arrowhead in the first section points to an axon collateral ipsilateral to the soma. Calibration = 200 pm.
The IBN in Figure 10A has an unusually dorsal location, at the ventromedial border of the abducens. A substantial portion of its dendritic territory is within the abducens nucleus. This cell's initial axonal trajectory is also unusual, in that the axon ascends rostrally in the VIIth nerve before crossing the midline; however, its projection sites are typi- cal for IBNs (not illustrated).
Axonal trajectory and brunching. IBN axons usually cross the midline at the level of or slightly rostral to the soma (Figs. 7, 15; 12, bottom section), and in all cases give rise to a finer caudal collateral in the contralateral reticu- lar formation within 500 p m of the midline (e.g., Figs. lOC, 15; 12, bottom section). This axon collateral branches and gives rise to boutons in the reticular formation ventral to
the prepositus and the caudal abducens (Figs. 12, bottom section, 14, bottom section). It also usually has additional sites of arborization in the abducens, the prepositus, or the medial vestibular nucleus (MVN). Neither the caudal col- lateral nor the main axon and its other collaterals extend further caudally than the caudal border of the prepositus.
The main axons generally course rostrally a short dis- tance in the contralateral reticular formation, and in some cases (44%) give rise to a rostral collateral that branches and terminates in the EBN region of the pontine reticular formation rostral to the abducens (e.g., Figs. 12, top section; 14, top section; 15A). Neither this collateral nor the main axon or any of its other branches ever extends rostrally beyond the rostral limit of the region of EBN somas found

364 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
50 - 40 -
6 30-
i3Q 20-
10-
0
Y J
A. IBNS
>
- I I I 1
- X=25 urn
B. MEDULLARY RF CELLS urn
O K 10 15 20 25 30 35 40
SOMA DIAMETER (urn) Fig. 8. Histograms of soma diameter. A. IBNs. B. Medullary reticular
neurons retrogradely labelled from an HRP injection in the contralateral abducens nucleus (see Fig. 8A-C of Strassman et al., '86).
in the previous study (Strassman et al., '86), approximately 1 mm rostral to the abducens. Thus, IBNs do not project to the midbrain or to the more rostral regions of the pontine reticular formation.
The main axons enter the abducens medially or ventrally and typically course caudolaterally through the nucleus, giving off several collaterals that arborize extensively and distribute hundreds of boutons in all regions of the nucleus (Figs. 12, middle section; 13). Some of the abducens collat- erals exit the nucleus caudally and complete their terminal arborization in the adjacent region of the MVN, along the caudal border of the abducens (Figs. 12, bottom section; 15A, cf. 17B). The main axon typically exits the nucleus laterally and courses caudally for a short distance, giving off collaterals that branch and terminate in the rostrolat- era1 part of the MVN (Figs. 11,12); in some cases (47%), the arborization of the main axon or its collaterals extends into the rostromedial part of the DVN and the ventral part of the lateral vestibular nucleus (LVN) immediately rostral to the DVN (Figs. 11,15B); 25% of the IBNs also have a rostral collateral to the SVN (Fig. 12, middle section).
One IBN axon gives rise to a short collateral just before crossing the midline that terminates in the ipsilateral retic- ular formation between the abducens nuclei, within 300 pm
of the midline (arrowhead in Fig. 7); in all other cases, IBN projection sites are strictly contralateral.
Figure 9D and E show portions of the arborization of art IBN in the right vestibular nuclei, in the DVN, and the adjacent ventrolateral part of the MVN. This region is a common target of IBN projections. Figure 11 is a recon- struction of the axonal arborization of an IBN in the left vestibular complex. Three major axonal branches are shown exiting the abducens nucleus laterally and arborizing through a region of the vestibular nuclei at the level of, and slightly caudal to, the caudal part of the abducens. This neuron arborizes heavily in the region lateral to the ventro- lateral corner of the MVN and ventral to the dorsal part of the LVN. This region (labelled DVN-VLVN) is occupied by the ventral LVN at the level of the abducens and is replaced caudally by the rostral pole of the DVN. The neuron also arborizes heavily in the MVN, especially along the lateral border. As is true for all IBNs, the arborization remains almost entirely outside the dorsal part of the LVN.
Figure 12 illustrates the axonal trajectory of an IBN with a more caudally distributed arborization in the MVN and the DVN, as well as a projection to the SVN. (The physiol. ogy of this neuron is illustrated in Fig. 1.) Circles and triangles indicate the points at which an axon or collateral passes onto the caudally or rostrally adjacent section, re- spectively. (Rostra1 circles connect to caudal triangles.) The soma is shown on the lower (caudal) section in the reticular formation ventral to the prepositus. The axon crosses the midline at the level of the soma, and gives rise to a caudal collateral which arborizes in the contralateral reticular for- mation and the MVN. The main axon ascends in the medial reticular formation and enters the ventromedial corner of the abducens (middle section), where it gives rise to a ros- tral collateral that ascends to the reticular formation ros- tral to the abducens (upper section). This collateral arborizes in the dorsomedial reticular formation and the adjacent cellular region within the ventromedial part of the MLF. The main axon courses laterally and caudally through the abducens, giving off several collaterals, and exits the nu- cleus caudally, along with two other branches in the abdu- cens (marked with circles on the middle section). The axon is shown on the lower section continuing laterally across the MVN and arborizing heavily in the MVN and the medial DVN. In addition, it gives rise to a rostral collateral that ascends through the LVN without giving rise to bou- tons, and enters the SVN (middle section), where it arbor- izes and terminates.
Figure 13 illustrates the terminations of one IBN in the abducens nucleus. Although the pattern of terminations in the abducens varies for different neurons, no relationship could be seen between this pattern and the neurons' phys- iological characteristics. The IBN population as a whole projects to all regions of the nucleus (Fig. 16C,D).
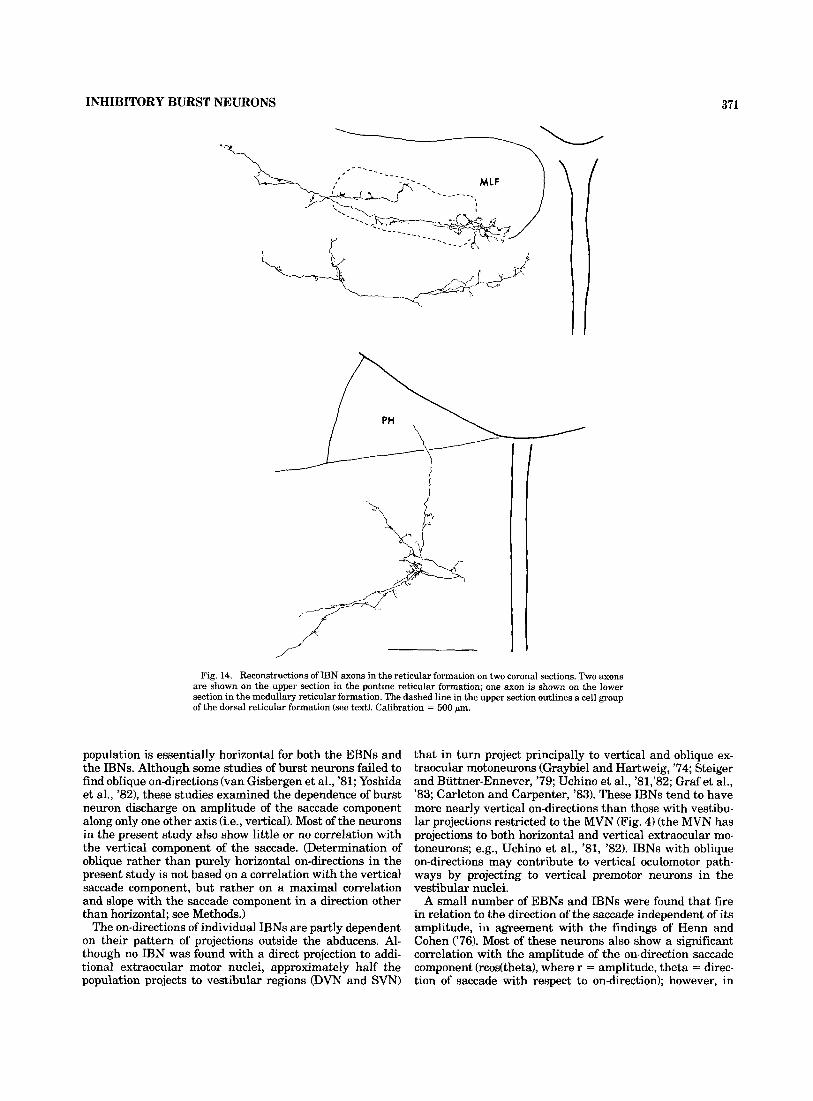
Figure 14 shows the arborization of two IBN axons in the reticular formation rostral to the abducens (upper section) and one IBN axon in the reticular formation caudal to the abducens (lower section). For all IBNs, these projections are confined primarily to the dorsal half of the reticular forma- tion. In some cases they extend medially into the raphe and dorsally into cellular regions within the MLF (Figs. 12, top section; 14, top section). The dashed line in Figure 14 out- lines a cellular region of the dorsal reticular formation that extends into the lateral MLF and is a target of IBN projec- tions. This cell group, which has been called the dorsal paramedian pontine reticular formation (McCrea et al., '%a), is present immediately rostral to the abducens, and

Fig. 9. A. IBN soma in the left reticular formation; reconstruction is shown in Figure 1OC. B. IBN soma in the right reticular formation; recon- struction is shown in Figure 10D. Axonal swelling marks the point of electrode penetration and injection of HRP. C. Two IBN somas in the left
reticular formation ventral to the prepositus. D,E. Portions of an IBN arborization in the right vestibular nuclei, along the border between the MVN and the DVN; a diagram of this neuron is shown in Figure 15B. Calibration = 50 p m in A,B 125 pm in C-E.
includes the dorsal part of the EBN region (6. Fig. 11B of Strassman et al., '86)
One IBN has a sparse projection to the nucleus of Roller, at the level of the caudal border of the prepositus (Figs. 16H, bottom of 17B).
Figure 15 diagrams the projections of two IBNs on hori- zontal maps of the brainstem. Filled circles are used to represent regions of termination in the abducens, the pre- positus, and the vestibular nuclei; open circles represent terminations in the reticular formation ventral to these nuclei. The neuron in 15A projects heavily to the abducens and has additional projections to the MVN and regions of the reticular formation rostral and caudal to the abducens. The neuron in 15B has a lighter projection to the abducens but has an extremely dense arborization in the region around the adjoining borders of the MVN, the DVN, and the ventral LVN (photomicrographs of this arborization are shown in Fig. 9D,E). This neuron also has a light projection to the prepositus and the reticular formation caudal to the abducens, but lacks a collateral to the reticular formation rostral to the abducens.
The two neurons in Figure 15 are representative of two subgroups that can be discerned among the IBNs. As men-
tioned above, every IBN projects to the MVN (as well as to the abducens and the reticular formation caudal to the abducens); however, only about half of the IBNs have addi- tional vestibular projections (i.e., to the SVN, the DVN, or the ventral LVN). This criterion for subdividing the popu- lation is of interest because the two subgroups have differ- ent on-directions (Fig. 4); as mentioned above, IBNs that project to the SVN or the DVN (filled arowheads in Fig. 4) tend to have more nearly vertical on-directions than IBNs that lack these projections. (All neurons that project to the ventral LVN also project to the DVN, and so are implicitly included in the first subgroup, represented by filled arrow- heads in Fig. 4.) The two neurons in Figure 15A and B are representative of this trend; their on-directions are 20" and 58", respectively.
An additional anatomical difference between the two subgroups is in the collateral to the reticular formation rostral to the abducens; in most cases (€is%), this collateral is present only on neurons that lack the more extended vestibular projection (the neuron illustrated in Fig. 12 is an exception).
Figure 16 summarizes the regions of termination for the entire population of IBNs on eight coronal sections. The

B
Fig. 10. Coronal soma-dendritic reconstructions of IBNs. Axons are indi- cated by arrows. The course of the axon to the midline is shown for each cell. Calibration = 100 pm for A-D.
VI
VI D

R F
Fig
11
Cor
onal
reco
nstr
uctio
n of
the
axo
nal
arbo
riza
tion
of a
n IB
N I
n th
e le
ft v
estib
ular
nuc
lei
The
pos
ition
of t
he la
tera
l bor
der o
f the
abd
ucen
s is
sho
wn
at th
e ro
stro
caud
el le
vel w
here
the
maj
or a
xona
l bra
nche
s exi
t the
nu
cleu
s, h
owev
er, t
he ar
bori
zatio
n in
the
vest
ibul
ar n
ucle
i is f
urth
er ca
udal
, at
the
leve
l w
here
the
abd
ucen
s ha
s de
crea
sed
in m
edio
late
ral
exte
nt,
as
wel
l as
imm
edia
tely
cau
dal t
o th
e nu
cleu
s T
he r
eeon
mar
ked
DV
N-V
LVN
1s
occ
upie
d by
the
ven
tral
LV
N a
t the
leve
l of
the
cau
dal a
bduc
ens,
and
IS
repl
aced
fur
ther
cau
dally
by
the
rost
ra1
pole
of
the
DV
N (
cf
Fig
17B
) C
alib
ratio
n =
400
Fm

368 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
heaviest terminations are present in the abducens, the MVN, the DVN, the ventral LVN, and the reticular forma- tion rostral and caudal to the abducens. Lighter projections are present in the SVN and the nucleus prepositus.
One neuron has been included with the IBNs that is similar in soma location and all aspects of axonal trajectory, branching, and termination, except that it lacks a projec- tion to the abducens; this neuron is indicated with an aster- isk in Figure 4.
One contralaterally projecting burst neuron was found that differs from the IBNs in having a more caudal soma location (filled triangle in Fig. 18C), a smaller soma and axon diameter, and a more ventral axonal course through the reticular formation. Its projections are also different, in being restricted medially to the prepositus and the medial part of the MVN. It also projects to the medullary reticular formation, but no projection was seen to the abducens. This neuron was not included with the IBNs in any of the phys- iological or anatomical analyses in this paper.
One burst neuron found in the IBN region is unique in that it projects to the abducens ipsilaterally rather than contralaterally. (This neuron has not been included in any of the figures or analyses.) This neuron has additional pro- jections ipsilaterally to the prepositus and the pontine and medullary reticular formation, and bilaterally to the sub- ventricular region rostral to the abducens and dorsal to the MLF. The on-direction for this neuron is ipsilateral, which suggests it has an excitatory synaptic action on its target neurons in the ipsilateral abducens.
Topography of projections: Comparison with EBNs The vestibular nuclei and the nucleus prepositus.
Although both IBNs and EBNs project to the vestibular nuclei and the prepositus, their areas of termination within these nuclei are different and largely nonoverlapping. Fig- ure 17 summarizes the projections of EBNs and IBNs in the abducens, the prepositus, and the vestibular nuclei on two horizontal maps of the brainstem. The density of dots in an area indicates the density of projections to that area (see Methods). The map for IBNs (Fig. 17B) suggests two pri- mary focal points outside of the abducens: one appears in the rostromedial corner of the DVN and extends rostrally into the ventral part of the LVN and medially into the rostrolateral part of the MVN; the other appears more medially in the rostral MVN, immediately caudolateral to the abducens. A considerably lighter zone is present in the prepositus and the adjacent region of the MVN.
In contrast, the EBNs' heaviest terminations outside the abducens are in the prepositus and contiguous regions of the MVN (Fig. 17A). EBNs also have a heavy projection to the region of the MVN immediately caudal t o the abducens; this is the only region in the vestibular nuclei or the pre- positus that receives a heavy projection from both burst neuron populations.
Reticular formation. Figure 18 shows three horizontal maps comparing the soma locations of the EBNs and IBNs (Fig. 18C) with the regions of the reticular formation receiv- ing projections from EBNs (Fig. 18A) and IBNs (Fig. 18B). The figure indicates two rostrocaudally separate regions of termination in the reticular formation for both the EBNs and the IBNs. The rostral projection zone of the IBNs closely matches the region containing EBN somas (Fig. 18C) in its rostrocaudal and mediolateral extent. The EBNs' rostral termination zone is considerably sparser, and extends fur- ther rostrally but not as far laterally, compared with that
of the IBNs. The caudal termination zones of both EBNs and IBNs cover the region containing the IBN somas.
DISCUSSION The present results, together with those of the preceding
study (Strassman et al., '86), confirm the existence of a premotor saccadic organization in the monkey similar to that found in the cat. In particular, these studies demon- strate that the abducens nucleus receives a major input from a class of exclusively saccade-related neurons, and that this input is reciprocally organized into separate ipsi- laterally and contralaterally projecting burst neuron populations.
In either species, the synaptic action of these two burst neuron populations can be inferred as excitatory and inhib- itory, respectively, by comparing their on-directions with those of their target neurons in the abducens. This conclu- sion is strengthened by results of electrical stimulation in the cat, whereby monosynaptic excitatory postsynaptic po- tentials (EPSPs) or IPSPs are produced in abducens moto- neurons, depending on whether the stimulating electrode is in, respectively, the ipsilateral reticular formation rostral to the abducens (Highstein et al., '76) or the contralateral reticular formation caudal to the abducens (Hikosaka and Kawakami, '77). These synaptic actions are further sup ported by electrophysiological studies in the cat of individ- ual burst neurons antidromically identified as projecting to the abducens: spike-triggered averaging reveals a mono- synaptic IPSP in abducens motoneurons following contra- lateral burst neuron spikes (Hikosaka et al., '78b), and a slow negativity in the extracellular field, indicating mono- synaptic excitatory postsynaptic currents, following ipsilat- era1 burst neuron spikes (Igusa et al., '80). Previous anatomical studies indicate the same basic organization of reticular formation projections to the abducens in the mon- key, although direct electrophysiological evidence is lack- ing. The present results confirm the projections of individual burst neurons to the abducens, extending the established anatomical similarity with the cat to the cellular level, and further justifying the description of these two populations in the monkey as excitatory and inhibitory burst neurons.
Physiology In common with medium-lead burst neurons of previous
studies in the cat and monkey, both the EBNs and the IBNs of the present studies have a firing pattern appropriate for providing the immediate premotor saccadic input to abdu- cens motoneurons. Specifically, the firing of these burst neurons is similar to that of the motoneurons in its close relationship with the onset, duration, and amplitude of the saccade.
The main values for latency from burst onset to saccade onset found for the EBNs (10.4 msec) and IBNs (12.7 msec) are similar to the latencies for burst neurons classified as medium-lead in previous studies in the cat (Kaneko et al., '81; Yoshida et al., '82) and rhesus monkey (Luschei and Fuchs, '72; Cohen and Henn, '72; Keller, '74; van Gisbergen et al., '81). These latency measurements indicate that onset of burst neuron discharge typically precedes the saccadic burst or pause of the motoneurons by approximately 2-5 msec (e.g., van Gisbergen et al., '81, in rhesus; unpublished observations on motoneurons in the squirrel monkey). Thus, not only has the synaptic connection of burst neurons with motoneurons been established, but also the timing of burst neuron discharge relative to the saccadic modulation in the

369 INHIBITORY BURST NEURONS
Fig. 12. Reconstruction of the full axonal trajectory of an IBN on three coronal sections. Circles and triangles mark the points at which axonal branches pass onto the caudally or rostratly adjacent section, respectively. Calibration = 500 pm.

370 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
Fig. 13. Terminations of an IBN in the right abducens nucleus on six coronal sections. The sections proceed caudally from left to right, starting at the upper left. Each dot represents a single bouton. Three adjacent 80-pm sections were combined for each section in the figure. Approximately 2,400 boutons are represented.
motoneurons is consistent with a primary role for burst neurons in generating that modulation.
The neurons of the present studies show both the strong correlations between burst duration and saccade duration. and between burst size and saccade amplitude, character- istic of both medium-lead burst neurons (Cohen and Henn, '72; Luschei and Fuchs, '72; Keller, '74; van Gisbergen et
al., '81; Yoshida et al., '82) and motoneurons (e.g., Robinson, '70; Henn and Cohen, '73). Consistent with most previous reports of medium-lead burst neurons, the on-direction al- ways has an ipsilateral component, and in many cases also has a vertical component that may be either up or down (Luschei and Fuchs, '72; Keller, '74; Kaneko et al., '81; Kaneko and Fuchs, '81). The mean on-direction for the
INHIBITORY BURST NEURONS 371
I
Fig. 14. Reconstructions of IBN axons in the reticular formation on two coronal sections. Two axons are shown on the upper section in the pontine reticular formation; one axon is shown on the lower section in the medullary reticular formation. The dashed line in the upper section outlines a cell group of the dorsal reticular formation (see text). Calibration = 500 pm.
population is essentially horizontal for both the EBNs and the IBNs. Although some studies of burst neurons failed to find oblique on-directions (van Gisbergen et al., '81; Yoshida et al., '82), these studies examined the dependence of burst neuron discharge on amplitude of the saccade component along only one other axis (i.e., vertical). Most of the neurons in the present study also show little or no correlation with the vertical component of the saccade. (Determination of oblique rather than purely horizontal on-directions in the present study is not based on a correlation with the vertical saccade component, but rather on a maximal correlation and slope with the saccade component in a direction other than horizontal; see Methods.)
The on-directions of individual IBNs are partly dependent on their pattern of projections outside the abducens. Al- though no IBN was found with a direct projection to addi- tional extraocular motor nuclei, approximately half the population projects to vestibular regions (DVN and SVN)
that in turn project principally to vertical and oblique ex- traocular motoneurons (Graybiel and Hartweig, '74; Steiger and Buttner-Ennever, '79; Uchino et al., '81,'82; Grafet al., '83; Carleton and Carpenter, '83). These IBNs tend to have more nearly vertical on-directions than those with vestibu- lar projections restricted to the It" (Fig. 4) (the MVN has projections to both horizontal and vertical extraocular mo- toneurons; e.g., Uchino et al., '81, '82). IBNs with oblique on-directions may contribute to vertical oculomotor path- ways by projecting to vertical premotor neurons in the vestibular nuclei.
A small number of EBNs and IBNs were found that fire in relation to the direction of the saccade independent of its amplitude, in agreement with the findings of Henn and Cohen ('76). Most of these neurons also show a significant correlation with the amplitude of the on-direction saccade component (rcos(theta), where r = amplitude, theta = direc- tion of saccade with respect to on-direction); however, in

372 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
I I //I I
I /
//I
I
I
I
I I
I
I
I
I I
I
I
I
I
I
I
I
I
MVN i
I I
I
I
I
I
I I
I
I
I
I
I
I
I
I
B
I
I
I
I I
I
I
I
I
I
I
I
I
I
I
I
I
I
Fig. 15. Diagrams of two IBNs on horizontal maps of the brainstem. Filled circles mark regions in the abducens, the prepositus, and t'le vestibular nuclei containing axonal boutons; open circles indicate regions containing boutons in the reticuiar formation ventral to these nuclei. The dashed line represents the midline on each section.
each case the correlation with codtheta) is higher, indicat- ing amplitude is an unrelated variable (i.e., the correlation with rcodtheta) is fortuitous, resulting from the correlation between rcodtheta) and cos(theta); Kaneko et al. ('81) ap- parently discounted this explanation in reaching their con- clusion that no neurons code direction independent of amplitude).
The only previous study that compared the physiological properties of EBNs and IBNs (Kaneko and Fuchs, '81) found differences that can be seen to varying degrees in the pres- ent data. These authors, working in the cat, observed a similarly wide distribution of on-directions for IBNs as com- pared with EBNs. In addition, they found a longer burst duration relative to saccade duration for IBNs than EBNs, a trend that can be discerned in the present results but with low statistical significance (P > .1). However, the two populations of burst neurons in the squirrel monkey show similarly high correlations between burst duration and sac-
cade duration whereas in the cat, EBNs have significantly lower correlations. Also, the substantially larger slope of IBNs than EBNs in the cat for the relation between number of spikes and saccade amplitude was not found in the squir- rel monkey. Overall, EBNs and IBNs in the squirrel mon- key are essentially similar in their coding of saccade parameters and cannot be identified or distinguished solely on the basis of recordings of unit activity in the alert animal.
Soma location and morphology
The IBN soma locations reported here coincide closely with those found in the cat (Hikosaka and Kawakami, '77; Kaneko and Fuchs, '81; Yoshida et al., '821, differing mainly in that Hikosaka and Kawakami found a small number of neurons rostra1 and dorsal to the abducens. In both species, the IBNs are found primarily in the dorsomedial part of the

INHIBITORY BURST NEURONS 373
Fig. 16. Summary of IBN projections on eight coronal sections. The sections proceed caudally from A to H, and are separated by 600-800 pm. The density of dots in each region represents the density of terminations for the entire population of IBNs.

374 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
from previous page.
reticular formation, ventral and ventromedial to the caudal part of the abducens and the rostra1 part of the prepositus.
The highly specialized functional role of the burst neu- rons is not reflected in any obvious way in their dendritic morphology, which is characteristic of so-called generalized neurons of the pontomedullary reticular formation (Val- verde, '61; Leontovich and Zhukova, '63; Ramon-Moliner and Nauta, '66).
Projections Previous studies of projections from the burst neuron
regions of the reticular formation that used extracellular
tracers succeeded in labelling all of the burst neuron projec- tion sites demonstrated in the present study. However, it is only by determining the projections of individual behav- iorally identified burst neurons that their projections can be distinguished from separate projections arising from un- related neurons in the same regions. In addition, intracel- lular staining is necessary to discern differences in the weighting of burst neuron projections to different regions, as well as to determine differences between the projections of burst neurons with different physiological characteristics.
A previous anterograde transport study in the cat (Gray- biel, '77) demonstrated contralateral projections from the

INHIBITORY BURST NEURONS
A. EBNS 375
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I I
I
I
I
I
I I
I
I
I
1
I
I
\
I
I
I
I
I
I
I
I
I
I
I
I
I
I
1
I
I
. . . . . . .
. .
. . . .
Fig. 17. Horizontal maps of excitatory (EBN) and inhibitory burst neuron projections to the abducens, the vestibular nuclei, and the prepositus. A. EBN projections. B. IBN projections. The density of dots represents the relative density of projections; see Methods.
IBN area in the pontomedullary reticular formation to all of the IBN projection regions found in the present study- the abducens, the prepositus, the dorsomedial reticular for- mation, and all four of the main vestibular nuclei. In con- trast to the present results, a heavier projection was found to the prepositus than to the MVN. In addition, the perihypoglossal projection apparently extended cau- dally to include the nucleus intercalatus, which was not found to receive an IBN projection in the present study. Other projections from this region, ipsilaterally and further rostrally in the pons and midbrain, indicate the presence of other classes of cells interspersed with the IBNs.
A study in the cat that used intracellular staining of behaviorally identified IBNs (Yoshida et al., '82) illustrated an axonal trajectory and branching pattern similar to that found in the present study. Terminations were demon- strated in the abducens, the prepositus, the MVN, and the reticular formation rostral and caudal to the abducens. Hikosaka et al. ('80), using microstimulation and anti- dromic identification, also showed axonal branching of IBNs in these regions, as well as in the rostral DVN and the
ventral LVN. With the exception of the apparent absence of a projection to the SVN in the cat, these results indicate a nearly complete correspondence with the IBN projections in the squirrel monkey.
The significance of burst neuron projections to the abdu- cens is relatively clear, because the two neuronal popula- tions contained within the nucleus are well defined anatomically and physiologically. IBNs inhibit both the motoneurons and the internuclear neurons (Hikosaka et al., '78b, '80) and are thus largely responsible for the pause shown by these neurons during contralateral saccades. Al- though it has not been demonstrated directly, it is likely that EBNs also contact both populations of abducens neu- rons and contribute greatly to their maximal firing during ipsilateral saccades.
The present results provide no evidence concerning the distribution of EBN or IEiN synapses on their target neu- rons. However, it is of interest to note that boutons contain- ing flattened synaptic vesicles have a predominantly somatic distribution on abducens motoneurons, whereas spherical vesicles are distributed fairly evenly over both

I
I A. EBNS I
6. I B N S C.
I
I
. . . .
. . , .
I . . . . . .
I
I '. I ' , . . . .
. . .
. . . .
. .
I I I
! ' I I , . . . , . . . . . . , . \.:
I
I .
I
I
I
I
I , 1
. . I . . . '
1 : I . ,
1 . , I . 1 . . . I
IA I
I
1 , I I I . I
I . . I I I I I I
Fig. 18. Horizontal maps of EBN and IBN projections to reticular forma- tion, shown with a map of burst neuron soma locations. A. EBN projections. B. IBN projections. C. Soma locations. Squares and open triangles in C represent EBNs and IBNs, respectively; the filled triangle represents an unusually caudally located contralaterally projecting burst neuron de-
the soma and proximal dendrites (Spencer and Sterling, '77). This raises the possibility of a differential distribution of IBN and EBN synapses on the soma and proximal den- drites, respectively, of their target neurons in abducens.
Projections to the reticular formation Burst neuron terminations in the reticular formation are
particularly difficult to interpret, because of the intermin- gling in this region of a variety of anatomical and physio- logical classes of neurons. It seems likely that a major target of the more caudal burst neuron terminations in Figure 18A and B is the IBN population. IBNs appear to be the major class of saccade-related neurons found in this region, although other neurons, which are excited by con-
scribed in the text. Borders of the abducens and the prepositus are indicated by dashed lines for rostrocaudal and medio-lateral landmarks; however, the terminations and somas represented in this figure are located in the reticu- lar formation, ventral to these nuclei.
tralateral head rotation (type I1 response) and are active during both phases of nystagmus, are also found here (Hi- kosaka et al., '78a). For both classes of neurons, excitation from ipsilateral EBNs would contribute to the burst during ipsilateral saccades, and inhibition by contralateral IBNs would contribute to the silence or reduced firing frequency during contralateral saccades.
Several classes of saccade-related neurons are found in the region of the reticular formation receiving the more rostral burst neuron projections in Figure 18A and B (Mark- ham et al., '81). The remarkable degree of overlap indicated in Figure 18B and C between the rostral IBN termination region and the distribution of EBN somas suggests EBNs may be a primary target of this projection. Such a projection

IMIIBITORY BURST NEURONS 377
from contralateral IBNs would tend to reduce or abolish EBN discharge during contralateral saccades and might contribute to the negative correlation found in over half the EBNs between number of spikes and saccade amplitude in the off-direction.
The terminations of EBNs in the reticular formation ros- tral to the abducens (Fig. 18A) may represent a self-excita- tory, positive feedback to other EBNs that could serve to produce a more rapid buildup of EBN discharge, resulting in greater eye acceleration at the beginning of the saccade. However, EBN arborizations in this region never enter the neuron's own dendritic field, indicating that these neurons are not directly self-excitatory. Also, the termination region of EBNs (Fig. 18A) has considerably less overlap than that of IBNs (Fig. 18B) with the region containing the EBN somas (Fig. 18C), which suggests that the EBN population is not the major target of these projections.
These rostral EBN projections may be part of an inhibi- tory feedback pathway to omnipause neurons that has been hypothesized by van Gisbergen et al. ('81). Omnipause neu- rons are a class of midline neurons that pause during all saccades and otherwise fire at a steady frequency (Keller, '77; Markham et al., '81; Evinger et al., '82). Omnipause neurons inhibit burst neurons (Nakao et al., '80; Furuya and Markham, '82), and so their saccadic pause is critical for releasing burst neurons from inhibition and allowing the execution of the saccade. Van Gisbergen et al. ('81) have proposed that an inhibitory '%rigger" input to omnipause neurons initiates the pause and disinhibits the EBNs, and that the EBNs in turn feed back on the omnipause neurons via an inhibitory "latch" neuron, which maintains the pause until the saccade is completed. Thus, one of the targets of the EBN terminations in the reticular formation found in the present study may be the hypothetical latch neurons.
Fuchs et al. ('85) have suggested that this inhibitory feed- back to omnipause neurons is provided by the IBNs. This suggestion is based partly on the results of Langer and Kaneko ('84)) who found labelled neurons in the pontine and medullary reticular formation following an injection of HRP in the omnipause neuron region. However, Nakao et al. ('80) found that IBNs could not be antidromically acti- vated from the omnipause region, and other studies of phys- iologically identified IBNs have also failed to find a projection to this region (Hikosaka et al., '80; Yoshida et al., '82).
The present results confirm the finding of Nakao et al. ('80) that IBNs do not project to the omnipause neuron region, which is the midline region at the level of, and rostral to the abducens (Keller,'77; Nakao et al., '80; Mark- ham et al., '81; unpublished observations on intracellularly stained omnipause neurons in the squirrel monkey). This seems to exclude a primary role for IBNs in inhibiting omnipause neurons during the saccade. IBN terminations in the medial part of the pontine reticular formation (Figs. 14, top section, 16A,B,C,18B) could potentially contact lat- erally extending dendrites of omnipause neurons, although terminations in this region are also found for EBNs (Figs. 18A, 15, 17A,B of Strassman et al., '86) and so must be considered weak evidence for an input to omnipause neu- rons. It thus appears that the inhibitory input to omnipause neurons during the saccade arises from a different popula- tion of burst neurons than those that project to the abdu- cens. Such neurons could be present in the same regions that contain the EBNs and IBNs, and display a similar firing pattern; the failure to encounter them in the present
study could be explained by the preferential sampling of neurons with axons that course through the abducens, where most of the electrode tracks were made (see Meth- ods). Alternatively, the latch input could be relayed to the omnipause neurons by the same inhibitory neurons that relay the longer-lead trigger input; neurons with a long- lead firing pattern are present in the EBN and IBN regions, as well as in more rostral portions of the pontine reticular formation.
Projections to the vestibular nuclei and the nucleus prepositus
Burst neurons have major projections to the vestibular nuclei and the propositus, where a large proportion of the neurons, and possibly most of the neurons projecting to the extraocular motor nuclei, have a modulation in their dis- charge during saccades and quick phases of nystagmus (Lopez-Barneo et a1.,'81, '82; Baker et al., '75; Lisberger and Miles, '80; Hikosaka et al., '77, '78a; McCrea et al., '80, '86a,b; Tomlinson and Robinson, '84). Both IBNs and EBNs project to horizontal type I1 neurons in the MVN (Hikosaka et al., '80; Sasaki and Shimazu, %1), which typically have an ipsilateral on-direction in their saccadic modulation, as well as a steady discharge proportional to ipsilateral eye position (Lisberger and Miles, '80). (Type I and I1 neurons have increased discharge during ipsilateral and contralat- eral head rotation, respectively; Duensing and Schaefer, '58.) Type I1 neurons have inhibitory projections to ipsilat- eral type I neurons in the MVN (Sasaki and Shimazu, '81; Nakao et al., '82), which typically show the reverse pattern of saccadic modulation (contralateral burst, ipsilateral pause). Thus EBNs and IBNs can contribute to the saccadic modulation in both classes of horizontal vestibular neurons by monosynaptic excitation and inhibition (respectively) of type 11 neurons and by disynaptic inhibition and disinhibi- tion of type I neurons. Since vestibular and prepositus neu- rons project to the abducens, burst neuron input to these neurons represents a potential indirect pathway by which burst neurons can influence motoneuron discharge during saccades, in addition to their direct abducens projection.
IBN projections to the vestibular nuclei also have a poten- tial role in generating a second major pattern of saccadic modulation found in vestibular neurons, that of a pause in firing for saccades in all directions (Fuchs and Kimm, '75; King et al., '76; Pola and Robinson, '78; Tomlinson and Robinson, '84; these vestibular neurons differ from reticu- lar omnipause neurons in also having a modulation related to head velocity or eye position). This pattern of discharge is found in horizontal vestibular neurons in the MVN and the ventral LVN, as well as in vertical vestibular neurons in each of the four main vestibular nuclei (Tomlinson and Robinson, '84; McCrea et al., '86a,b). Although all of the IBNs have a clear directional specificity in their discharge, many of the neurons have a substantial burst for saccades in all directions and thus might be expected to produce a pause during all saccades in any target neuron that did not also receive an EBN input (compare Fig. 17A,B). In addi- tion, the close correspondence between burst duration and saccade duration usually holds for saccades in all direction, and thus the postsynaptic neurons might be expected to pause equally for all saccades.
Integration of the saccadic signal The postsaccadic firing level in the motoneurons must be
precisely matched with the eye position reached at the end of the saccade (initial position plus saccade amplitude) to

378 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
avoid postsaccadic drift and resultant blurring of vision. It has been proposed that the same neural signal (burst neu- ron discharge) that determines the amplitude of the move- ment is also passed through a neural integrator to construct the signal required for holding the eye in its new position after the movement (Robinson, '71, '75). The existence of such a neural integrator is apparent during the vestibulo- ocular reflex FOR), when the head velocity signal carried by the vestibular nerve is transformed into an eye-position signal on the motoneuron (e.g., Skavenski and Robinson, '73; Shinoda and Yoshida, '74). Theoretical considerations of the alternating slow and quick phases of vestibular nys- tagmus, as well as evidence from unit recording and lesion studies, suggest that the saccadic system uses the same integrator (reviewed in Robinson, '75); however, the exis- tence of separate integrators for the vestibular and saccadic systems has been suggested on the basis of differential sensitivities to pharmacolgical manipulation (Godaux and Laune, '83).
In order to receive a direct burst neuron input, the hori- zontal saccadic integrator must be at least partly contained within the burst neurons' projection regions. Among these regions, the prepositus and the vestibular nuclei (the MVN and the ventral LVN) provide the major eye-position-re- lated input to the abducens. However, the present study indicates that most regions of the vestibular nuclei and the prepositus receive a major projection from only one of the two burst neuron populations, whereas the integrator re- quires both an inhibitory and excitatory saccadic input to have a bidirectional modulation. Neurons in the vestibular nuclei that receive a primary saccadic input from the IBNs alone might be expected to pause for all saccades, which would exclude them from playing a role in the integrator (Tomlinson and Robinson, '84). On the other hand, neurons in the prepositus display a spectrum of eye-position- and eye-velocity-related firing patterns that seem ideally suited to playing a major role in the integrator (Baker et al., '75, '81; Lopez-Barneo et al., '82; cf. Tomlinson and Robin- son, '84). The prepositus receives a heavy EBN projection but only a sparse IBN projection, and so would seem to require an additional inhibitory input during contralateral saccades; this could perhaps be supplied by an inhibitory projection from prepositus or MVN (type ID neurons on the contralateral side. In any case, the present results may be incompatible with models postulating a symmetrical EBN- IBN input to the integrator (van Gisbergen et al., '81).
EBNs and IBNs with oblique on-directions may also pro- vide part of the input to the vertical saccadic integrator by projecting to vertical premotor neurons in the nucleus pre- projecting to vertical premotor neurons in the nucleus pre- positus and the vestibular nuclei, respectively. Although vestibular neurons that pause for all saccades cannot con- tribute to the integrator, vertical premotor neurons with a saccadic firing are also present in the vestibular nuclei (Tomlinson and Robinson, '84; McCrea et al., '86a) and may receive an excitatory saccadic input from vertically related neurons in other brainstem regions (e.g., the nucleus pre- positus (Lopez-Barneo et al., '82) and interstitial nucleus of Cajal (King et al., '81)).
Afferent projections to burst neurons IBNs receive a polysynaptic, predominantly ipsilateral
input from the vestibular nerve (Hikosaka and Kawakami, '77). In addition, projections from the vestibular nuclei to this region have been demonstrated with anterograde trac-
ing techniques in the cat (Ladpli and Brodal, '68; Carleton and Carpenter, '83), as well as by intracellular staining in the alert squirrel monkey (McCrea et al., '86a,b). The pres- ent finding of IBN and EBN dendrites in some cases extend- ing into the abducens nucleus suggests an additional potential pathway for vestibular input to burst neurons. These vestibular pathways may be involved in the activa- tion of burst neurons during the quick phases of vestibular nystagmus, although little is known about the way sus- tained vestibular stimulation results in a biphasic pattern of burst neuron firing.
The only experimentally verified input to burst neurons from a physiologically identified cell type is an inhibitory projection to IBNs from omnipause neurons (Nakao et al., '80; Furuya and Markham, '82). Anatomical studies sug- gest the EBN region also receives a projection from omni pause neurons Ganger and Kaneko, '83), and it is likely that these cells are responsible for suppressing discharge of all burst neurons during fixation.
The superior colliculus appears to play a critical role in generating saccades. The pontomedullary reticular forma- tion receives a projection from the intermediate and deep layers of the contralateral superior colliculus (Harting, '77; Kawamura et al., '74), including a projection from saccadic burst neurons in the colliculus (Keller, '79). However, there is little evidence regarding the way the colliculus's spa- tially organized saccadic command is transformed into the temporally precise pattern of firing exhibited by EBNs and IBNs. It is unlikely that this occurs through a direct colli. cular input to these neurons, and it has been suggested that long-lead burst neurons in the reticular formation play an intermediate role in this transformation (Hepp and Henn, '83; Fuchs et al., '85). A detailed study of the projections of' physiologically characterized long-lead burst neurons could provide critical information on the immediate saccadic in. put to EBNs and IBNs; identification of this input is essen- tial for a further understanding of saccadic organization.
ACKNOWLEDGMENTS The authors would like to thank Herschel Goldstein for
his generous assistance with the computer programming. This study was supported by National Institutes of Health grant EY05433, and was completed in partial fulfillment of A. Strassman's doctoral requirements at the Albert Ein- stein College of Medicine.
LITERATURE CITED
Baker, R., M. Gresty, and A. Berthoz (1975) Neui-onal activity in the prepos itus hypoglossi nucleus correlated with vertical and horizontal eye movement in the cat. Brain Res. 102:366-371.
Baker, R., C. Evinger, and R.A. McCrea (1981) Some thoughts about the three neurons in the vestibular ocular reflex. Ann. N.Y. Acad. Sci. 374t171-188.
Carleton, S.C., and M.B. Carpenter (1983) Merent and efferent connections, of the medial, inferior and lateral vestibular nuclei in the cat and monkey. Brain Res. 27829-51.
Cohen, B., and V. Henn (1972) Unit activity in the pontine reticular forrna- tion associated with eye movements. Brain Res. 46:403-410.
Duensing, F., and K:P. Schaefer (1958) Die Aktivitat einzelner Neurone im Bereich der Vestibulariskerne hei Horizontalheschleun-igungen unter Besunderer beriicksichtigung des vestibularen Nystagmus. Arch. Psy- chiat. Nervenkr. 198:225-252.

INHIBITORY BURST NEURONS 379
Keller, E.L. (1977) Control of saccadic eye movements by midline brainstem neurons. In R. Baker and A. Berthoz (eds): Control of Gaze by Brain Stem Neurons, Developments in Neuroscience, Vol. 1. New York: Elsev- iermorth-Holland, pp. 291-301.
Keller, E.L. (1979) Colliculoreticular organization in the oculomotor system. In R. Granit and 0. Pompeiano (eds): Reflex Control of Posture and Movement. Amsterdam: Elsevier, Prog. Brain Res. 50:725-734.
King, W.M., S.G. Lisberger, and A.F. Fuchs (1976) Responses of fibers in medial longitudinal fasciculus (MLF) of alert monkeys during horizontal and vertical conjugate eye movements evoked by vestibular or visual stimuli. J. Neurophysiol. 39:1135-1149.
King, W.M., A.F. Fuchs, and M. Magnin (1981) Vertical eye movement- related responses of neurons in midbrain near interstitial nucleus of Cajal. J. Neurophysiol. 46:549-562.
Ladpli, R., and A. Brodal (1968) Experimental studies of commissural and reticular formation projections from the vestibular nuclei in the cat. Brain Res. 8t65-96.
Langer, T.P., and C.R.S. Kaneko (1983) Efferent projections of the cat ocu- lomotor reticular omnipause neuron region: An autoradiographic study. J. Comp. Neurol. 217:288-306.
Langer, TP., and C.R.S. Kaneko (1984) Brainstem afferents to the omni- pause region in the cat: A horseradish peroxidase study. J. Comp. Neu- rol. 230:444-458.
Leontovich, T.A., and G.P. Zhukova (1963) The specificity of the neuronal structure and topography of the reticular formation in the brain and spinal cord of carnivora. J. Comp. Neurol. 121:347-379.
Lisberger, S.G., and F.A. Miles (1980) Role of primate medial vestibular nucleus in long-term adaptive plasticity of vestibuloocular reflex. J. Neurophysiol .43: 1725- 1745.
Lopez-Barneo, J., J. Ribas, and J.M. Delgado-Garcia (1981) Identification of prepositus neurons projecting to the oculomotor nucleus in the alert cat. Brain Res. 214:174-179.
Lopez-Barneo, J., C. Darlot, A. Berthoz, and R. Baker (1982) Neuronal activity in prepositus nucleus correlated with eye movement in the alert cat. J. Neurophysiol. 47t329-352.
Luschei, E.S., and A.F. Fuchs (1972) Activity of brain stem neurons during eye movements of alert monkeys. J. Neurophysiol. 35t445-461.
Markham, C.H., S. Nakao, and I.S. Curthoys (1981) Cat medial pontine neurons in vestibular nystagmus. Ann. N.Y. Acad. Sci. 374t189-209.
McCrea, R.A., K. Yoshida, A. Berthoz, and R. Baker (1980) Eye movement related activity and morphology of second order vestibular neurons terminating in the cat abducens nucleus. Exp. Brain Res. 4Ot468-473.
McCrea, R.A., A. Strassman, and S.M. Highstein (1986a) Anatomical and physiological characteristics of vestibular neurons mediating the verti- cal vestibuloocular reflexes in the squirrel monkey. J. Comp. Neurol. (submitted).
McCrea, R.A., A. Strassman, E. May, and S.M. Highstein (1986b) Anatomi- cal and physiological characteristics of vestibular neurons mediating the horizontal vestibuloocular reflex in the squirrel monkey. J. Comp. Neurol. (submitted).
Nakao, S., I.S. Curthoys, and C.H. Markham (1980) Direct inhibitory projec- tion of pause neurons to nystagmus-related pontomedullary reticular burst neurons in the cat. Exp. Brain Res. 40~283-293.
Nakao, S., S. Sasaki, R.H. Schor, and H. Shimazu (1982) Functional organi- zation of premotor neurons in the cat medial vestibular nucleus related to slow and fast phases of nystagmus. Exp. Brain Res. 45:371-385.
Olszewski, J,, and D. Baxter (1954) Cytoarchitecture of the Human Brain Stem. Basel: J.B. Lippincott Co.
Pola, J., and D.A. Robinson (1978) Oculomotor signals in medial longitudi- nal fasciculus of the monkey. J. Neurophysiol. 41.245-259.
Ramon-Moliner, E., and W.J.H. Nauta (1966) The isodendritic core of the brain stem. J. Comp. Neurol. 126:311-336.
Robinson, D.A. (1970) Oculomotor unit behavior in the monkey. J. Neuro- physiol. 33t393-404.
Robinson, D.A. (1971) Models of oculomotor neural organization. In P. Bach- Y-Rita, C.C. Collins, and J.E. Hyde (eds): The Control of Eye Move- ments. New York: Academic Press, pp. 519-538.
Robinson, D.A. (1975) Oculomotor control signals. In P. Bach-Y-Rita and G. Lennerstrand (eds): Basic Mechanisms of Oculomotor Motility and Their Clinical Implications. Oxford: Pergamon Press, pp. 337-378.
Sasaki, S., and H. Shimazu (1981) Reticulovestibular organization partici- pating in generation of horizontal fast eye movement. Ann. N.Y. Acad. Sci. 374:130-143.
%hiller, P.H. (1970) The discharge characteristics of single units in the
Evinger, C., C.R.S. Kaneko, and A.F. Fuchs (1982) The activity of omni- pause neurons in alert cats during saccadic eye movements and visual stimuli. J. Neurophysiol. 47:827-844,
Fuchs, A.F., and J. Kimm (1975) Unit activity in vestibular nucleus of the alert monkey during horizontal angular acceleration and eye move- ment. J. Neurophysiol. 38:1140-1161.
Fuchs, A.F., and E.S. Luschei (1970) Firing patterns of abducens neurons of alert monkeys in relationship to horizontal eye movement. J. Neurophy- siol. 33:382-392.
Fuchs, A.F., C.R.S. Kaneko, and C.A. Scudder (1985) Brainstem control of saccadic eye movements. Annu. Rev. Neurosci. 8:307-337.
Furuya, N., and C.H. Markham (1982) Direct inhibitory synaptic linkage of pause neurons with burst inhibitory neurons. Brain Res. 245139-143.
Godaux, E., and J.-M. Laune (1983) The saccadic system and the vestibulo- ocular reflex in the cat do not share the same integrator. Neurosci. Lett. 38:263-268.
Goldstein, H.P. (1983) The Neural Encoding of Saccades in the Rhesus Monkey. Ph.D. Thesis, Johns Hopkins University.
Graf, W., R.A. McCrea, and R. Baker (1983) Morphology of posterior canal related secondary vestibular neurons in rabbit and cat. Exp. Brain Res. 52125-138.
Graybiel, A.M. (1977) Direct and indirect preoculomotor pathways of the brainstem: An autoradiographic study of the pontine reticular forma- tion in the cat. J. Comp. Neurol. 175:37-78.
Graybiel, A.M., and E.A. Hartweig (1974) Some afferent connections of the oculomotor complex in the cat: An experimental study with tracer tech- niques. Brain Res. 81:543-551.
Harting, J.K. (1977) Descending pathways from the superior colliculus: An autoradiographic analysis in the rhesus monkey (Macaca mulatta). J. Comp. Neurol. 173583-612.
Henn, V., and B. Cohen (1973) Quantitative analysis of activity in eye muscle motoneurons during saccadic eye movements and positions of fixation. J. Neurophysiol. 36:115-126.
Henn, V., and B. Cohen (1976) Coding of information about rapid eye movements in the pontine reticular formation of alert monkeys. Brain Res. 108:307-325.
Hepp, K., and V. Henn (1983) Spatio-temporal recoding of rapid eye move- ment signals in the monkey paramedian pontine reticular formation (PPRF). Exp. Brain Res. 52105-120.
Highstein, S. M., K. Maekawa, A. Steinaker, and B. Cohen (1976) Synaptic input from the pontine reticular nuclei to abducens motoneurons and internuclear neurons in the cat. Brain Res. 112:162-167.
Hikosaka, O., and T. Kawakami (1977) Inhibitory reticular neurons related to the quick phase of vestibular nystagmus-their location and projec- tion. Exp. Brain Res. 27t377-396.
Hikosaka, O., M. Maeda, S. Nakao, H. Sbimazu, and Y. Sbinoda (1977) Presynaptic impulses in the abducens nucleus and their relation to postsynaptic potentials in motoneurons during vestibular nystagmus. Exu. Brain Res. 27:355-376.
Hikosaka, O., Y. Igusa, and H. Imai (1978a) Firing pattern of prepositus hypoglossi and adjacent reticular neurons related to vestibular nystag- mus in the cat. Brain Res. 144t395-403.
Hikosaka, O., Y. Igusa, S. Nakao, and H. Shimazu (1978b) Direct inhibitory synaptic linkage of pontomedullary reticular burst neurons with abdu- cens motoneurons in the cat. Exp. Brain Res. 33t337-352.
Hikosaka, O., Y. Igusa, and H. Imai (1980) Inhibitory connections of nystag- mus-related reticutar burst neurons with neurons in the abducens, pre- positus hypoglossi and vestibular nuclei in the cat. Exp. Brain Res. 39t301-311.
Igusa, Y., S. Sasaki, and H. Shimazu (1980) Excitatory premotor burst neurons in the cat pontine reticular formation related to the quick phase of vestibular nystagmus. Brain Res. 182t451-456.
Kaneko, C.R.S., and A.F. Fuchs (1981) Inhibitory burst neurons in alert trained cats: Comparison with excitatory burst neurons and functional implications. In A.F. Fuchs and W. Becker (eds): Progress in Oculomotor Research, Developments in Neuroscience, V01.12.
Kaneko, C.R.S., C. Evinger, and A.F. Fuchs (1981) Role of cat pontine burst neurons in generation of saccadic eye movements. J. Neurophysiol. 46t387-408.
Kawamura, K., A. Brodal, and G.H. Hoddevik (1974) The projection of the superior colliculus onto the reticular formation of the brain stem. An experimental anatomical study in the cat. Exp. Brain Res. 19:l-19.
Keller, E.L. (1974) Participation of the medial pontine reticular formation in eye movement generation in monkey. J. Neurophysiol. 37:316-332.

380 A. STRASSMAN, S.M. HIGHSTEIN, AND R.A. McCREA
oculomotor and abducens nuclei of the unanesthetized monkey. Exp. Brain Res. IO:347-362.
Shinoda, Y., and K. Yoshida (1974) Dynamic characteristics of responses to horizontal head angular acceleration in vestbuloocular pathway in the cat. J. Neurophysiol. 37:653-673.
Skavenski, A.A., and D.A. Robinson (1973) Role of abducens neurons in vestibuloocular reflex. J. Neurophysiol. 36324-738.
Spencer, R.F., and P. Sterling (1977) An electron microscope study of moto- neurons and interneurons in the cat abducens nucleus identified by retrograde intraaxonal transport of horseradish peroxidase. J. Comp. Neurol. 176:65-85.
Steiger, H.-J., and J.A. Biittner-Ennever (1979) Oculomotor nucleus affer- ents in the monkey demonstrated with horseradish peroxidase. Brain Res. 16O:l-15.
Strassman, A,, S.M. Highstein, and R.A. McCrea (1986) Anatomy and phys- iology of saccadic burst neurons in the alert squirrel monkey. I. Excita- tory burst neurons. J. Comp. Neurol. 249t337-357.
Taber, E. (1961) The cytoarchitecture of the brain stem of the cat I. Brain stem nuclei of the cat. J. Comp. Neurol. 116:27-69.
Tomlinson, R.D., and D.A. Robinson (1984) Signals in vestibular nucleus mediating vertical eye movements in the monkey. J. Neurophysiol. 51:1121-1136.
Uchino, Y., N. Hirai, S. Suzuki, and S. Watanabe (1981) Properties of secondary vestibular neurons fired by stimulation of ampullary nerve of the vertical, anterior or posterior, semicircular canals in the cat. Brain Res. 223:273-286.
Uchino, Y., N. Hirai, and S. Suzuki (1982) Branching pattern and properties of vertical- and horizontal-related excitatory vestibuloocular neurons in the cat. J. Neurophysiol. 48.391-903.
Valverde, F. (1961) Reticular formation of the pons and medulla oblongata. A Golgi st,udy. J. Comp. Neurol. 116:71-99.
van Gisbergen, J.A.M., D.A. Robinson, and S. Gielen (1981) A quantitative analysis of generation of saccadic eye movements by burst neurons. J. Neurophysiol. 45t417-442.
Yoshida, K., R.A. McCrea, A. Berthoz, and P.P. Vidal(1982) Morphological and physiological characteristics of inhibitory burst neurons controlling horizontal rapid eye movements in the alert cat. J. Neurophysiol. 48%- 784.