análisis fisicoquímico y biológico comparado en dos...
TRANSCRIPT

Revista de Investigaciones No. 17 - Universidad del Quindío
57
Revista de Investigaciones No. 17 - Universidad del Quindío p p 57- 80 Armenia, Año 2007 ISSN 1794-631 X
Análisis Fisicoquímico y Biológico
Comparado en Dos Quebradas
de Alta Montaña NeotropicalCarlos A. García-Alzate1, César Román-Valencia2, James A. Vanegas-Ríos2 y Dahiana K. Arcila-Mesa2.
Universidad del Quindío, Programa Académico de Biología, Laboratorio de Ictiología, A. A. 2639, Armenia, Quindío, Colombia.
RESUMEN
Se analizaron características físicas, químicas y biológicas (fitoplancton, zooplancton, macroinvertebrados y peces) de las quebradas Doña Juana y Villa Paola afluentes de tipo secundario del río Quindío, Alto Cauca. Se encontró que el oxígeno y el porcentaje de saturación fueron altos. La quebrada Doña Juana mostró un coeficiente de variación bajo en pH, conductividad, y durezas (cálcicas, magnésicas y totales); la acidez fue alta en lluvias, mientras los sólidos altos en sequía. La quebrada Villa Paola mostró valores altos de sólidos, durezas (cálcicas, magnésicas y totales) y Demanda Química de Oxigeno (DQO). El déficit de oxígeno cambió fuerte y negativamente en la quebrada Villa Paola (coeficiente de variación, C. V., dos veces más altos al comparar con la quebrada Doña Juana). Se registraron cuatro clases y 10 órdenes de macroinvertebrados, cinco divisiones, 14 familias y 15 géneros de fitoplancton, y seis clases, 11 familias y 13 géneros de zooplancton, dos órdenes, tres familias y cinco especies de peces; los que difieren en presencia, frecuencia y abundancia entre quebradas. Estos drenajes se comportan como ecosistemas oligotróficos de alta montaña neotropical. Palabras Clave: Quebradas, limnología, Colombia, Quindío, Neotrópico.
ABSTRACT
A limnological analysis (phytoplankton, zooplankton, macroinvertebrates and fishes) from Doña Juana and Villa Paola creeks, secondary tributaries of Quindío river, Alto Cauca was studies. The results of the physic-chemical variables showed that the oxygen is high. The Doña Juana creek showed a low variation coefficient pH, conductivity, and hardness (calcium, magnesium and total); the acidity was high in rains and solids in dry season. The Villa Paola creek showed high values of solids, hardness (calcium, magnesium and total) and Chemical Demand of Oxygenate. The deficit of oxygen had a change negatively strong in the Villa Paola creek (variation coefficient C. V. twice higher than the Doña Juana creeks). Four classes and 10 macroinvertebrates orders, five divisions, 14 families and 15 phytoplankton genera and six classes, 11 families and 13 zooplankton genera, and two orders, three families and five species of fish were registered; with differences between creeks. These drainages are ecosystems oligotrophic of Neotropical high mountain Andean.
Key words: Creeks, limnology, Colombia, Quindío, Neotropical.Critical period hypothesis, mother tongue, second language, bilingualism
1 Correo electrónico: [email protected] Correo electrónico: [email protected]

Revista de Investigaciones No. 17 - Universidad del Quindío
58
INTRODUCCIÓN
os análisis limnológicos se orientan a determinar características fisicoquímicas del agua y de las
comunidades asociadas. Se parte del principio de que a cada tipo de ecosistema acuático esta asociada una comunidad particular de organismos (Roldán, 1992, 1996; Lampert y Sommer, 1997; Wetzel y Likens, 2000). Los aspectos biológicos han adquirido una creciente importancia en el estudio de los ecosistemas acuáticos, porque las variables fisicoquímicas dan una idea puntual sobre
la calidad del agua, pero no informan sobre las variaciones en el tiempo (Alba-Tercedor, 1996; en Caicedo y Palacio, 1998), ya que las características de las comunidades acuáticas (por ejemplo macroinvertebrados, peces, fitoplancton y zooplancton) actúan como
testigos del deterioro ambiental de las corrientes superficiales (Caicedo y Palacio, 1998). En el
neotrópico existe una apreciable degradación por procesos agropecuarios e industriales y por el vertido de aguas negras domésticas (provenientes de sistemas de tratamiento o en su defecto de la intervención antrópica) sobre los drenajes naturales, lo que causa variaciones drásticas en sus condiciones fisicoquímicas y
biológicas (Roldán, 1992; Román-Valencia y Muñoz, 2001; Román-Valencia et al., 2003; Román-Valencia y Perdomo, 2004), Además, otro factor que modifica los ambientes acuáticos
y terrestres es el turismo. El aumento de su transformación posee consecuencias negativas para los peces y la biodiversidad ribereña en general (Palacio–Núñez, et al. 2007).La mayoría de las investigaciones que señalan aspectos físico-químicos y biológicos en la cuenca media del Cauca (lagos, represas, ríos y quebradas) se han desarrollado por: Machado y Roldán (1981); Correa et al. (1981); Matthias y Moreno (1983); Caicedo y Palacio (1998);
Sierra et al. (2004); Urrego y Ramírez (2000); Arboleda y Ramírez (2002); Ramírez y Alcaraz (2002); Gallo-Sánchez et al. (2004); Hernández-Atilano et al. (2005). También existen algunos trabajos sobre bioindicación con macroinvertebrados (Rincón, 1996, 2002; Medellín et al., 2004). Para la cuenca alta del río Cauca se resaltan los trabajos hechos por Cardona et al.(1998); Román-Valencia y Muñoz (2001); Román-Valencia et al. (2003); Román-Valencia y Perdomo (2004); Román-Valencia y Ruiz (2005); Román-Valencia et al. (2005). En ésta perspectiva, el objetivo de éste artículo es analizar y comparar la estructura fisicoquímica
y biológica en dos drenajes del alto Cauca.
MATERIALES Y MÉTODOS
Zona de estudio. El trabajo se realizó en las quebradas Villa Paola y Doña Juana, afluentes
del río Quindío, cuenca del Río La Vieja, alto Cauca (Colombia) entre los 1758 (4° 27’ 4” N y 75° 44’ 12” O) y 1734 (4° 37’ 51” N y los 75° 36’ 10” O) msnm respectivamente. La quebrada Villa Paola se caracteriza por presentar vegetación de Bromeliáceas, Melastomatáceas, Aráceas, Musáceas, Pináceas (Pinus), Mirtáceas (Eucalyptus) y Gramíneas (en especial Guadua angustifolia). Sus aguas presentan color cristalino y marrón, con sustrato pedregoso-arenoso. La quebrada Doña Juana se caracteriza por una vegetación de Melastomatáceas, Aráceas, Musáceas, Pináceas (Pinus), Mirtáceas (Eucalyptus), Cupresáceas y Gramíneas (G. angustifolia). Además, presenta sustrato abundante en arena y detrito, con piedras de diferentes tamaños, dando lugar a zonas de corrientes rápidas, lentas y remansos. Ambos drenajes están rodeados de potreros y fincas
agrícolas, son una unidad litológica similar por estar ubicados sobre la formación geológica del abanico del Quindío, comparten sedimentos
L

Revista de Investigaciones No. 17 - Universidad del Quindío
59
volcánicos con depósitos piroclasticos, lahares y acumulaciones fluviales; además de fragmentos
de rocas angulares y subangulares (González y Nuñez, 1991); caracterizados por sedimentitas, es decir aluviones recientes de inicios del cuaternario, influenciados por procesos de
origen endógeno, estos han dado lugar a zonas de fuertes depresiones o inclinaciones topográficas, producto del levantamiento de la
Cordillera Central y la actividad de los sistemas de fallas que recubren la región. Tal es el caso del sistema Silvia-Pijao, San Jerónimo, Navarco y los volcanes presentes en el pie de monte de la cordillera Central en Colombia (Botero et
al., 2005), los cuales han ejercido una gran influencia sobre la dinámica hidrogeológica de
la zona de estudio.
Fase de campo y laboratorio. La toma de datos se llevó a cabo entre el 31 de marzo y 1 de abril (época de sequía) y el 6-7 y 20-21 de mayo de 2006 (época de lluvias). Se obtuvieron los datos pluviométricos (suministrados por el IDEAM) de la estación La Vieja (4° 38’ N y 75° 34’ O) ubicada a 1895 msnm, en el municipio de Salento, departamento del Quindío, Colombia. Las determinaciones de las variables fisicoquímicas se hicieron in situ como sigue: coloración y sustrato por observación directa; oxígeno disuelto, porcentaje de saturación de oxígeno y temperaturas con oxímetro OXI196-Microprocesador; humedad relativa con termohigrometro; pH con potenciómetro PIN POINT-BNC (registradas cada hora); ancho y profundidad con decámetro; velocidad de corriente con cronómetro y una bola de icopor, donde se registró el tiempo que tarda en recorrer esta bola una distancia conocida. La dureza (total, cálcica y magnésica), alcalinidad, demanda química de oxígeno (D. Q. O), demanda bioquímica de oxígeno (D. B. O), cloruros, sólidos (totales, suspendidos y disueltos), alcalinidad y acidez se determinaron
de acuerdo con la metodología recomendada por la APHA (1998) y Wetzel y Likens (2000), en el laboratorio de aguas de la Universidad del Quindío. Además, se utilizaron dos botellas DBO: una oscura y otra sin interferencia de luz (botella clara) para registrar el oxígeno disuelto y el porcentaje de saturación cada hora en el ciclo nictemeral. Se colectaron muestras de agua en botellas de un litro, para registrar el fitoplancton y zooplancton presente. Estas se
fijaron in situ con formol al 4% y se preservaron
con lugol, luego se transportaron al laboratorio de biología de la Universidad del Quindío, donde se determinaron con descripciones y claves de Lackey (1956), Kudo (1966), Bicudo y Bicudo (1970), Uhlerkovich y Schmidt (1974) y Needham y Needham (1978). Además, se muestrearon macroinvertebrados acuáticos con nasa acuática y captura directa. La muestra se determinó in situ; aquellos organismos que necesitaron ser verificados se fijaron
en alcohol al 70 % en frascos debidamente rotulados. Posteriormente, con ayuda de claves y descripciones (Needham y Needham, 1978; Correa et al., 1981; Roldán, 1996; Wetzel y Likens, 2000; Posada-García y Roldán-Pérez, 2003) se determinaron hasta género cuando fue posible. La colecta de peces se realizó con redes de arrastre como aparejo entre dos personas haciendo arrastres a favor de la corriente, en contra y de orilla a orilla tratando de abarcar todo los biotopos de la quebrada: remanso, corriente, o ambos. La determinación se realizó in-situ; los ejemplares que no se pudieron determinar en campo se fijaron en formol al 10% y trasladaron
al laboratorio de Ictiología de la Universidad del Quindío (IUQ) para su posterior determinación con base en claves, descripciones y registros (Román-Valencia 1995, 2003, Román-Valencia y Ruiz-C. 2007, Ruiz-C y Román-Valencia 2006 a, b).
Análisis de datos. Para los datos fisicoquímicos

Revista de Investigaciones No. 17 - Universidad del Quindío
60
obtenidos cada hora, se utilizó una ANDEVA de dos factores (factor uno igual a variación nictemeral o temporal; y factor dos igual a variación entre quebradas o épocas) cuando los registros fueron representativos (mayores a 20). La media aritmética se utilizó como medida de tendencia central. Además, se calculó el coeficiente de variación (C. V.) como medida de
dispersión relativa, acompañada de la amplitud (valor máximo y mínimo). Con los datos pluviométricos sé hizo un análisis multianual mensual. En el caso del oxígeno disuelto de las quebradas y de las botellas DBO se calculó el déficit de oxígeno (D. O. = Oxígeno final-
Oxígeno inicial) tanto para la botella clara como la oscura, para medir producción y consumo del mismo.Se hizo un conteo de macroinvertebrados, fitoplancton y zooplancton. En el caso de los
dos últimos se utilizó la técnica de registro por gotas observadas, lo que permitió organizar los datos en individuos por ml (ind/ml). Los datos biológicos (fitoplancton, zooplancton,
macroinvertebrados) se procesaron para obtener los índices de diversidad alfa y beta. Para macroinvertebrados se calculó el promedio general de la diversidad alfa, al unificar la
información obtenida hasta familia. En cambio para fitoplancton y zooplancton se calcularon la
diversidad por división y el promedio general a partir de los datos de familias, mientras para peces se calcularon índices de diversidad alfa. Los índices calculados fueron los de Shannon-Wiener (H’) en bits/ individuo, dominancia de Simpson (D), equidad de Pielou (E) y riqueza especifica de Margalef (M). En todos
los casos se utilizó el logaritmo en base dos. Además, se calculó el índice de Cody (BC2) para medir la diversidad beta (Humphries et al., 1995) y la abundancia relativa (A. R.). Para comparar los valores arrojados por los índices de diversidad se utilizó una prueba t-student de dos muestras, solo para los datos de diversidad
alfa de fitoplancton y zooplancton. El nivel de
significancia utilizado para las pruebas fue de 5
%. Por conveniencia se reportaron los valores estadísticamente significativos. Todos los
análisis estadísticos se procesaron en el paquete Statistix 7,0 bajo Windows.
RESULTADOS
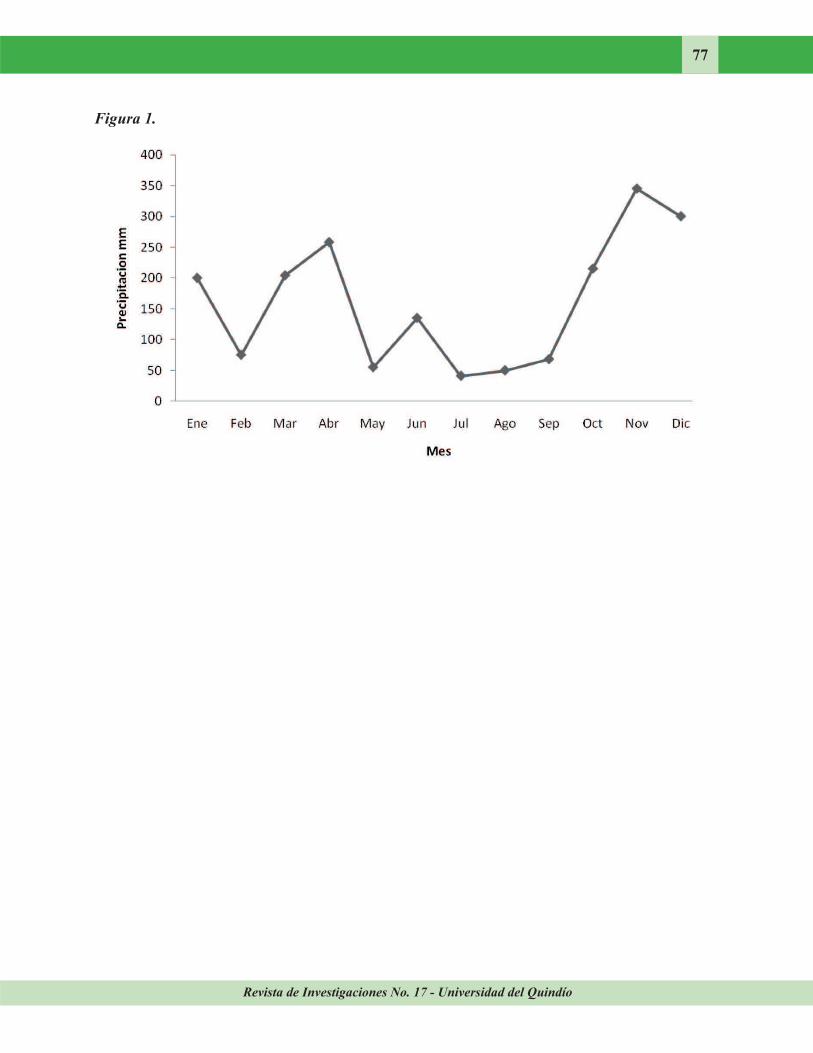
Variables fisicoquímicas. La pluviosidad de la estación La Vieja en el municipio de Salento en 2006 mostro precipitaciones promedio de 189,2 mm (Fig. 1). El análisis mensual multianual de 2006 presentó una tendencia bimodal, con incrementos fuertes a finales de abril y noviembre
(Fig. 1). Es decir, entre febrero y abril se registró una tendencia a aumentar la precipitación (de 75 a 258 mm respectivamente), al igual que entre septiembre y noviembre (de 68 a 354 mm respectivamente). Así los datos de sequía y lluvias del año de estudio se ajustaron al registro multianual de la precipitación pluvial.
Quebrada Doña Juana. Las variables fisicoquímicas, con excepción del déficit de
oxígeno, registraron un bajo coeficiente de
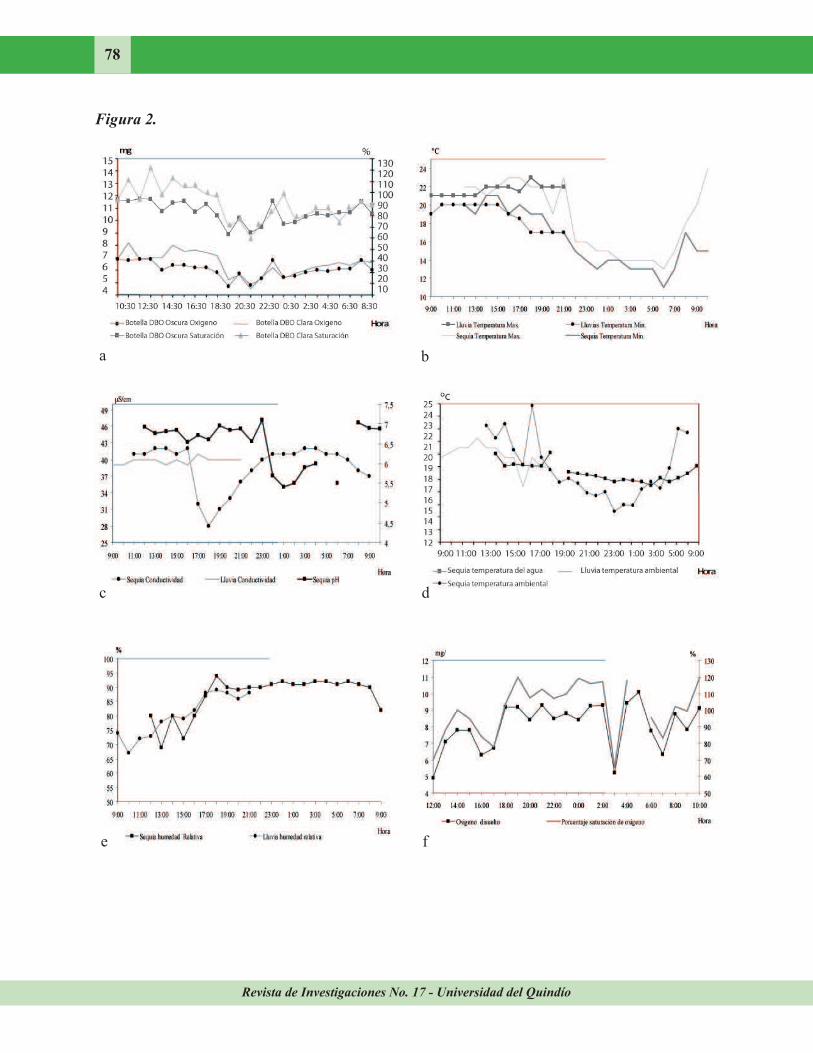
variación (tabla 1). El oxígeno disuelto y el porcentaje de saturación de la botella DBO clara registraron en promedio para sequía 7,6 mg/l y 93,2 % respectivamente, mientras en lluvias se presentó una ligera disminución (7,0 mg/l y 87,6 %). Para la botella DBO oscura se presentó el mismo fenómeno de disminución entre épocas (tabla 1). En la curva de oxígeno disuelto y porcentaje de saturación de las botellas DBO no se observó marcada variación, se obtuvieron resultados similares para los periodos muestreados (Fig. 2a). La temperatura máxima y mínima mostró en ambas épocas una disminución hacia las horas de la noche (Fig. 2b). Según el ANDEVA no existen diferencias significativas en la temperatura máxima

Revista de Investigaciones No. 17 - Universidad del Quindío
61
entre épocas y entre horas (F= 0,727, p= 0,393; F= 8,449, p= 0,995, respectivamente). Sin embargo, en la temperatura mínima se registraron diferencias significativas en la
variación nictemeral (F= 8,894, p= 0,002), aunque entre épocas se observó lo contrario (F= 30,541, p= 0,134). El pH presentó tendencia por debajo de neutro (5,4-7,1) en época seca (Fig. 2c). La conductividad en lluvias (C. V. = 1,9 %) registró poca variación en sequía (C. V.= 10,5) (Fig. 2c), aunque en ambos casos, el promedio bajó (Sequía 38,7 μS/cm; Lluvia 39,8 μS/cm).
La temperatura del agua tuvo en época seca un promedio de 17,3 ºC (C. V.= 6,1 %) y en el período de lluvia fue de 18,2 ºC (C. V.= 3,9 %) (tabla 1). Sin embargo, en sequía se registró una tendencia a disminuir con la llegada de la noche (Fig. 2d). No se observaron diferencias significativas entre los cambios de horas y
épocas (F= 2,130, p= 0,144; F=22,891, p= 0,407). La humedad relativa en ambas épocas tuvo una tendencia a aumentar con la llegada de la noche (Fig. 2e). El ANDEVA no mostró influencia significativa de las épocas (F=
31,442, p=0,087) aunque sí señaló influencia de
las horas (F=12,565, p= 0,004). El porcentaje de saturación (en la época seca) mostró tendencia a aumentar durante la noche (18:00 hr. 104,0 % - 2:00 hr.= 117,0 %) (Fig. 2f). Sin embargo, los coeficientes de variación no son altos (C. V.=
16,4 %; C. V.= 17,3 %, respectivamente). En la época lluviosa el oxígeno disuelto y porcentaje de saturación estuvieron en promedio entre 7,5 mg/l y 91,3 %. Las durezas (totales, suspendidas y disueltas), los cloruros y la D. Q. O. presentaron leves cambios entre épocas (tabla 1). Sin embargo, fueron bajos. La alcalinidad fue más alta en lluvias que en sequía (sequía 22,9 mg/l CaCO3; lluvias 73,8 mg/l CaCO3). La acidez registró igual tendencia (sequía 5,6 mg/l CaCO3; lluvia 64,0 mg/l CaCO3). Los sólidos totales y disueltos, fueron más altos en sequía que en
lluvias (sequía 43 mg/l, lluvia 17 mg/l y 41 en sequía, lluvia 6 mg/l respectivamente) (tabla 1). En general, se observan registros bajos (a excepción de la alcalinidad y la acidez).
Quebrada Villa Paola. A excepción del déficit
de oxígeno, se registraron coeficientes de
variación bajos (tabla 1). El oxígeno disuelto y el porcentaje de saturación de oxígeno de las botella DBO Winklers en sequía obtuvieron poca variación (C. V. < 12 %) (Fig. 3a). Igual ocurrió con las botellas DBO en lluvias (tabla 1) (Fig. 3b). El ANDEVA mostró influencia
significativa entre épocas para el porcentaje de
saturación y el oxígeno disuelto de las botellas DBO (Botella oscura: F= 8,166, p= 0,004; F= 16,666, p= 0,000; Botella clara: F= 24,827, p= 0,359; F= 9,782, p= 0,001, respectivamente). El oxígeno disuelto y el porcentaje de saturación en sequía oscilaron entre 7,5-8,6 mg/l y 79,5-89,8 %, mientras en lluvias lo fue entre 5,6-7,6 mg/l y 76,2-101,2 % (Fig. 3c) (tabla 1). Se encontró influencia significativa de las épocas sobre el
oxígeno disuelto y la saturación de oxígeno (F= 23,000, p= 0,000; F=5,260, p= 0,021). El pH en lluvias estuvo cercano a la neutralidad (6,3), aunque disminuyó con la llegada de la noche (5,8). La conductividad (en lluvias) registró poca variación en el ciclo nictemeral (C. V. = 1,3 %) (Fig. 3d). La temperatura máxima y mínima registró en ambas épocas disminución en la noche (Fig. 3e). Según el ANDEVA sólo en la temperatura máxima se registró diferencia significativa entre épocas (F= 12,565, p= 0,005).
La temperatura del aire (sequía: C. V.= 17,6 %; lluvia: C. V.= 7,4 %) en ambos períodos fue un poco más alta que la temperatura del agua (sequía y lluvias: C. V.= 4,9 %) (Fig. 3f). El ANDEVA mostró diferencias significativas para el factor
época (temperatura superficial: F= 37,964, p=
0,018; temperatura aire: F= 36,362, p= 0,027). Los cloruros (sequía 20,0 mg/l; lluvia 21,1 mg/l) y la alcalinidad (sequía 25,2 mg/l CaCO3;

Revista de Investigaciones No. 17 - Universidad del Quindío
62
lluvia 29,4 mg/l CaCO3) fueron bajos y no presentaron altas diferencias entre épocas (tabla 1). La D. Q. O. fue baja, aunque en sequía (48,1 mg/l O2) fue mayor que en lluvias (28,9 mg/l). La acidez, los sólidos y durezas totales, cálcicas y magnésicas fueron mas altas en lluvias que en sequia (tabla 1).
Comparación de las variables fisicoquímicas entre las quebradas. El ANDEVA mostró diferencias significativas entre quebradas
y durante el ciclo diurno-nocturno para la temperatura del aire (F= 9,095, p= 0,028, F= 51,658, p= 0,000). El oxígeno disuelto de ambas quebradas fue distinto, con tendencia a ser mayor en la quebrada Doña Juana para ambas épocas (tabla 1). Lo contrario ocurrió con la conductividad, que fue más alta en la quebrada Villa Paola con relación a la quebrada a Doña Juana (53,0 y 46,8 μS/cm, respectivamente).
Las durezas de la quebrada Villa Paola fueron más altas que las de la quebrada Doña Juana. En general el déficit de oxígeno de la botella
DBO clara y oscura mostró una fuerte variación en la quebrada Villa Paola (C. V. dos veces más altos que la quebrada Doña Juana). Las demás variables fisicoquímicas no registraron cambios
significativos (tabla 1).
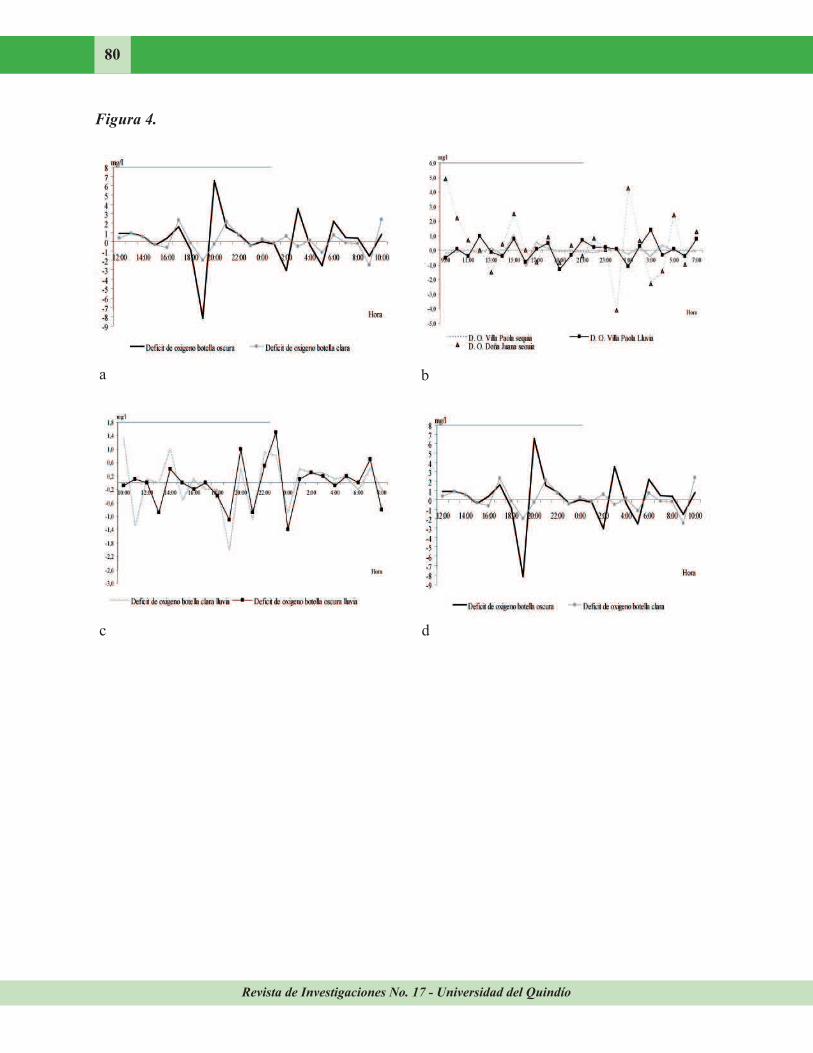
Déficit de oxígeno. En sequía la quebrada Doña Juana presentó variaciones más fuertes en la botella clara (C. V.= 974,9 %) que en el oscuro (C. V.= 828, 8 %) (Fig. 4a). Al comparar el déficit de oxígeno entre las quebradas Doña
Juana (sequía) y Villa Paola (sequía y lluvia) se obtuvo que la primera osciló entre 4,9 y 1,3 mg/l (C. V. = 926,3 %). Sin embargo, la quebrada Villa Paola en ambas épocas registró un C. V. mayor (sequía 3419,8 %; lluvia 2173,7 %) (Fig. 4b). En la quebrada Villa Paola, durante las lluvias se obtuvo un déficit de oxígeno que
osciló en la botella clara y oscura alrededor de los -0,1 mg/l (Fig. 4c). En sequía se observó un
déficit de oxígeno que osciló en botella clara y
oscura alrededor de los –0,1 y 0,1 mg/l (Fig. 4d). Además, en ambas épocas ésta quebrada registró déficit de oxígeno simétrico en cuanto
al comportamiento nictemeral de las Botellas. En general, para las Botellas en las quebradas Doña Juana y Villa Paola se registró tendencia a disminuir el déficit de oxígeno (valores
negativos) y fluctuar con mayor intensidad entre
las 17:00 ó 18:00 hr. (Fig. 4).
Macroinvertebrados. Se encontraron diez órdenes y cuatro clases (tabla 2). En la quebrada Villa Paola se registró mayor abundancia de Hemiptera (Sequía 100 % y Lluvia 21 %) y Diptera (Sequía 0 % y Lluvia= 39,8 %). A su vez las familias predominantes en ambas épocas fueron Naucoridae (47,0 %), Vellidae (8,4%), Simuliidae (8,2 %), Tipulidae (7,0%) y Gerridae (5,3%). En la quebrada Doña Juana los Hemiptera (Sequía 40,8% y Lluvia 36,9%) y Trichoptera (Sequía 16,3 % y Lluvia 23,1 %) fueron los más representativos. Los grupos con mayor abundancia relativa (ambas épocas) fueron Naucoridae (19,5 %), Hydropsychidae (16,6 %), Vellidae (15,3 %), Gerridae (10,2 %) y Ptilodactilydae (6,8 %). En cuanto a la diversidad alfa, la quebrada Villa Paola registró una riqueza de 0,50. La dominancia fue alta (0,82), mientras la equidad de Pielou (0,31) y la diversidad de Shannon-Wiener fueron bajas (1,0 bits/ind.). En la quebrada Doña Juana se registró una riqueza de 0,6. En Villa Paola la dominancia fue alta (0,78), la equidad (0,33) y diversidad (0,87 bit/ind.) bajas. El índice de diversidad beta entre quebradas arrojo 0,40.
Fitoplancton. Esta comunidad estuvo representada por cinco divisiones, 19 familias y 15 géneros (tabla 3). En Doña Juana se encontraron 1,73 ind./ml en sequía y 25,88 ind./ml en lluvias, mientras que en la quebrada

Revista de Investigaciones No. 17 - Universidad del Quindío
63
Villa Paola se hallaron 97,20 ind./ml en sequía y 15,00 ind./ml en lluvias. En la quebrada Villa Paola las divisiones más abundantes fueron Cyanophycea (Sequía 74,7 %, Lluvia 54,2 %), Bacillarophycea (Sequía 5,3 %, Lluvia 29,1 %) y Chlorophyta (Sequía 20,1 %, Lluvia 0 %). A su vez, los grupos más representativos en las dos épocas fueron Chroococcaceae (37,0 %), Bacillariaceae (14,9 %), Characeae (9,5 %) y Rhodophyceae (8,3 %). Los géneros más abundantes en ambas épocas fueron Coelosphaerium (37,0 %) y Nitzschia (14,9 %). Chlorophyta fue la división más representativa en la quebrada Doña Juana con 82,6 % en sequía y 15,9 % en lluvia, seguida por Bacillarophycea con 11,3 % en sequía y 81,8 % en lluvias. A su vez las más representativas en ambas épocas fueron Characeae con 38,6 %, Mixophiceae con 17,0 % y Bacillariaceae con 12,9 %.
Zooplancton. Esta comunidad estuvo representada por seis clases, 11 familias y 13 géneros (tabla 4). En Doña Juana se encontraron 7,70 ind./ml en sequía y 6,15 ind./ml en lluvias, mientras que en la quebrada Villa Paola se encontraron 29,40 ind./ml en sequía y 0,63 ind./ml en lluvias. Para la quebrada Villa Paola el taxón más abundante fue Rhizopoda (Sequía 60,0 %, Lluvia 0 %), seguido por Amoebae (Sequía 22,0 %, Lluvia 0 %) y Granuloreticulocea (Sequía 12,0 %, Lluvia 0 %). De esta forma, los de mayor abundancia relativa en ambas épocas fueron Vorticellidae (13, 0%), Reticulocidae (8,0 %) y Biomixidae (6,0 %). Los géneros con mayor representatividad fueron Vorticella (13,0 %) y Biomyxa (6,0 %). En la quebrada Doña Juana el taxón más representativo fue Rhizopoda (Sequía 49,5, Lluvia 0 %), seguido por Amoebae (Sequía 37,8, Lluvia 0 %) y Ciliophora (Sequía 13,5, Lluvia 0 %). La más abundantes en ambas épocas fueron Amoebidae (25,0 %), Actinocomidae (18,9 %), Arcellidae (14,9 %) y Vorticellidae (5,4 %). A su vez los
géneros más representativos en ambas épocas fueron Ammoebas (25,0 %), Chlamydophris (25,0 %) y Acanthocystis (18,92 %). La diversidad alfa para fitoplancton y
zooplancton se consigna en la tabla 5. Con respecto al fitoplancton, en sequía la quebrada
Doña Juana registró alta dominancia (0,62) y baja equidad (0,37). En lluvias ocurrió lo contrario (D= 0,22, E= 0,85). En ambos épocas la diversidad fue baja (Sequía 0,87; Lluvia = 0,83) y la riqueza alrededor de 0,90. En la quebrada Villa Paola se registró igual tendencia de valores de diversidad alfa entre épocas (tabla 5). Con respecto al zooplancton en la quebrada Doña Juana, en promedio, la diversidad alfa se constituyo así: H’= 0,30 bit/ind., D= 0,45, E= 0,88 y M= 0,54. A su vez para Villa Paola la diversidad alfa arrojó que H’= 0,45 bit/ind., D= 0,57, E= 0,46 y M= 0,45. La prueba t-student para analizar la diversidad alfa de ambas comunidades de zooplancton arrojó diferencias significativas entre sequía y lluvia
en Villa Paola, en cuanto al índice de Simpson (t= 2,321). Sin embargo, no hay de diferencias significativas entre la diversidad alfa de ambas
quebradas (tabla 6). La diversidad beta para fitoplancton y zooplancton en ambas quebradas
arrojo un valor de 0,56.
Peces: Se registraron dos órdenes que incluyen cinco géneros y cinco especies, con un total de 150 individuos. La quebrada Villa Paola albergó una especie más que Doña Juana y un mayor número de ejemplares (82 en total) (tabla 7). Hemibrycon sp. fue la especie más abundante y común entre los afluentes; 86 individuos
en total. La diversidad y dominancia para los dos drenajes fueron bajos (H’= 1,09, D= 0,44 en Villa Paola y H’= 1,08, D= 0,33) en Doña Juana. Los demás componentes de la diversidad alfa arrojaron: E= 0,68 y M= 0,90 en Villa Paola y E= 0,79 y M= 0,71 en Doña Juana. No se observaron diferencias para la diversidad alfa

Revista de Investigaciones No. 17 - Universidad del Quindío
64
entre épocas y para los dos afluentes.
DISCUSIÓN
Las variables fisicoquímicas fueron óptimas para
caracterizar el ambiente acuático. El análisis de precipitación pluvial multianual mensual permitió inferir que el presente estudio fue realizado en sequía y lluvias. Según Blanco et al. (2003) los cambios en precipitación y temperatura del aire en las cuencas hidrográficas tienen efectos sobre
la vegetación, el suelo y el agua, que en éste caso producto de las perturbaciones climáticas pudo causar el desajuste en la abundancia y frecuencia de lluvias y temperaturas, con las épocas propuestas de lluvia y sequía. Las variables fisicoquímicas (excepto déficit de
oxígeno) de las quebradas Villa Paola y Doña Juana registraron bajos coeficientes de variación,
posiblemente debido a su posición geográfica y
localización topográfica en ambientes de alta
montaña neotropical, donde las variaciones son muy leves. Además, en el neotrópico algunas variables tienen un margen muy estrecho de fluctuaciones (Roldán, 1992). Se registró el
oxigeno disuelto de la botella clara en ambos drenajes fue un poco más alto que la oscura; esto podría tener explicación en el hecho de que las quebradas no presentaron una estructura boscosa ribereña lo suficientemente densa como
para regular la entrada de luz al cuerpo de agua, tal como lo demostraron Román-Valencia et al. (2005). En el caso de la quebrada Doña Juana se registró una disminución en el oxígeno disuelto de las Botellas DBO en lluvias, coincidente con la idea anterior, pues en esta época la radiación solar disminuye y por ende la temperatura que finalmente afecta la actividad fotosintética
(Roldán, 1992; Blanco et al. 2003). Se comprueba con la ANDEVA para las botellas DBO en la quebrada Villa Paola, influencia significativa de
las épocas sobre el oxígeno, los cuales pueden responder a un complejo juego de factores.
Durante el ciclo nictemeral se obtuvieron en las quebradas Doña Juana y Villa Paola tendencias a la disminución de las temperaturas del agua máxima, mínima y del aire con la llegada de la noche. También hubo tendencias a valores más altos en la temperatura superficial y a un
aumento de la humedad relativa en la noche. Según Roldán (1992), Urrego y Ramírez (2000) y Román-Valencia et al. (2005) el enfriamiento convencional nocturno es un fenómeno común, que se relaciona a su vez con el aumento de la humedad relativa. Además, la temperatura del agua tiende a fluctuar menos y ser más baja que
las otras temperaturas ya que posee un alto calor especifico, que junto con los procesos de mezcla
(causados por la velocidad de la corriente) y el sustrato crean una resistencia a los rayos solares (Urrego y Ramírez, 2000; Román-Valencia et al., 2005). Un comportamiento atípico se registró en el pH para ambas quebradas (Figs. 2 y 3), ya que a finales de la tarde (17: 00 hr.)
se registró disminución notable, posiblemente a causa de dos eventos: 1) lluvias durante el muestreo generaron procesos de escorrentía y lixiviado que llegaron a los drenajes elementos ácidos producto del tipo de vegetación ribereña (Pinus y Eucalyptus, los cuales acidifican los
suelos) y 2) las quebradas en ese momento tuvieron un déficit en el sistema buffer provisto
de bicarbonatos, carbonatos y CO2 (Roldán, 1992; Horne y Goldman, 1994). Inclusive es posible que la estructura boscosa ribereña evite que la carga de nutrientes y fertilizantes (origen agrícola), producto de la escorrentía y el lixiviado, fuera de mayor impacto sobre el comportamiento nictemeral (Chapin et al., 2002). La conductividad de las quebradas Doña Juana y Villa Paola, estuvieron dentro de los rangos reportados para ecosistemas de alta montaña neotropical (Roldán, 1992). El oxígeno disuelto y al porcentaje de saturación para ambas quebradas se hallaron dentro de los rangos óptimos para el desarrollo de las comunidades

Revista de Investigaciones No. 17 - Universidad del Quindío
65
acuáticas (Roldán, 1992). Además, su aumento en la noche se debe a que en esos momentos la temperatura del agua permitió la solubilidad de este gas (Roldan, 1992). Caso especifico de
la influencia de las épocas sobre las quebradas
fue el que ocurrió en Villa Paola, donde el ANDEVA mostró diferencias significativas,
lo cual indicó que en sequía y lluvia se alteran de forma distinta el oxígeno de los cuerpos de agua. La D. B. O. se registró en ambos afluentes
menor que la D. Q. O, debido a que el número de compuestos oxidados químicamente es mayor a los oxidados biológicamente, como lo explica Roldán (1992). Los sólidos y las durezas en época lluviosa de la quebrada Villa Paola fueron altos. Las aguas de éstas quebradas pueden clasificarse como moderadamente
duras, mientras que las de quebrada Doña Juana fueron blandas. En ambas quebradas la acidez fue menor a la alcalinidad, lo cual es producto del sistema buffer en el cual el pH, la alcalinidad y la acidez están regulados por la concentración de CO2. Con respecto al cambio de éstas variables entre épocas Blanco et al. (2003) plantearon que con la disminución de las lluvias y el incremento de la temperatura se reduce el crecimiento de la vegetación terrestre y aumenta la evapotranspiración y la mineralización de los suelos, lo que conlleva a una disminución del flujo de materia orgánica (caso contrario para
lluvias). El déficit de oxígeno registrado para
ambas quebradas mostró que en la quebrada Villa Paola se presentó un mayor consumo de oxígeno (Fig. 4) identificado en valores
negativos. Además, se registraron fluctuaciones
altas en el ciclo día-noche. Según Roldán (1992) el consumo de oxígeno, para valores negativos, es más pronunciado en un hábitat acuático durante la noche, cuando la luz está ausente y la actividad fotosintética es casi nula. Es decir, que la quebrada Villa Paola fue un sistema más productivo; sus altos déficit de oxígeno
reflejaron altas tasas de producción. Además, los
valores de oxígeno estuvieron cercanos al límite de calidad (5 mg/l). Los Sólidos y durezas altas y alto déficit de oxígeno indican un cuerpo de
agua en proceso de eutroficación. La quebrada
Doña Juana, por su parte, se conserva dentro de valores típicos de un sistema oligotrófico. En
general, las variables fisicoquímicas reportadas
en éste trabajo corresponden con lo reportado para quebradas del alto Cauca (Román-Valencia y Muñoz 2001; Román-Valencia et al., 2003; Román-Valencia y Ruiz, 2005; Román-Valencia et al., 2005).La comunidad de macroinvertebrados para ambas quebradas estuvo dentro de lo registrado para otros drenajes (Rincón, 2002; Londoño et al., 2005). Hemiptera fue el grupo más representativo; según Roldán (1992) y Rincón (2002) este grupo comparte el mismo hábitat de Odonata, e indica desde aguas limpias hasta medianamente contaminadas. Sin embargo, existió una diferencia notable entre los dos sistemas acuáticos, ya que en la quebrada Doña Juana fueron más abundantes los Trichoptera (Hydropsychidae), mientras que en Villa Paola predominaron los Diptera, en especial los Simuliidae y Tipulidae. Este fenómeno resalta el posible proceso de eutroficación
que sufre el ecosistema de Villa Paola. La diversidad de ambas quebradas con respecto a los macroinvertebrados fue baja comparado con otros estudios reportados para quebradas neotropicales (Roldán et al., 2001; Rincón, 2002). La diversidad beta arrojó poca diferencia entre las especies que se pierden y las que se ganan en cuanto a macroinvertebrados en ambos drenajes.La comunidad fitoplanctónica anotada aquí
coincide con otros estudios para el neotrópico (Ramírez y Alcaráz, 2002; Sierra et al., 2004 Hernández-Atilano et al., 2005). Sin embargo, para Chroococcaceae (Coelosphaerium) no se encontró ningún estudio especifico. Roldán
(1992) señala que los Chroococcaceae del

Revista de Investigaciones No. 17 - Universidad del Quindío
66
género Coelosphaerium prefiere ambientes
productivos, más comunes en lagos. La quebrada Villa Paola registró más abundancia de Cyanophyceae y Bacillarophyceae, mientras que en Doña Juana predominó Chlorophyta. Estas últimas son propias de ecosistemas de alta montaña donde tienen un desarrollo óptimas cuando hay disponibilidad de luz y sustrato (Roldán 1992). El género Nitzschia
(diatomea) fue abundante en Villa Paola, mientras que en Doña Juana fue Volxox. Según Roldán (1992) y Roldán et al. (2001) éstos dos géneros son típicos en Colombia. Las diatomeas (Bacillarophyceae) fueron representativas en ambas quebradas lo cual coincide con lo hallado por Roldán et al. (2001). Las diatomeas se han utilizado ampliamente como indicadoras de las condiciones del medio acuático. Su importancia radica en la capacidad que tienen para registrar rápidamente cambios con influencia de las
características físicas y químicas; por ejemplo el género Nitzschia es abundante cuando el pH es mayor a seis (Díaz-Quirós y Rivera-Rondón, 2004).El zooplancton presente (Rhizopoda principalmente en ambas quebradas) modifica
la estructura de la comunidad fitoplanctónica de
géneros como Nitzschia y Volvox, a través del consumo diferencial de algunas especies que le son más fáciles de ingerir por su tamaño y forma. La diversidad alfa de fitoplancton y zooplancton
fue baja, debido a la dominancia entre los ecosistemas (cercanos a uno). Según Townsend et al. (1997) y Blanco et al. (2003) se acepta que en ambientes de aguas con corrientes de baja velocidad la ausencia de agentes que causan remoción física de especies lleva al monopolio del espacio por parte de unas pocas. Sin embargo,
la riqueza del zooplancton fue apreciable para considerar una eficiente optimización de las
condiciones físicas y químicas del agua. La diversidad beta en ambas comunidades fue de 0,56, lo cual es bajo. Este fenómeno de alguna forma lo corroboró el resultado de la prueba t-student, la cual muestra que no se presentaron diferencias entre los promedios de la diversidad entre ambos drenajes. La comunidad de peces coincide con registros previos (Román-Valencia 1993; 1995). La abundancia relativa representada por las especies de carácidos encontradas hacen pensar que las quebradas no se encuentran fuertemente intervenidas por acción antrópica (distinto a lo arrojado por el análisis de las variables fisicoquímicas); ya que los caracidos
son los primeros en cambiar su estructura comunitaria al presentar algún cambio en las condiciones fisicoquímicas de los cuerpos de
agua (Lowe-McConnell, 1987). La abundancia de Hemibrycon sp. coincidió con lo reportado por Román-Valencia et al. (2005) para dos afluentes de la zona de estudio. El reporte de
Cetopsorhamdia boquillae en la quebrada Villa Paola es indicador biótico de la eutrofización, ya
que esta especie se caracteriza por presentarse en este tipo de ambientes, con abundante materia orgánica en descomposición e insectos (Román-Valencia y Giraldo, 2006). Por lo tanto, las quebradas Villa Paola y Doña Juana son cuerpos de agua oligotróficos, aunque la quebrada Villa
Paola tiende hacia la eutroficación. La diversidad
alfa de ambas quebradas fue baja mientras que la diversidad beta indicó que las dos quebradas son parecidas en cuanto a su composición de familias y géneros.

Revista de Investigaciones No. 17 - Universidad del Quindío
67
BIBLIOGRAFÍA
APHA, American Public Health Association, American Waterworks, Association (AWWA), 1. Water Pollution Control Federation (WPCF). 1998. Standard Methods for examination of Water and Sewage and Wastewater. 20a ed. Nueva York
Arboleda, J. y Ramirez J. (2002). Efecto de la presencia de macrófitas acuáticas en variación de 2. la biomasa, la producción primaria y la eficiencia fotosintética de la comunidad fitoplanctónica
de la laguna Francisco José de Caldas. Rev. Actual. Biol. 24 (76): 59-67.
Bicudo, E. y Bicudo R. 1970. Algas de águas continentais brasileiras. São Paulo: Fundação 3. brasileira para o desenvolvimento do ensino de Ciências. 157 p.
Blanco, J; Vásquez G.; Ramírez J.; y Navarrete A. 2003. Variación de algunos parámetros 4. fisicoquímicos en el río Pescador, Valle del Cauca, durante el ciclo El Niño 1997/1998-La Niña
1998/1999. Rev. Actual. Biol. 25(78): 59-69.
Botero, P.; García L.; Hernández C.; Sepúlveda L.; y Torres E. 2005. Evaluación de la actividad 5. tectónica reciente del frente montañoso occidental de la cordillera Central entre Calarcá y Pijao (Quindío-Colombia). Revista Geográfica Venezolana. Numero especial: 57-68.
Caicedo, O. y Palacio J. 1998. Los macroinvertebrados bénticos y la contaminación orgánica en 6. la quebrada La mosca (Guarne, Antioquia, Colombia). Rev. Actual. Biol. 20 (69):61-73.
Cardona, M.; Román-Valencia C.; Jiménez J. y Hurtado H. 1998. Composición y diversidad 7. de los peces de la quebrada San Pablo en alto Cauca, Colombia. Bol. Ecotrópica: Ecosistema Tropicales 32:11-24.
Chapin, I.; Matson P. y Mooney H. 2002. Principles of Terrestrial Ecosystem Ecology. Nueva 8. York: Springer Science+Business Media, Inc. 436p.
Correa, M.; Machado T. y Roldan G. 1981. Taxonomía y ecología del orden Trichoptera en el 9. departamento de Antioquia en diferentes pisos altitudinales. Rev. Actual. Biol. 10(36): 35-48.
Díaz-Quiros, C. y Rivera-Rondon C. 2004. Diatomeas de pequeños ríos andinos y su utilización 10. como indicadores de condiciones ambientales. Caldasia. 26(2): 381-394.
Gallo-Sanchez, L.; Gaviria-Melo S. y Ramirez-Restrepo J. 2004. Dinámica de la comunidad 11. zooplanctonica (excepto protozoa) en la laguna del Parque Norte, Medellín (Antioquia, Colombia). Rev. Actual. Biol. 26(81): 231-241.
Gonzalez, H. y Nuñez A. 1991. Mapa geológico generalizado del departamento del Quindío 12. escala 1:100.000. Memoria explicativa. Ingeominas, Bogotà. 1-47 p.

Revista de Investigaciones No. 17 - Universidad del Quindío
68
Hernández-Atilano, E.; Aguirre N. y Palacio J. 2005. Variación espacio-temporal de la estructura 13. de la comunidad de algas perifíticas en la microcuenca de la quebrada La Vega, municipio de San Roque (Antioquia), Colombia. Rev. Actual. Biol. 27(82): 67-77.
Horne, A. y Goldman C. 1994. Limnology. 2 ed. Nueva York: McGraw-Hill, Inc. 215 p.14.
Humphries, C.; Williams P. y Vane-Wright R. 1995. Measuring biodiversity value form 15. conservation. Ann. Rev. Ecol. Syst. 26: 93-111.
Kudo, R. 1966. Protozologia. México, D. F. 345 p.16.
Lackey, J. 1956. Zooflagelados. En: Ward y Whipple, editors. Freshwater biology. Nueva York: 17. Edimmsem. p.190-231.
Lampert, W. y Sommer U. 1997. Limnoecology: The ecology of Lakes and Streams. Nueva 18. York: Oxford University Press; 382 p.
Londoño, A.; Arrubla J.; Toro J.; Torres D.; Zarate M. y Beltrán M. 2005. Determinación de la 19. calidad ambiental en la quebrada Agua Bonita, corregimiento de Barcelona, departamento del Quindío. Revista de Investigaciones Universidad del Quindío 15: 55-64.
Lowe-McConnell, R. H. 1987. Ecological studies in tropical fish communities. New York: 20. Cambridge, University Press. 382 p.
Machado, T. y Roldán G. 1981. Estudio de las características fisicoquímicas y biológicas del río 21. Anori y sus principales afluentes. Rev. Actual. Biol (35): 10-19.
Matthias, U. y Moreno H. 1983. Estudio de algunos parámetros fisicoquímicos y biológicos en 22. el río Medellín y sus principales afluentes. Rev. Actual. Biol 12(46): 106-117.
Medellín, F.; Ramírez M. y Rincón M. 2004. Trichoptera del Santuario de Iguaque (Boyacá, 23. Colombia) y su relación con la calidad del agua. Revista Colombiana de Entomología 30(2): 197-203.
Needham, J. y Needham P. 1978. Guía para el estudio de los seres vivos de las aguas dulces. 24. Barcelona: Reverte, S. A. 153 p
Palacio–Núñez, J., Verdú, J. R., Galante, E., Jiménez–García, D. & Olmos–Oropeza, G. 2007. 25. Birds and fish as bioindicators of tourist disturbance in springs in semi–arid regions in Mexico:
a basis for management. Animal Biodiversity and Conservation, 30.1: 29–41.
Posada-García, J. y Roldan-Pérez G. 2003. Clave ilustrada de las larvas de Trichoptera en el 26. nor-occidente de Colombia. Caldasia 25(1): 169-192.

Revista de Investigaciones No. 17 - Universidad del Quindío
69
Ramírez, J. y Alcaraz H. 2002. Dinámica de la producción primaria fitoplanctónica en un 27. sistema eutrófico tropical: laguna del Parque Norte, Medellín, Colombia. Caldasia 24 (2): 411-
423.
Rincón, M. 1996. Aspectos bioecológicos de los Tricópteros de la quebrada Carrizal (Boyacá, 28. Colombia). Revista Colombiana de Entomología 22 (1): 53-60.
Rincón M. 2002. Comunidad de insectos acuáticos de la quebrada Mamarramos (Boyacá, 29. Colombia). Revista Colombiana de Entomología 28 (1): 101-108.
Roldán, G. 1992. Fundamentos de Limnología Neotropical. Fen-Universidad de Antioquia, 30. Medellín, 529 p.
Roldán, G. 1996. Guía para el estudio de los macroinvertebrados acuáticos del departamento 31. de Antioquia. FEN-Colciencias, Bogotá, 217 p.
Roldan, G.; Posada J.; y Gutiérrez J. 2001. Estudio limnológico del recurso hídrico del parque 32. Piedras Blancas. Academia Colombia de Ciencias Exactas, Físicas y Naturales. Colección “Jorge Alvares Lleras”, Bogotá, Colombia. 152 p.
Román-Valencia, C. 1993. Composición y estructura de las comunidades de peces de la cuenca 33. del río La Vieja, Alto Cauca. Biol. y Educ. (5): 8-19.
Román-Valencia, C. 1995. Lista anotada de los peces en la cuenca del río La Vieja, Alto Cauca, 34. Colombia. Bol. Ecotrópica: Ecosistema Tropicales (29): 11-22.
Román-Valencia, C. y Muñoz A. 2001. Ecología trófica y reproductiva de Bryconamericus 35. caucanus (Pisces: Characidae). Boll. Mus. Reg. Sci. Nat. Torino. 18(2): 459-467.
Román-Valencia, C. 2003. Sistemática de las especies colombianas de Bryconamericus 36. (Characiformes: Characidae). Dahlia, Rev. Asoc. Colomb. Ictiol. 6: 17-58.
Román-Valencia, C.; Botero A. & Ruiz-C. R. 2003. Trophic and reproductive ecology of 37. Roeboides dayi (Teleostei: Characidae) from upper Rio Cauca, Colombia. Boll. Mus. Reg. Sci. nat. Torino. 20(2): 487-496.
Román-Valencia, C. y Perdomo A. 2004. Ecología trófica y reproductiva de Argopleura 38. magdalenensis (Pisces: Characidae) en la cuenca alta de los ríos Cauca y Magdalena, Colombia. Rev. Mus. Argentino Cienc. Nat., n. s. 6 (1): 175-182.
Román-Valencia, C. y Ruiz R. I. 2005. Diet and reproduction aspects of Astyanax aurocaudatus 39. (Teleostei: Characidae) from the upper part of the Rio Cauca, Colombia. Dahlia (Rev. Asoc. Colomb. Ictiol.) 8: 9-17.

Revista de Investigaciones No. 17 - Universidad del Quindío
70
Román-Valencia, C.; Cadavid J.; Vanegas J. y Arcila D. 2005. Análisis de algunas variables 40. físicas, químicas y biológicas en tres quebradas de la Cuenca Alta del Río Cauca, Colombia. Revista de Investigaciones Universidad del Quindío 15: 83-96.
Román-Valencia, C. y Giraldo A. 2006. Tròfica y reproducción de Cetopsorhamdia boquillae 41. (Pisces: Siluriformes) en Río La Vieja, Alto Cauca, Colombia. Revista de Investigaciones Universidad del Quindío 16: 49-61.
Román-Valencia, C., Ruiz-C, R. 2007. Una nueva especie de pez del género Hemibrycon 42. (Characiformes: Characidae) del Alto Río Atrato, noroccidente de Colombia. Caldasia, 29(1):75-85.
Ruiz-C, R. I. y Román-Valencia, C. 2006a. Osteología de Astyanax aurocaudatus, Eigennman, 43. 1913 (Pisces: Characidae), con notas sobre la validez de Carlastyanax, Gèry, 1972. Animal Biodiversity and Conservation. 29 (1): 49-51
Ruiz-C, R. I. y Román-Valencia, C. 2006b. Aspectos taxonómicos de Cetopshorhamdia 44. boquillae y C. nasus (Pisces: Heptapteridae), con anotaciones sobre su ecología en al cuenca alta de los ríos Magdalena y Cauca, Colombia. Animal biodiversity and Conservation, 29 (2): 123-131.
Sierra, O.; Ramírez J. y Marín O. 2004. Dinámica temporal de la comunidad fitoplanctónica 45. y variables físicas y químicas asociadas al lago Santander (Rió Negro, Antioquia, Colombia). Rev. Actual. Biol. 26 (80): 30-38.
Townsend, C.; Scarsbrook M. y Doledec S. 1997. The intermediate disturbance hypothesis, 46. refugia and biodiversity in streams. Limnol. Oceanogr. 42(5): 938-949.
Uhlerkovich, A. y Schmidt G. 1974. Phyplanktantaxo in dem zentralamazonishen 47. schwemmlandsee lago do Castanho. Amazoniana 2: 243-283.
Urrego, A. y Ramírez J. 2000. Cambios diurnos de variables físicas y químicas en la zona de 48. ritral del río Medellín. Caldasia 22(1): 127-141.
Wetzel, R. y Likens G. 2000. Limnological analyses, 3 ed. Nueva York: Springer-Verlag. 429 49. p
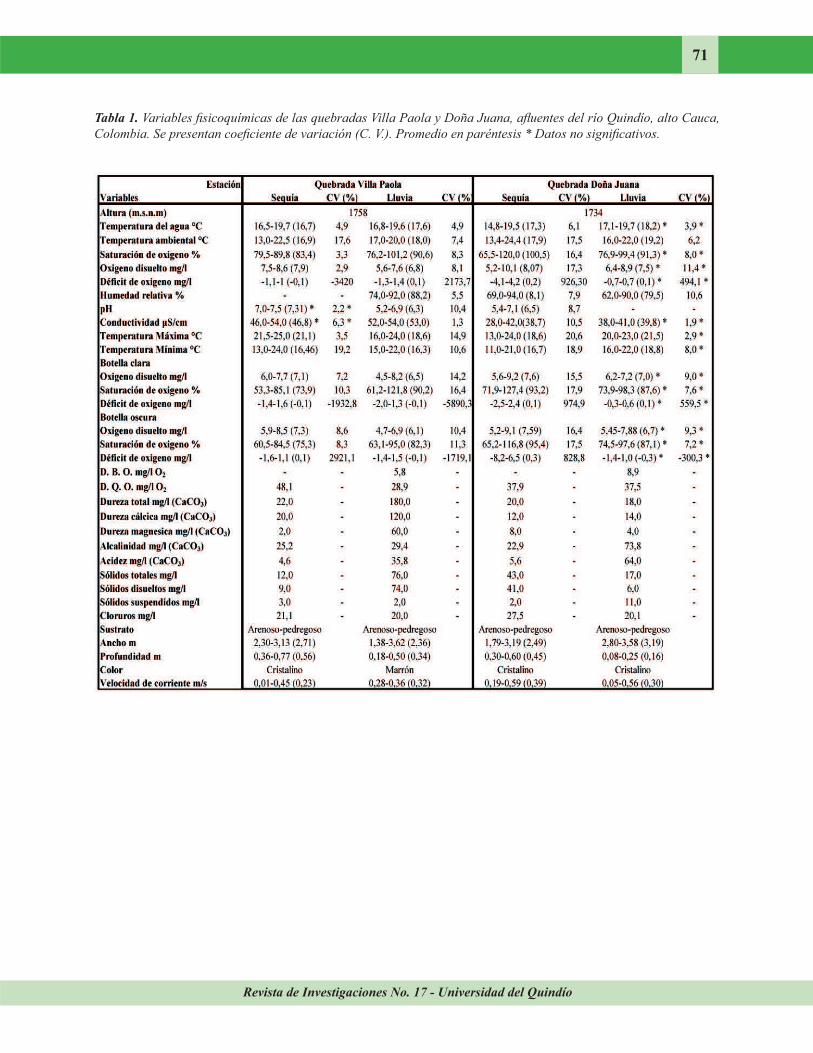
Revista de Investigaciones No. 17 - Universidad del Quindío
71
Tabla 1. Variables fisicoquímicas de las quebradas Villa Paola y Doña Juana, afluentes del río Quindío, alto Cauca,
Colombia. Se presentan coeficiente de variación (C. V.). Promedio en paréntesis * Datos no significativos.
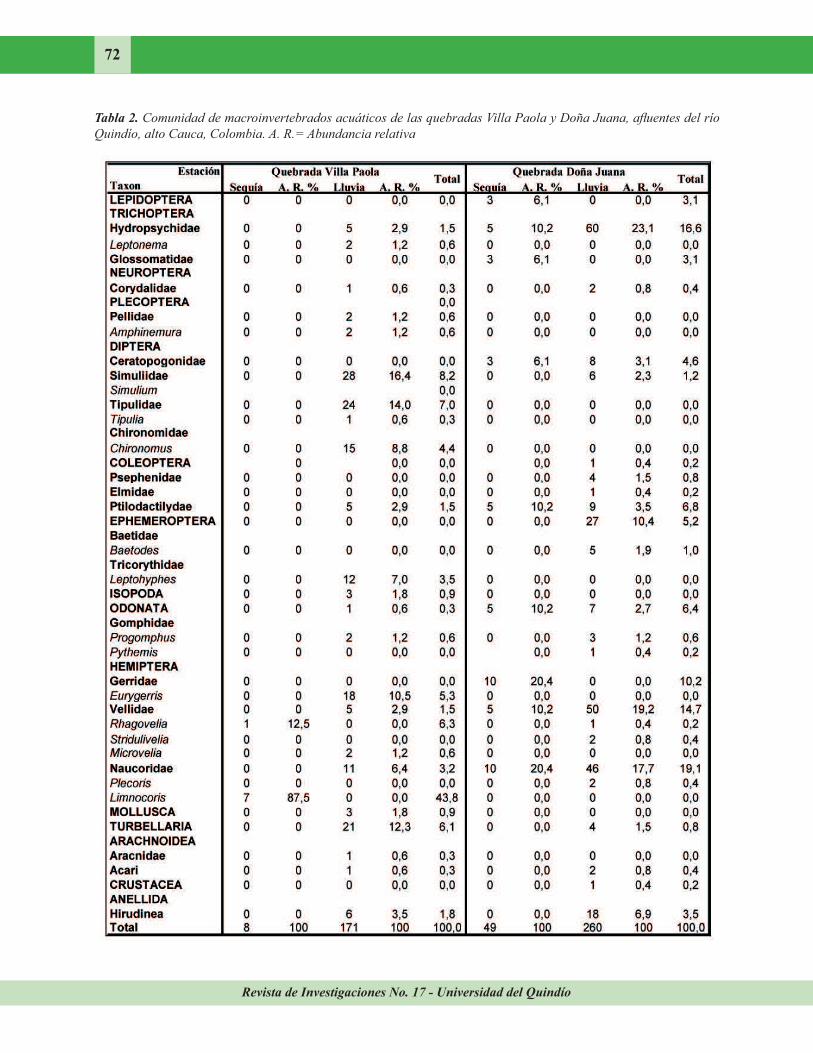
Revista de Investigaciones No. 17 - Universidad del Quindío
72
Tabla 2. Comunidad de macroinvertebrados acuáticos de las quebradas Villa Paola y Doña Juana, afluentes del río
Quindío, alto Cauca, Colombia. A. R.= Abundancia relativa

Revista de Investigaciones No. 17 - Universidad del Quindío
73
Tabla 3. Comunidad fitoplanctónica de las quebradas Villa Paola y Doña Juana, afluentes del río Quindío, alto Cauca,
Colombia. A. R.= Abundancia relativa
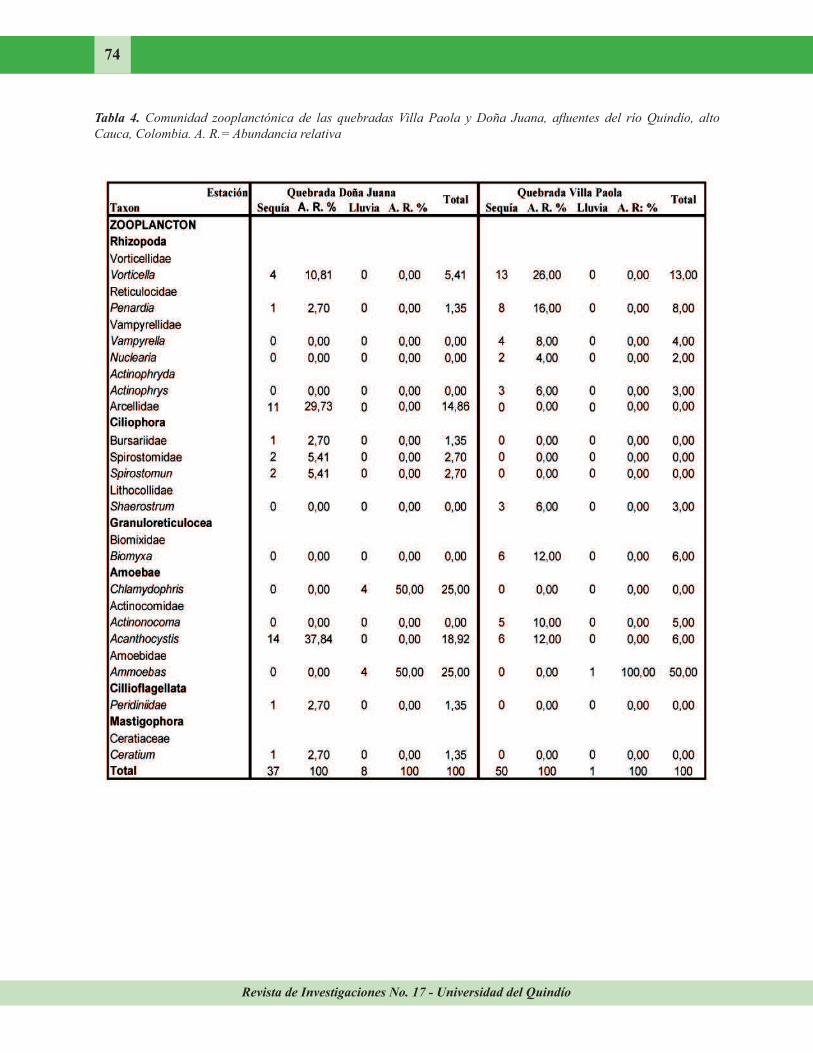
Revista de Investigaciones No. 17 - Universidad del Quindío
74
Tabla 4. Comunidad zooplanctónica de las quebradas Villa Paola y Doña Juana, afluentes del río Quindío, alto
Cauca, Colombia. A. R.= Abundancia relativa
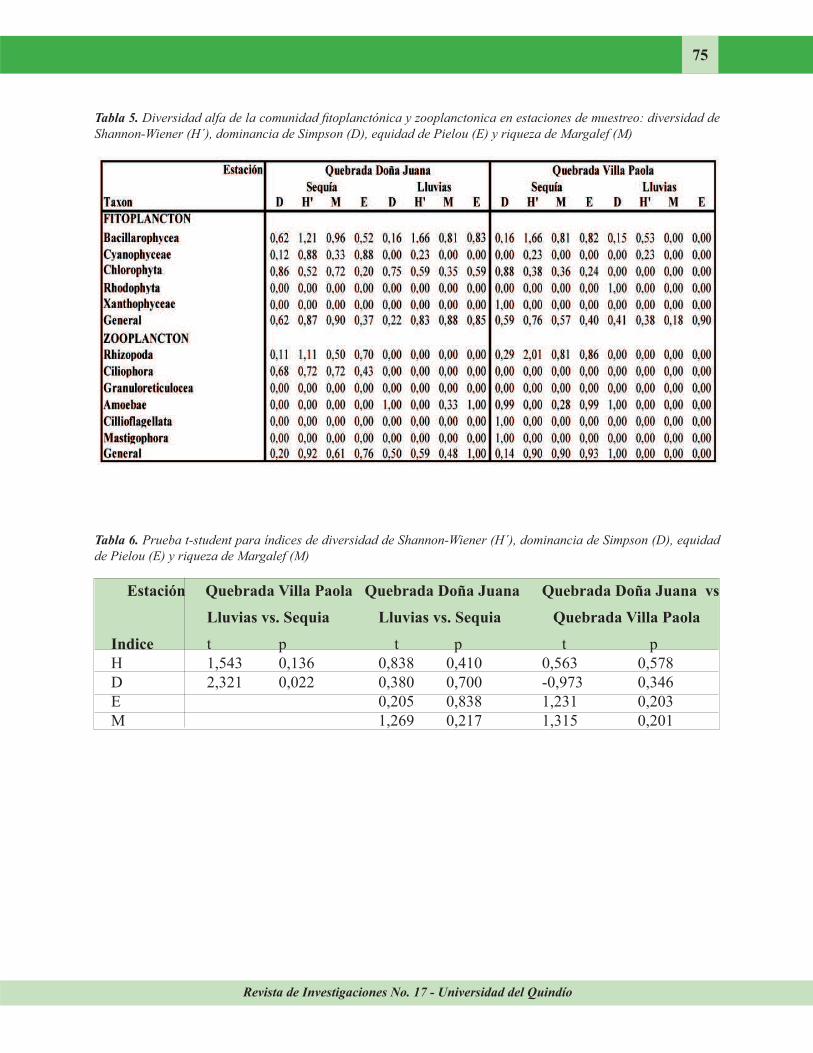
Revista de Investigaciones No. 17 - Universidad del Quindío
75
Tabla 5. Diversidad alfa de la comunidad fitoplanctónica y zooplanctonica en estaciones de muestreo: diversidad de
Shannon-Wiener (H´), dominancia de Simpson (D), equidad de Pielou (E) y riqueza de Margalef (M)
Tabla 6. Prueba t-student para índices de diversidad de Shannon-Wiener (H´), dominancia de Simpson (D), equidad
de Pielou (E) y riqueza de Margalef (M)
Estación Quebrada Villa Paola Quebrada Doña Juana Quebrada Doña Juana vs
Lluvias vs. Sequia Lluvias vs. Sequia Quebrada Villa Paola
Indice t p t p t p H 1,543 0,136 0,838 0,410 0,563 0,578 D 2,321 0,022 0,380 0,700 -0,973 0,346 E 0,205 0,838 1,231 0,203 M 1,269 0,217 1,315 0,201
Revista de Investigaciones No. 17 - Universidad del Quindío
76
Tabla 7. Abundancia absoluta (AB) y relativa (AR) de peces de las quebradas Villa Paola y Doña
Juana, Río Quindío, Alto Cauca.
Leyenda de figuras:
Figura 1. Datos pluviométricos de la estación La Vieja, Salento, Colombia. Año 2006. Datos de IDEAM.
Figura 2. Comportamiento nictemeral de variables fisicoquímicas de la quebrada Doña Juana,
afluente del río Quindío, alto Cauca, Colombia. a. Botellas DBO oscura y clara en época seca, b.
Temperatura máxima y mínima en época de sequía y lluvia, c. pH en época seca y conductividad en sequía y Lluvia d. Temperatura del agua en sequía y temperatura ambiental en sequía y lluvia, e. Humedad relativa en época seca y lluviosa, f. Oxígeno disuelto y el porcentaje de saturación de oxígeno en época seca.
Figura 3. Comportamiento nictemeral de variables fisicoquímicas de la quebrada Villa Paola,
afluente del río Quindío, alto Cauca, Colombia. a. Botellas DBO oscura y clara en época seca,
b. Botellas DBO oscura y clara en época lluviosa, c. Oxígeno disuelto y porcentaje de saturación de oxígeno en sequía y lluvias, incluye la humedad relativa en lluvias, d. pH y conductividad en lluvias, e. Temperatura máxima y mínima en lluvias, f. Temperatura superficial y del aire en época
seca y lluviosa, en esta el eje secundario de Y (parte derecha) representa los datos correspondientes a temperatura del aire en ambas épocas.
Figura 4. Comportamiento nictemeral del déficit de oxígeno en las quebradas Villa Paola y Doña
Juana, afluentes del río Quindío, alto Cauca, Colombia. a. Botellas DBO clara y oscura en la
quebrada Doña Juana durante sequía, b. Oxígeno disuelto medido en las quebradas Doña Juana (sequía) y Villa Paola (sequía y Lluvias), c. Botellas DBO clara y oscura en la quebrada Villa Paola durante lluvias, d. Botellas DBO clara y oscura en la quebrada Villa Paola durante sequía.
Revista de Investigaciones No. 17 - Universidad del Quindío
77
Figura 1.
Revista de Investigaciones No. 17 - Universidad del Quindío
78
f
Figura 2.
a b
dc
e

Revista de Investigaciones No. 17 - Universidad del Quindío
79
Figura 3.
f
a b
dc
e
Revista de Investigaciones No. 17 - Universidad del Quindío
80
Figura 4.
a b
dc

Revista de Investigaciones No. 17 - Universidad del Quindío
81
Revista de Investigaciones No. 17 - Universidad del Quindío p p 81- 102 Armenia, Año 2007 ISSN 1794-631 X
Germinación, Nodulación y Crecimiento
de Elaeagnus Angustifolia L.Mercedes Girón Vanderhuck1, Francisco Bermúdez de Castro y Navia2, Esther Pérez Corona2
RESUMEN
En este estudio se determinó la viabilidad y germinación de las semillas de E. angustifolia, se evaluó la capacidad de las plántulas para producir nódulos fijadores de nitrógeno y se comparó el crecimiento de las plantas noduladas y no noduladas con diversos parámetros de uso habitual en este tipo de estudios. La disponibilidad lumínica, el lavado y la escarificación de las semillas de E. angustifolia son efectivos para inducir la germinación. Las plantas inoculadas con Frankia que crecieron en vermiculita nodularon, mientras que las plantas inoculadas y control, que se mantuvieron en macetas con suelo, no nodularon. Las raíces de las plantas noduladas desarrollaron pocos nódulos, una media de 2.5 racimos nodulares por planta. Cada racimo nodular estaba formado por 7.2 lóbulos, teniendo cada lóbulo una longitud aproximada de 1 mm. Las plantas inoculadas con Frankia y los individuos control, que crecieron en suelo, no se diferenciaron significativamente con respecto a los parámetros de crecimiento. Por el contrario, las plantas noduladas en vermiculita presentaron valores de tasa de crecimiento relativa, tasa de asimilación neta, área foliar específica y proporción del peso del tallo superiores a los de los individuos control que crecieron en vermiculita. La proporción del peso de la raíz fue mayor en los ejemplares plantados en vermiculita que en los que crecieron en suelo. Las plantas que crecieron en suelo y las noduladas en vermiculita presentaron concentraciones de nitrógeno y carbono mayores que los controles plantados en vermiculita.
Palabras clave: Elaeagnus angustifolia, germinación, nódulo, crecimiento, fijadora de nitrógeno.
ABSTRACT
In this study it was determined the viability and germination of the seeds of E. angustifolia, the capacity of the seedling was evaluated to produce nitrogen fixers nodules and it was compared the growth of the nodulated and non nodulated plants with diverse parameters of habitual use in this type of studies. The light readiness, the wash and the scarification of the seeds of E. angustifolia is effective to induce germination. The plants inoculated with Frankia that grew in vermiculite become nodulated, while inoculated and control plants that stayed in pots with soil, didn´t become nodulated. The roots of the plants developed few nodules, a stocking of 2.5 nodular clusters per plant. Each nodular cluster was formed by 7.2 lobes, having each lobe an approximate longitude of 1 mm. The plants inoculated with Frankia and the control individuals that grew in soil, didn’t have significant differences when compared to the parameters of growth. On the contrary, the nodulated plants in vermiculite, presented values of relative growth rate, net assimilation rate, specific leaf area and stem weight rate superiors to those of the control individuals that grew in vermiculite. The root weight rate was bigger in the seedlings planted in vermiculite than in those that grew in soil. The seedlings that grew in soil and nodulated plants in vermiculite, presented nitrogen and carbon concentrations higher than the controls planted in vermiculite.
Key words: Elaeagnus angustifolia, germination, nodule, growth, nitrogen fixer.
1 [email protected] Bióloga Ph.D. Docente Programa de Licenciatura en Biología y Educación Ambiental,
Universidad del Quindío.2 Ph. D. Docente en el Departamento de Ecología, Facultad de Ciencias Biológicas, Universidad Complutense de Madrid.

Revista de Investigaciones No. 17 - Universidad del Quindío
82
INTRODUCCIÓN
laeagnus angustifolia L., conocida como árbol del paraíso, es una planta actinorrícica que presenta simbiosis con un actinomiceto tipificado por
Becking (1970) como Francia (Miller y Baker, 1985; Zitzer y Dawson, 1989; Moretti, 1993). Esta especie, debido a su capacidad de fijar nitrógeno atmosférico, puede colonizar
terrenos muy pobres en nitrógeno y prepararlos para que se instalen otras especies vegetales (Chapin et al., 1994; Huss-Danell, 1997). Por consiguiente, desempeña una función muy importante en etapas serales de baja madurez, tanto en sucesiones primarias como secundarias (Bermúdez de Castro, 1988; Hibbs y Cromack, 1990). Al beneficio anterior se debe
añadir la regulación eficiente que ejerce sobre
la comunidad microbiana edáfica (Tarrant y
Trappe, 1971; Gutiérrez Mañero y Bermúdez de Castro, 1983; Gutiérrez Mañero, 1987; Llinares, 1990; Llinares, 1993; Pozuelo, 1991; Pozuelo et al., 1992; Llinares et al., 1993; Pérez Hernández, 1996; Mateus, 1997).
E. angustifolia tiene su origen biogeográfico en
Asia occidental donde forma parte de “schibljak” (Ron, 1971), comunidad submediterránea de arbustos caducifolios termófilos, que resisten
veranos muy calurosos y secos, e inviernos muy fríos y no excesivamente lluviosos (Rikli, 1943). En España, desde hace ya varios años se emplea esta especie como ornamental y actualmente se encuentra en parques y jardines de diferentes ciudades y pueblos (Zamorano, 1988). No se sabe exactamente cuando fue introducida, pero Linneus (1753) ya la reporta en el siglo XVIII. Según Ron (1971), cuando E.
angustifolia encuentra hábitats similares a los de su zona de origen, se naturaliza superando con éxito la etapa de ecesis. Éxito que se explica por su capacidad para crecer en un amplio
Erango de condiciones ambientales (Shafroth et
al., 1995; Katz y Shafroth, 2003).
Debido a su presencia en un sitio lejos de su lugar de origen y a su establecimiento en zonas donde las condiciones climatológicas y edáficas
son adversas para la mayoría de las plantas leñosas, surgió el interés de estudiar la viabilidad y germinación de las semillas de E. angustifolia. Se ha considerado que, entre los factores que pueden afectar a la germinación, estaría la disponibilidad lumínica, que se ha encontrado relevante para otras especies y que, en este caso, podría deberse, en condiciones naturales, a la existencia de doseles herbáceos o arbóreos que reducirían de forma significativa la luz que
llegaría a la semilla. Los resultados podrían ayudar a evaluar la capacidad de regeneración de esta especie y su posible evolución, a través del éxito en la reproducción sexual y la viabilidad de las semillas producidas.
Otro aspecto muy importante en el estudio de E. angustifolia es la capacidad que tiene esta especie para formar nódulos fijadores de
nitrógeno. Esta capacidad puede influir de
forma determinante en el éxito del desarrollo de las plántulas germinadas. Por esto, también se consideró el estudio de la producción de nódulos fijadores de nitrógeno en las plántulas y se
evaluó el crecimiento de las plantas noduladas y no noduladas con diversos parámetros de uso habitual en este tipo de estudios y que dan información sobre las distintas estrategias de desarrollo en diferentes condiciones. Los parámetros considerados son: RGR (tasa de crecimiento relativa), NAR (tasa de asimilación neta), LAR (proporción de área foliar), SLA (área foliar específica), LWR (proporción del
peso de las hojas), SWR (proporción del peso del tallo), RWR (proporción del peso de la raíz) y NUE (eficiencia del uso del nitrógeno).

Revista de Investigaciones No. 17 - Universidad del Quindío
83
MATERIALES Y MÉTODOS
Germinación de las semillas. Para el experimento de germinación se recolectaron al azar frutos maduros de árboles jóvenes de E.
angustifolia (Figura 1) en Valdemoro (Madrid) en octubre de 2000 y 2001. Los frutos cosechados se introdujeron en frascos de cristal tapados con algodón y se almacenaron en el frigorífico a una
temperatura de 3-4ºC, manteniéndolos hasta el inicio del experimento en estas condiciones. Un par de días antes de comenzar las pruebas de germinación se separaron las semillas del fruto eliminando la parte carnosa exterior. Las semillas se volvieron a guardar en el frigorífico
hasta su uso. El diseño experimental realizado considera el efecto de tres factores en la germinación de las semillas: 1) Viabilidad de las semillas procedentes de dos años de recogida consecutivos, 2) efecto del tratamiento previo o no del endocarpo y 3) efecto de la disponibilidad lumínica sobre la germinación.
En el experimento se utilizaron 800 semillas: 400 cosechadas en octubre de 2000 y otras 400 en 2001. Para cada año se dejó la mitad de las semillas sin tratamiento previo del endocarpo y la otra mitad se trataron. El tratamiento del endocarpo consistió en sumergirlas en agua destilada a 70ºC con agitación constante durante 24 horas. Después, en el extremo cercano al embrión se practicó una incisión en cruz, teniendo cuidado de no dañarlo. Las 400 semillas de cada cosecha, sometidas a este tratamiento del endocarpo, se distribuyeron en 16 bandejas con vermiculita exfoliada ASFALTEX, humedecida previamente con agua destilada. Se colocaron 25 semillas en cada bandeja cubriéndolas con una capa delgada de vermiculita. La misma distribución se realizó con las semillas no tratadas. Las 32 bandejas se colocaron en una cámara de germinación,
en condiciones de temperatura y humedad controladas (26ºC y 65% respectivamente). La mitad de las bandejas (16) se sometieron a un fotoperíodo de 12 horas de luz. Para ello se utilizaron 3 tubos fluorescentes de 18 watios cada
uno y 2 bombillas incandescentes de 100 watios que proporcionaban una intensidad lumínica a las bandejas de 2 500 lux y 40 mmol m-2 s-1 de PAR (radiación fotosintéticamente activa). La otra mitad de las bandejas (16) se mantuvo en oscuridad. Por lo tanto para cada combinación de tratamientos se tienen 8 réplicas. Los tratamientos son: a) Semillas con tratamiento del endocarpo y luz, b) semillas sin tratamiento del endocarpo y luz, c) semillas con tratamiento del endocarpo y oscuridad y d) semillas sin tratamiento del endocarpo y oscuridad. Estos tratamientos se repitieron para las semillas del año 2000 y del año 2001. Para cada año y tratamiento se consideraron entonces 8 réplicas. La distribución de las bandejas en cada caso se hizo al azar. Se realizó un seguimiento diario, durante dos meses, para registrar el número de semillas germinadas.
Trasplante, inoculación y crecimiento de las plantas. Con el fin de obtener el inóculo,
se cosecharon racimos nodulares de plantas jóvenes de E. angustifolia en Valdemoro (Madrid) en junio de 2002 y se transportaron en una nevera portátil, a una temperatura de 4ºC, al laboratorio.
Los racimos nodulares se limpiaron quitando las partículas de suelo, después se lavaron con agua destilada y se secaron con papel de filtro. Una
vez secos, se pesaron 2 gramos de nódulos y se trituraron en un mortero que contenía arena de mar lavada y 20 ml de agua destilada, obteniendo así un inóculo al 20%. Todo el procedimiento se realizó en condiciones estériles.
El experimento se inició el 10 de junio de 2002.

Revista de Investigaciones No. 17 - Universidad del Quindío
84
Antes de proceder a la inoculación, se eligieron 60 plántulas de E. angustifolia sin nódulos y se distribuyeron al azar en dos grupos de 30. Un grupo se consideró como control (sin inóculo) frente a la nodulación y se dividió en dos subgrupos: 20 plántulas que se trasplantaron a macetas con suelo y 10 a macetas con vermiculita. El control de suelo se utilizó para comprobar si el suelo recogido cerca del tronco de los árboles inducía nodulación. El otro grupo se destinó a la inoculación con el simbionte y su distribución en los macetas se hizo de igual forma que el grupo control. Se utilizaron macetas de plástico, cilíndricas, con un diámetro de 12 cm y una profundidad de igual dimensión. Cada maceta se lavó previamente con detergente libre de fosfato y abundante agua corriente y se finalizó el lavado
con agua destilada. Algunas características del suelo utilizado en el experimento aparecen en la Tabla 1. El suelo se recogió debajo del dosel de 5 árboles de E. angustifolia, a una distancia de 1 m del tronco del árbol y una profundidad de 20 cm. Cada plántula se inoculó sumergiendo su sistema radical en el macerado de nódulos durante 30 segundos. En todos los tratamientos se colocaron 2 plántulas por maceta.
Las plantas se regaron cada dos días con solución nutritiva de Crone, sin nitrato, a ¼ de su concentración y 1 ml l-1 de la solución de oligoelementos A-Z de Hoagland y se dejaron en el laboratorio en condiciones de fotoperíodo natural durante 27 semanas, registrando diariamente las temperaturas máximas y mínimas.
Cosecha y estimación del crecimiento de las plantas. Las plantas se cosecharon en diciembre de 2002 cuando tenían 6 meses de edad. Se separó cuidadosamente el suelo o la vermiculita de las raíces y se comprobó la existencia o no de nódulos en cada caso.
En las plantas que nodularon se estudió la ubicación de los nódulos midiendo la distancia desde la base de la raíz hasta el punto donde sobresalía el racimo nodular. Cuando los racimos nodulares aparecían en las raíces secundarias se tuvo en cuenta la distancia desde la base de la raíz hasta el punto de ramificación y de éste
hasta donde se encontraba el racimo nodular. Una vez que las plantas fueron cosechadas, se separaron en raíz, tallo y hojas. Se escanearon las hojas y se midió su área con el programa SCION IMAGE (Release beta 4.0.2 para Windows). Cada una de las partes de la planta se secó en una estufa de aire forzado MEMMERT 800 a 75-78ºC hasta peso constante (aproximadamente 48 horas). Entonces se pesaron en una balanza METTLER TOLEDO modelo COLLEGE B-303 con una precisión hasta de 3 decimales y se obtuvo el peso seco de raíz, tallo y hojas.
Para realizar el análisis de crecimiento se tuvo en cuenta la biomasa seca inicial y final de
las plántulas utilizadas en el experimento de inoculación. La biomasa seca inicial se infirió
a partir de 10 plántulas que fueron pesadas en seco y en fresco al principio del experimento. Con los datos del porcentaje peso fresco/peso seco de estas plantas y del peso fresco inicial de todas las plántulas utilizadas en el tratamiento, se calculó el peso seco inicial de las mismas.
También se determinó el carbono orgánico de las diferentes partes de la plántula utilizando el método de Walkley y Black (1934), optimizado posteriormente por Walkley (1935, 1947) y el nitrógeno total siguiendo el método de Kjeldahl (1883) modificado por Bremner y Tabatabai
(1972).
Con los pesos, áreas y contenido de nutrientes de las plántulas se obtuvieron los parámetros de crecimiento sugeridos por Welschen y Bergkotte (1994).

Revista de Investigaciones No. 17 - Universidad del Quindío
85
Análisis estadístico. Los resultados obtenidos en la germinación de las semillas de época diferente, tratadas y no tratadas y sometidas al efecto de luz y oscuridad, se compararon por medio de un ANOVA de tres factores. Antes de proceder al análisis estadístico se comprobó la normalidad y homogeneidad de las variables. Cuando no se encontró, los datos se transformaron logarítmicamente. En el caso de que se obtuvieran diferencias significativas se
realizaba la prueba LSD para conocer entre qué tratamientos se encontraban las diferencias.
Los resultados obtenidos en el análisis de crecimiento de las plantas inoculadas y sin inocular sembradas en suelo y vermiculita se compararon también por medio de un ANOVA de dos factores. Los análisis estadísticos se realizaron con el programa STATISTICA 5.1 (1997).
RESULTADOS Germinación de las semillas. La germinación de las semillas de E. angustifolia se detectó por la emergencia de la radícula y los cotiledones. La germinación de las semillas expuestas a la luz, en todos los tratamientos, fue a los 12 días de instalado el ensayo. En las semillas de 2000 sometidas a tratamiento del endocarpo se observó una germinación máxima de 19% a los 36 días, mientras que las no tratadas presentaron un máximo de 17% a los 30 días. En las semillas de 2001 tratadas, el valor más alto de germinación (20%) se observó a los 30 días y, en las no tratadas, con un valor del 11%, a los 36 días. Las últimas plántulas germinadas aparecieron a los 48 días de haber iniciado el experimento.
En general, el porcentaje de germinación en la oscuridad fue muy bajo y cuando se producía, las
plántulas se volvían cloróticas. Las semillas de 2000 sin tratamiento del endocarpo germinaron a los 48 días y su porcentaje de germinación fue del 2%; las sometidas a tratamiento empezaron a germinar a los 12 días de iniciado el experimento, con un porcentaje de germinación del 1% que se mantuvo hasta el día 24, a partir del cual no se volvió a observar emergencia de plántulas, hasta el día 48 que se registró de nuevo un 1% de germinación. Las semillas de 2001 sin tratamiento del endocarpo comenzaron a germinar a los 30 días y la germinación máxima fue del 2% a los 48 días; mientras que las sometidas a tratamiento iniciaron la emergencia a los 12 días y 36 días después se produjo una germinación del 5%.
El porcentaje de germinación en oscuridad fue muy inferior al obtenido con luz. La germinación de las semillas expuestas a la luz fue del 51% para las cosechadas en el 2000 y sometidas a tratamiento y 47% para las no tratadas. En el caso de las tratadas y sin tratar de 2001 se obtuvo un porcentaje de germinación del 53 y 40% respectivamente (Figura 2). Aunque las semillas tratadas presentaron mayor porcentaje de germinación que las no tratadas, los resultados estadísticos no mostraron diferencias significativas (F = 2,272; p = 0,115). En la
oscuridad el porcentaje de germinación obtenido en todos los casos fue muy bajo. Las semillas tratadas y no tratadas de 2000 presentaron porcentajes de 4 y 2 % respectivamente. En las semillas de 2001 el porcentaje de germinación fue de 9 % para las tratadas y 3 % para las no tratadas. Los resultados obtenidos en el análisis estadístico indican que sólo la luz está influyendo positivamente sobre la germinación
(F=150,321; p=0,000) y no existe interacción entre los efectos de los factores estudiados (F=0,0084; p=0,775).
Nodulación. Las 10 plántulas inoculadas que

Revista de Investigaciones No. 17 - Universidad del Quindío
86
crecieron en vermiculita nodularon, mientras que ninguna de las 20 plantas inoculadas, que se mantuvieron en macetas con suelo, noduló.
Las raíces de las plantas noduladas desarrollaron pocos nódulos, una media de 2,500 ± 0,342 racimos nodulares por planta, distribuidos en la raíz principal y en las raíces secundarias (Figura 3). Los racimos nodulares estaban formados por 7,200 ± 1.046 lóbulos (n = 25). La longitud de cada lóbulo era aproximadamente de 1 mm.
En la raíz principal, los racimos nodulares se encontraban situados a una distancia entre 8 y 12 cm del cuello, mientras que en las raíces secundarias estaban ubicados a una distancia entre 0,5 y 3,0 cm de la base del punto de ramificación. El valor medio de racimos por
raíz principal fue de 1,100 ± 0,261 (n = 10) y por raíz lateral, de 1,400 ± 0,298 (n = 10). No se encontraron diferencias significativas
entre el número de racimos nodulares de la raíz principal y lateral (F = 0,717, P = 0,408). Las plantas control sembradas en vermiculita no nodularon.
Crecimiento. La Tabla 2 recoge los resultados del ANOVA para los parámetros de crecimiento y nutrientes de las plantas que crecieron en dos tipos de sustrato (suelo y vermiculita) y sometidas a tratamiento (con inóculo y control). Las plantas con inóculo y control que crecieron en suelo no se diferenciaron significativamente
con respecto a los parámetros de crecimiento, por esto la apariencia de las plantas fue similar (Figura 4), mientras que las noduladas en vermiculita presentaron valores de RGR, NAR, SLA Y SWR superiores a los de los individuos control que crecieron en vermiculita (Tabla 3). Esto se refleja en el aspecto diferente de las
plantas, como se puede observar en la figura 5.
En las plantas control, las que crecieron en suelo presentaron una RWR inferior a la de las que
crecieron en vermiculita, pero en las primeras los valores de RGR, NAR, LAR, SLA, LAR y SWR fueron superiores a los de las plantas control en vermiculita. En los ejemplares inoculados, los que crecieron en suelo exhibieron valores de LAR y LWR superiores a los de los plantados en vermiculita, pero la RWR de estos últimos fue mayor que la de los ejemplares plantados en suelo.
La concentración de nitrógeno y el porcentaje de carbono diferían significativamente entre las
plantas con inóculo y control que crecieron en vermiculita. Se registró un valor mayor en las inoculadas con respecto a las plantas control. Los ejemplares, control e inoculados, que crecieron en suelo no presentaron diferencias en las concentraciones de estos nutrientes. Los nodulados en suelo y vermiculita no diferían significativamente, pero las plantas control en
suelo tuvieron concentraciones de nitrógeno y carbono mayores que los controles plantados en vermiculita.
En la relación C/N se encontró algo similar a lo anterior, pero la relación C/N de las plantas control que crecieron en vermiculita fue mayor que la de las plantadas en suelo.
Al comparar la eficiencia en el uso de nutrientes
(NUE) de las plantas que crecieron en suelo y vermiculita para un mismo tratamiento, sólo se encontraron diferencias significativas entre
los individuos control de los dos sustratos. El uso de eficiencia de nitrógeno (media ± error
estándar) fue mayor en las plantas control (0,325 ± 0,036 mg dia-1) que crecieron en suelo que en las plantadas en vermiculita (0,070 ± 0,004 mg dia-1). No se encontraron diferencias entre los inoculados y los controles que crecieron en suelo, mientras que las plantas control e inoculadas en vermiculita diferían significativamente.

Revista de Investigaciones No. 17 - Universidad del Quindío
87
DISCUSIÓN
Los resultados de la germinación de E.
angustifolia obtenidos en condiciones de laboratorio indican que las semillas cosechadas en 2000 y 2001 conservaban su viabilidad puesto que la respuesta de la germinación fue positiva. Este resultado concuerda con el encontrado por otros investigadores, quienes señalaron que las semillas de esta especie eran viables más de 3 años (Pearce y Smith, 2001; Young y Young, 1992). Mantener el potencial germinativo durante mucho tiempo favorece la supervivencia a largo plazo de E. angustifolia, ya que la capacidad de sus semillas para permanecer latentes, pero viables, hasta que las condiciones ambientales favorezcan la germinación y establecimiento de las plántulas, es un aspecto interesante que puede relacionarse con las posibilidades de colonización de nuevos espacios y, en definitiva, de su mantenimiento
como especie en el futuro.
La respuesta de las semillas de E. angustifolia a la luz fue positiva, ya que se obtuvo un porcentaje de germinación entre 40 y 53 % y las semillas germinaron con regularidad a partir del día 12, en que aparecieron los primeros brotes, hasta el día 35, en que finalizo el
experimento. Los resultados registrados en este estudio difieren de los de Zamorano (1988),
quien encontró un porcentaje de germinación del 25% y una ocurrencia de brotes distribuida de manera muy irregular en un período de 3 meses. La fotosensibilidad de las semillas de E. angustifolia, también fue comprobada por Shafroth et al. (1995), quienes hallaron que esta especie germinaba bien bajo la radiación solar directa, así como con un 89% de sombra, siempre y cuando existieran buenas condiciones de humedad.
El bajo porcentaje de plantas germinadas en
condiciones de oscuridad (2 - 9%) puede explicar la ausencia de plántulas bajo condiciones naturales (Girón, 2004) ya que la presencia de una cubierta cerrada de herbáceas posiblemente esté inhibiendo la germinación, debido al efecto de sombra producido por el follaje. Es posible también, que las semillas al caer al suelo queden cubiertas por la hojarasca, de las especies del sotobosque y de los árboles de E. angustifolia, que actúa como barrera al interceptar la luz solar, produciendo así, un efecto negativo sobre las semillas de E. angustifolia que son fotosensibles.
No se hallaron diferencias significativas en la
germinación de las semillas cuyo endocarpo fue tratado previamente y las no tratadas. Hogue y LaCroix (1970) no consiguieron que germinaran semillas sin tratar, pero con aquellas a las que les practicaron una incisión o corte registraron un porcentaje de germinación muy similar (58%) al encontrado en este estudio. Posiblemente, el lavado efectuado a las semillas, antes de proceder a la escarificación mecánica,
influyó sobre la germinación y por esto no se
hallaron diferencias significativas entre los
tratamientos de la testa. Como es sabido, las semillas tienen sustancias hidrosolubles que inhiben la germinación. Hamilton y Carpenter (1976) afirman que la testa y endocarpo de las
semillas de E. angustifolia poseen cumarina que produce un efecto inhibitorio. Por consiguiente, si esta sustancia se elimina, ya sea por lavado practicado en condiciones de laboratorio o por agua lluvia en condiciones de campo, se puede producir el brote de las semillas.
Los resultados de nodulación no fueron los esperados, puesto que las plántulas inoculadas y control, trasplantadas a macetas con suelo recogido cerca de los árboles de E. angustifolia, no nodularon. Teniendo en cuenta que: La nodulación de esta especie en campo es la que

Revista de Investigaciones No. 17 - Universidad del Quindío
88
habitualmente se encuentra en estos casos; que Frankia vive saprofíticamente en el suelo después de la necrosis de los nódulos hasta que, de nuevo, se introduce en otro huésped (Akkermans y van Dijk, 1981; Jeong y Myrold, 2001); que la distribución de Frankia
está relacionada, entre otros factores, con la distribución de las plantas huésped (Lie et al., 1984), y que, incluso, coloniza suelos donde no existe la planta huésped (Rodríguez-Barrueco, 1968; Wollum et al., 1968; Bermúdez de Castro et al., 1976; Smolander y Sundman, 1987; Arveby y Huss-Danell, 1988; Sprent y Parsons, 2000; Gtari et al., 2004), cabría esperar que el suelo recogido cerca del tronco de los árboles de E. angustifolia indujera nódulos en las plántulas. Sin embargo no fue así, ya que ninguna de las plántulas que crecieron en macetas con este suelo noduló. Bermúdez de Castro et al., (1976) plantean que la nodulación desigual, e incluso nula, de las raíces de plantas mantenidas en suelo recogido cerca de ejemplares nodulados, en los que se supone que se encuentra el endofito, es un hecho que se detecta en este
tipo de experimentos. Además, Smolander y Sundman (1987) encontraron que la nodulación era muy desigual, cuando inocularon plántulas de híbridos de Alnus incana (L.) Moench. x A. glutinosa con una suspensión de suelo recogido a diferentes distancias de árboles de aliso, y que uno de los suelos no indujo nódulos en las plántulas, lo que coincide con los resultados obtenidos en este estudio.
Se sabe que los factores edáficos, como pH,
salinidad, concentración de nitrógeno, fósforo, potasio, concentración de oligoelementos, inhiben parcial o totalmente la nodulación cuando se encuentran cerca de los límites de los intervalos de tolerancia (Griffiths y McCormick,
1984; Smolander et al., 1988; Crannell et al., 1994). Por eso, los experimentos en los que se determina la capacidad noduladora de los
suelos, directamente, creciendo plantas en ellos, como en este caso, el de Bermúdez de Castro et al. (1976), Huss-Danell y Frej (1986) y otros, corren el peligro de potenciar el efecto negativo de algún factor edáfico, al confinar una muestra
relativamente pequeña de suelo en un maceta e infravalorar el número de unidades infectivas de Frankia que verdaderamente hay en dicho suelo.
Ninguno de los autores citados explica convenientemente por qué un suelo determinado se muestra ineficaz para inducir nódulos: Si
es por ausencia de Frankia, por haber pocas unidades infectivas en el suelo o por diversas circunstancias que impiden que Frankia
llegue hasta las raíces. Una de las razones que puede justificar la ineficacia del suelo
utilizado en este estudio para inducir nódulos es, posiblemente, el enriquecimiento del suelo con nitrógeno procedente de la descomposición de la hojarasca de E. angustifolia (Bermúdez de Castro et al., 1990; Domenach et al., 1994; Simons Seastedt, 1999; Girón, 2004), ya que la concentración de nitrógeno estaba entre 1.6 y 1.8 %. Varios autores han señalado que un suelo rico en nitrógeno inhibe la formación de nódulos y la actividad nitrogenásica, debido a que las plantas prefieren absorber compuestos
nitrogenados directamente del suelo, lo que significa un gasto energético menor que cuando
establecen simbiosis con microorganismos fijadores de nitrógeno atmosférico (Hardarson
et al., 1988; Huss-Danell, 1997). En este sentido, Streeter (1988) señala que el proceso de infección y desarrollo de nódulos en plántulas de leguminosas, sembradas en un sustrato enriquecido con nitrógeno, se inhibe a concentraciones altas de este elemento. Un efecto similar también se ha observado en plantas actinorrícicas (MacConnel y Bond, 1957; Bond y Mackintosh, 1975; Ingestad, 1980; Kohls y Baker, 1989; Thomas y Berry, 1989; Arnone et

Revista de Investigaciones No. 17 - Universidad del Quindío
89
al., 1994). Rodríguez-Barrueco (1971) halló que la nodulación de E. angustifolia era sensible a la presencia de nitrógeno, inhibiéndose cuando las plántulas se colocaban en una solución de 20 mg de nitrógeno por litro o más. Aunque ésta, quizás, no es razón suficiente para explicar el
fallo en la nodulación, puesto que otros autores encontraron que las plántulas de E. angustifolia producían nódulos cuando crecían en sustratos enriquecidos con diferentes concentraciones de nitrógeno (Kohls y Baker, 1989; Zitzer y Dawson, 1992). Además, se han obtenido resultados positivos en nodulación de plántulas sembradas en suelos recogidos cerca de los árboles de E. angustifolia y suelos donde no se encuentra la planta huésped (Schmitz et al., 1990; Gtari et al., 2004). Lo que lleva a plantear, que posiblemente existen otras razones, además de la fertilidad del suelo, que afectan la simbiosis de E. angustifolia con el endofito.
Los daños producidos en las raíces y el estrés de adaptación de las plántulas al trasplantarlas del semillero a las macetas, sería otra razón que podría explicar la falta de nodulación en las plántulas que crecieron en suelo. Cuando las raíces se estropean, la planta incrementa su tasa respiratoria y emplea los factores de crecimiento para restituir sus raíces (Milthorpe y Moorby, 1988). Además, disminuye la absorción de agua y se produce una reducción del crecimiento radical (Kramer, 1985). Esto perturbó probablemente de manera desigual la nodulación de las plantas inoculadas, ya que las raíces encuentran más dificultad al crecer en
suelo que en vermiculita.
La textura del suelo también afecta al crecimiento de las raíces y, en este caso, posiblemente a la nodulación. En suelos arcillosos y limosos, como el utilizado en este estudio, cuando están parcialmente saturados de agua, se dan procesos de humectación y desecación que compactan el
suelo e inciden negativamente sobre las raíces. Este efecto se potencia cuando se ponen en macetas de tamaño reducido y se someten a riegos periódicos. La deformación física del suelo y la compactación tienden a disminuir la cantidad de agua y de nutrientes susceptibles de ser captados por las raíces (Porta et al., 1999), lo que dificultaría, e incluso impediría quizá la
nodulación de las plántulas de E. angustifolia.
La salinidad también pudo incidir negativamente en el establecimiento de la simbiosis entre las plántulas de E. angustifolia y Frankia. Aunque en otras especies fijadoras de nitrógeno, como
Vicia faba L. y C. equisetifolia se demostró que las concentraciones de sales altas afectaban al crecimiento de las plantas y al peso de los nódulos, no se comprobó que la salinidad del sustrato inhibiera parcial o totalmente la infección y nodulación (NG, 1987; Cordovilla et al., 1999). Sin embargo, Chiharu et al. (2000) encontraron que concentraciones altas de sal inhibían el crecimiento de Frankia. Así, que no se puede descartar un efecto de la salinidad sobre la nodulación de las plántulas de E.
angustifolia.
A diferencia de las plántulas inoculadas en suelo que no lograron nodular, las inoculadas en vermiculita sí nodularon. Los nódulos se observaron en la raíz principal y raíces laterales. Esta distribución también se observó en Discaria trinervis (Hook. y Arn.) Reiche. En ambas especies, el proceso de infección ocurre por penetración apoplástica. El filamento hifal
se abre paso a través de la cutícula y penetra a los espacios intercelulares del cortéx de la raíz por la lamela media entre dos células epidérmicas adyacentes (Miller y Baker, 1985; Valverde y Wall, 1999). En el caso de E. angustifolia, los racimos nodulares observados tenían forma coraloide, se dividían en lóbulos alargados y su longitud sólo alcanzaba 1 mm. Se debe

Revista de Investigaciones No. 17 - Universidad del Quindío
90
considerar que las plántulas tenían solamente 6 meses cuando se cosecharon y que el tamaño de los lóbulos nodulares va a depender, en parte de la edad y que, según Huss-Danell (1997), los racimos nodulares producidos por Frankia
pueden variar en longitud desde unos pocos milímetros hasta varios centímetros cuando el sustrato está bien o pobremente aireado. El número de racimos nodulares formados no fue muy grande. Rodríguez-Barrueco (1971) registró, para esta misma especie, un promedio de 44 nódulos por planta y Zitzer y Dawson (1992) encontraron un valor medio de 19.8 nódulos.
La tasa de crecimiento de los ejemplares plantados en suelo y los nodulados en vermiculita fue alta comparada con la de los controles en vermiculita. Las plantas que crecieron en suelo incrementaron su tamaño debido a la disponibilidad de nitrógeno en el suelo y las que nodularon en vermiculita obtuvieron el nitrógeno necesario para su crecimiento de la simbiosis con Frankia. La fracción del peso seco total que las plantas destinaron a la producción de hojas fue mayor en las plantadas en suelo que en las que crecieron en vermiculita, mientras que hubo una mayor fracción del peso seco total destinado a la raíz en estas últimas, debido a que crecieron en un sustrato con déficit
de nutrientes, especialmente nitrógeno. Las plantas que se desarrollan en un sustrato pobre en este elemento distribuyen mayor biomasa a las raíces que al tallo, lo que constituye una respuesta fenotípica a la escasez de nutrientes (Chapin, 1980; Aerts y Chapin, 2000).
Las plantas en suelo tuvieron una ventaja sobre las noduladas en vermiculita, ya que desde el inicio pudieron invertir más energía en el desarrollo del follaje y menos en el aparato radicular porque disponían de suficiente nitrógeno en
el suelo. Por el contrario, las que crecieron en
vermiculita sin nitrógeno desarrollaron más las raíces como un mecanismo de respuesta ante la ausencia de nutrientes. Las plantas que crecen con poca disponibilidad de nutrientes generalmente son pequeñas e invierten más en biomasa radicular (Poorter y Nagel, 2000; Navas y Garnier, 2002). Cuando las inoculadas en vermiculita lograron fijar nitrógeno atmosférico,
alcanzaron una tasa de crecimiento muy similar a la de los individuos que crecieron en suelo. Por esta razón, la concentración de nitrógeno foliar de las plantas crecidas en suelo fue muy parecida a la de las hojas de E. angustifolia en condiciones naturales (Girón, 2004), aspecto que contribuyó al incremento de la biomasa, encontrándose valores relativamente altos de RGR, NAR, LAR y SLA. Las características foliares como concentración de nitrógeno y SLA altos son determinantes en la productividad potencial de las plantas y permiten que tengan más plasticidad en sus respuestas a los cambios ambientales (Reich et al., 1992; Hunt y Cornelissen, 1997; Moreira dos Santos Jr. et
al., 2006). Los individuos control sembrados en vermiculita presentaron una concentración de nitrógeno baja, y, por esta razón, los valores de RGR, NAR, LAR y SLA fueron también bajos. Se sabe que las hojas con poca concentración de nitrógeno tienen una tasa de asimilación disminuida que afecta negativamente a la tasa de crecimiento (Hirose y Werger,1987; Reich et
al., 1992; Lawlor, 2002).
Las plantas de E. angustifolia utilizaron eficientemente el nitrógeno disponible en el
suelo (plantas en macetas con suelo) o el fijado
por Frankia (plantas noduladas en macetas con vermiculita), excepto las control que crecieron en vermiculita, debido a la ausencia de nutrientes en el suelo.
Los resultados obtenidos en este trabajo reflejan
la productividad potencial de las plantas de E.

Revista de Investigaciones No. 17 - Universidad del Quindío
91
angustifolia y comprueban su adaptación para crecer en suelos con fertilidad tanto baja como
alta, aspectos que influyen en la efectividad de
esta especie para colonizar áreas diferentes.
BIBLIOGRAFÍA
Aerts, R. y Chapin, F. S.III. (2000). “The mineral nutrition of wild plants revisited: A re-1. evaluation of processes and patterns”. En Advances in Ecological Research, 30, 1-67.
Akkermans, A. D. L. y van Dijk, C. (1981). “Non-leguminous root-nodule symbioses with 2. actinomycetes and Rhizobium”. En: Broughton, W. J. (ed.), Nitrogen fixation ecology (57-
103). Oxford University Press, Oxford.
Allue Andrade, J. L. (1990). Atlas fitoclimático de España: Taxonomias. Ministerio de 3. Agricultura Pesca y Alimentación. Instituto Nacional de Investigaciones Agrarias, Madrid.
Arnone, J. A., Kohls, S. J. y Baker, D. D. (1994). “Nitrate effects on nodulation and nitrogenase 4. activity of actinorhizal Casuarina estudied in split-root systems”. En Soil Biology and Biochemestry, 26, 599-606.
Arveby, A.S. y Huss-Danell, K. (1988). Presence and dispersal of infective Frankia in peat and 5. meadow soils in Sweden. En Biol. Fert. Soils, 6, 39-44.
Becking, J. H. (1970). “Frankiaceae Fam. Nov. (Actinomycetales) with one new combination 6. and six new species of genus Frankia Brunchorst 1886”. En International Journal of Systematic Bacteriology, 20, 201-220.
Belcher, E. W. y Karrfalt, R. P. (1979). Improved methods for testing the viability of Russian 7. olive seed. En Journal of Seed Technology, 4, 57-64.
Bermúdez de Castro, F. (1988). “Las plantas actinorrizas y la sucesión”. En: Villar, L. (ed.), 8. Homenaje a Pedro Montserrat (435-439). Monografía del Instituto Pirenaico de Ecología 4, Jaca, España.
Bermúdez de Castro, F., Miguel, C. y Rodríguez-Barrueco, C. (1976). “A study of the capacity 9. of soil to induce nodules in Alnus glutinosa (L.) Gaertn and Myrica gale (L.) with special reference to the specificity of the endophytes”. En Annales de Microbiologie, 127a, 307-315.
Bermudez de Castro, F., Aranda, Y. y Schmitz, M. F. (1990). “Acetylene-reducing activity and 10. nitrogen inputs in a bluff of Elaeagnus angustifolia L”. En Orsis, 5, 85-89.
Bond, G. y Mackintosh, A. H. (1975). “Effect of nitrate-nitrogen on the nodule symbiosis of 11. Coriaria and Hippophaë”. En Proceedings of the Royal Society of London Serie B. Biological

Revista de Investigaciones No. 17 - Universidad del Quindío
92
Sciences, 190, 199-209.
Bremner, J. M. y Tabatabai, M. A. (1972). “Use of an ammonia electrode for determination 12. of ammonium in Kjeldahl analysis of soils”. En Communications in Soil Science and Plant Analysis, 3, 159-165.
Chapin, F. S. III. (1980). “The mineral nutrition of wild plants”. En Annual Review of Ecological 13. Systematic, 11, 233-260.
Chapin, F. S., Walker, C. L., Fastie, C. L. y Sharman, L. C. (1994). “Mechanisms of primary 14. succession following deglaciation at Glacier Bay, Alaska”. En Ecological Monograph, 64, 149-175.
Cordovilla, M. P., Ligero, F. y Lluch, C. (1999). “Effects of NaCl on growth and nitrogen 15. fixation and assimilation of inoculated and KNO3 fertilized Vicia faba L. and Pisum sativum L.
plants”. En Plant Science, 140, 127-136.
Crannell, W. K., Tanaka, Y. y Myrold, D. D. (1994). “Calcium and pH interaction on root 16. nodulation of nursery-grown red alder (Alnus rubra Bong.) seedlings by Frankia”. En Soil Biology & Biochemistry, 26(5), 607-614.
Domenach, A.M., Moiroud, A. y Jocteur-Monrozier, L. (1994). “Leaf carbon and nitrogen 17. consituents of some actinorhizal tree species”. En Soil Biology and Biochemistry, 5, 649-653.
Girón, V. M. (2004). Evolución del bosquete de Elaeagnus angustifolia L. en Valdemoro 18. (Madrid). Tesis doctoral. Departamento de Ecología, Facultad de Ciencias Biológica, Universidad Complutense de Madrid. España.
Griffiths, A. P. y McCormick, L. H. (1984). “Effects of soil acidity on nodulation ofAlnus 19. glutinosa and viability ofFrankia”. En Plant and Soil, 79(3), 429-434.
Gtari, M., Brusett, L., Skander, G., Mora, D., Boudabous, A. y Daffonchio, D. (2004). “Isolation 20. of Elaeagnus-compatible Frankia form soils collected in Tunisia”. En FEMS Microbiology Letters, 234, 349-355.
Gutiérrez Mañero, F. J. (1987). Estudios sobre la influencia de Myrica gale en la microflora 21. edáfica del ciclo del nitrógeno. Tesis doctoral. Facultad de Ciencias biológicas. Universidad
Complutense de Madrid.
Gutiérrez Mañero, F. J. y Bermúdez de Castro, F. (1983). “Modificaciones de la microflora 22. edáfica del ciclo de nitrógeno bajo Myrica gale L”. En Anales de Edafología y Agrobiología,
42, 1233-1244.

Revista de Investigaciones No. 17 - Universidad del Quindío
93
Hamilton, D. F. y Carpenter, P. L. (1976). “Regulation of seed dormancy in Elaeagnus angustifolia 23. by endogenous growth substances”. En Canadian Journal of Botany, 54(10), 1068-1073.
Hardarson, G., Zapata, F. y Danso, S. K. A. (1988). Effect on N fertilization en N2 fixation in 24. fabacean (Vicia faba L.var. minor). Abst. 7 th Int. Congress on N-nitrogen fixation, Köln. pp.
6-32.
Hibbs, D. E. y Cromack, Jr. (1990). “Actinorhizal plants in Pacific Northwest forests”. En: C. 25. R. Schwintzer y J. D. Tjepkma (eds.), The biology of Frankia and actinorhizal plants (343-363). Academic Press San Diego, California.
Hirose, T. y Werger, M. J. A. (1987). “Nitrogen use efficiency in instantaneous and daily 26. photosynthesis of leaves in the canopy of a Solidago altissima stand”. En Physiologia plantarum, 70, 215-222.
Hogue, E. J. y LaCroix, L. J. (1970). “Seed dormancy of Russian Olive (Elaeagnus angustifolia 27. L.)”. En Journal of the American Society Horticultural Science, 95, 449-452.
Hunt, R. y Cornelissen, J. H. C. (1997). “Components of relative growth rate and their 28. interrelations in 59 temperate plant species”. En New Phytology, 135, 395-417.
Huss-Danell, K. (1997). “Tansley Review Nº 93: Actinorhizal symbioses and their N2 fixation”. 29. En New Phytologist, 136, 375-405.
Huss-Danell, K. y Frej. A. K. (1986). “Distribution of Frankia in soils from forests and 30. afforestation sites in northern Sweden”. En Plant and Soil, 90, 407-418.
Ingestad, T. (1980). “Growth, nutrition, and nitrogen fixation in grey alder at varied rate of 31. nitrogen addition”. En Physiologia Plantarum, 50, 353-364.
Jeon, Soon-Chun y Myrold, D. D. (2001). “Population size and diversity of Frankia in soils of 32. Ceanothus velutinus and Douglas-fir stands”. En Soil Biology & Biochemistry, 33, 931-941.
Katz, G. L. y Shafroth, P. B. (2003). “Biology, ecology and management of Elaeagnus 33. angustifolia L. (Russian Olive) in western North America”. En Wetlands, 23(4), 763-777.
Kjeldahl, J. (1883). “Neue methode zur bestimmung des stickstoffs in organischen körpern”. 34. En Zeitschrift Analytische Chemie, 195, 171-184.
Kohls, S. J. y Baker, D. D. (1989). “Effects of substrate nitrate concentration on symbiotic 35. nodule formation in actinorhizal plants”. En Plant and Soil, 118, 171-179.
Kramer, P. J. (1985). Plant and soil water relationship. McGraw-Hill, New York.36.

Revista de Investigaciones No. 17 - Universidad del Quindío
94
Lawlor, D.W. (2002). “Carbon and nitrogen assimilation in relation to yield: mechanisms are 37. the key to understanding production systems”. En Journal of Experimental Botany, 53, 773-787.
Lie, T. A. Akkermans, A. D. L. y van Egeraat, A. W. S. M. (1984). “Natural variation in symbiotic 38. nitrogen-fixing Rhizobium and Frankia spp”. En Antonie van Leeuwenhoek, 50, 489-503.
Linnaeus, C. (1753). Species Plantarum. London. 39.
Llinares, F. D. (1990). Densidad y actividad de los microorganismos del ciclo de nitrógeno 40. bajo el dosel de Elaeagnus angustifolia L. Memoria de Licenciatura. Facultad de Ciencias Biológicas. Universidad Complutense de Madrid.
Llinares, F. D. (1993). Regulación del ciclo del nitrógeno a nivel edáfico por Elaeagnus 41. angustifolia L. y Alnus glutinosa (L.) Gaertn. Tesis doctoral. Facultad de Ciencias Biológicas. Universidad Complutense de Madrid.
Llinares, F. D., Muñoz-Mingarro, J. M., Pozuelo, B. R. y Bermúdez de Castro, F. (1993). 42. “Microbial inhibition and nitrification potential in soils incubated with Elaeagnus angustifolia
L. leaf litter”. En Geomicrobiology Journal, 11, 149-156.
MacConnell., J. T. y Bond, G. (1957). “A comparison of the effect of combined nitrogen on 43. nodulation in non-legumes and legumes”. En Plant and Soil, 8: 378-388.
Mateus, P. C. L. (1997). Interacciones del aliso Alnus acuminata H.B.K. en el sistema suelo 44. planta y su efecto en el ciclo del nitrógeno. Memoria tesis doctoral. Facultad de Ciencias Biológicas. Departamento de Ecología. Universidad Complutense. Madrid.
Miller, I. M. y Baker, D. D. (1985). “The initiation, development and structure of root nodules 45. in Elaeagnus angustifolia L. (Elaeagnaceae)”. En Protoplasma, 128, 107-119.
Milthorpe, F. L. y Moorby, J. (1988). An introduction to crop physiology. Cambridge University 46. Press, Cambridge.
Monturiol, R. F. y Alcalá del Olmo, B. L. (1990). Mapa de asociaciones de suelos de la 47. comunidad de Madrid. CSIC, Madrid.
Moreira dos Santos Jr., U., José Francisco de Carvalho Gonçalves, J. y Feldpausch, T. R. (2006). 48. Growth, leaf nutrient concentration and photosynthetic nutrient use efficiency in tropical tree
species planted in degraded areas in central Amazonia. En Forest Ecology and Management, 226, 299-309.
Moretti, A. (1993). Actinorhizal symbioses. En Gionale Botanico Italiano, 127, 434-443.49.

Revista de Investigaciones No. 17 - Universidad del Quindío
95
Navas Marie-Laure y Eric Garnier, E. (2002). “Plasticity of whole plant and leaf traits in 50. Rubia peregrina in response to light, nutrient and water availability”. En Acta Oecologica, 23, 375-383
NG. B. H. (1987). “The effects of salinity on growth, nodulation and nitrogen fixation of 51. Casuarina equisetifolia”. En Plant and Soil, 103, 123-125.
Pearce, C. M. y Smith, D. G. (2001). “Plains cottonwood’s last stand: Can it survive invasion 52. of russian olive onto the Milk River, Montana floodplain?” En Environmental Management,
28(5), 623-637.
Pérez Hernández, M. C. (1996). Entradas biológicas de nitrógeno en un bosque ripario. Tesis 53. doctoral. Facultad de Ciencias Biológicas. Universidad Complutense, Madrid.
Poorter, H. y Nagel, O. (2000). “The role of biomass allocation in the growth response of plants 54. to different levels of light, CO2, nutrients and water: a quantitative review”. En Australian Journal of Plant Physiology, 27, 595-607.
Porta, J., López-Acevedo, M. y Roquero de Laburu, C. (1999). Edafología para la agricultura y 55. el medio ambiente. Ed. Mundi-Prensa, Madrid.
Pozuelo González, J. M. (1991). Estudio de grupos funcionales de microorganismos edáficos 56. en la rizosfera de Alnus glutinosa (L.) Gaertn. Tesis doctoral. Universidad Complutense de Madrid.
Pozuelo González, J. M., Gutiérrez Mañero, F. J., Llinares, F.Probanza, A. y Bermúdez de 57. Castro, F. (1992). “Densidad y actividad de microorganismos del ciclo de carbono bajo el dosel de Myrica gale L”. En Microbiología SEM, 8, 32-39.
Reich, P. B., Walters, M. B. y Ellsworth, D. S. (1992). “Leaf life-span in relation to leaf, plant, 58. and stand characteristics among diverse ecosystems”. En Ecological Monographs, 62, 365-392.
Rikli, M. (1943). Das pflanzankleid der mittelmeerländer, I. Berna: Hans Huber. 3 V. con 10 59. fasc.
Rodríguez-Barrueco, C. (1968). “The occurrence of the root-nodule endophyte of Alnus 60. glutinosa and Myrica gale in soils”. En Journal of General Microbiology, 52, 189-194.
Rodríguez-Barrueco, C. (1971). “Efecto del nitrógeno combinado sobre el desarrollo nodular 61. y crecimiento de Elaeagnus angustifolia L”. En Anales de Edafología y Agrobiología, 30, 149-161.

Revista de Investigaciones No. 17 - Universidad del Quindío
96
Ron, M. E. (1971). “Sobre el carácter subespontáneo de Elaeagnus angustifolia L”. En Anales 62. de la Real Academia de Farmacia, 37, 229-240.
Schmitz M. F., Aranda, Y., Esteban, M. L. y Bermúdez de Castro, F. (1990). “Nodulación de 63. Elaeagnus angustifolia L. en el bosquete de Valdemoro (Madrid)”. En Ecología, 4, 121-129.
Shafroth, P. B., Auble, G. T. y Scott, M. L. (1995). “Germination and establishment of native 64. plains cottonwood (Populus deltoides Marshall subsp. monillifera) and the exotic Russian-olive (Elaeagnus angustifolia L.)”. En Conservation Biology, 9, 1169-1175.
Simons, S. B. y Seastedt, T. R. (1999). “Decomposition and nitrogen release from foliage 65. of cottonwood (Populus deltoides) and Russian-olive (Elaeagnus angustifolia) in a riparian ecosystem”. En Southwestern Naturalist, 44, 256-260.
Smolander, A. y Sundman, V. (1987). “Frankia in acid soils of forest devoid in actinorhizal 66. plants”. En Physiological Plantarum, 70, 297-303.
Smolander, A., Van Dijk, C. y Sundman, V. (1988). “Survival of Frankia strains introduced into 67. soil”. En Plant and Soil, 106(1), 65-72.
Sprent, J. I. y Parsons, R. (2000). “Nitrogen fixation in legume and non-legume trees”. En Field 68. Crops Research, 65, 183-196.
Streeter, J. (1988). “Inhibition of legume nodule formation and N2 fixation by nitrate. CRC 69. Crit”. En Review Plant Science, 7, 1-23.
Tarrant, R. F. y Trappe, J. M. (1971). “The role of Alnus in improving the forest environment”. 70. En Plant and Soil, 76, 287-295.
Thomas, K. A. y Berry, A. M. (1989). “Effects on continuous nitrogen application and nitrogen 71. preconditioning on nodulation and growth of Ceanothus griseus var horizontalis”. En Plant and Soil, 118, 181-187.
Valverde, C. y Wall., L. G. (1999). “Time course of nodule development in Discaria trinervis 72. (Rhamnaceae)-Frankia symbioses”. En New Phytologist, 141, 345-354.
Zamorano, I. V. (1988). Elaeagnus angustifolia L. en la Villa de Madrid: Introducción a un 73. estudio ecofisiológico. Tesina Departamento de Ecología, Facultad de Ciencias Biológicas,
Universidad Complutense, Madrid.
Walkley, A. (1935). “An examination of a rapid method for determining organic carbon and 74. nitrogen in soils”. En Journal of Agricultural Science, 25, 598-609.
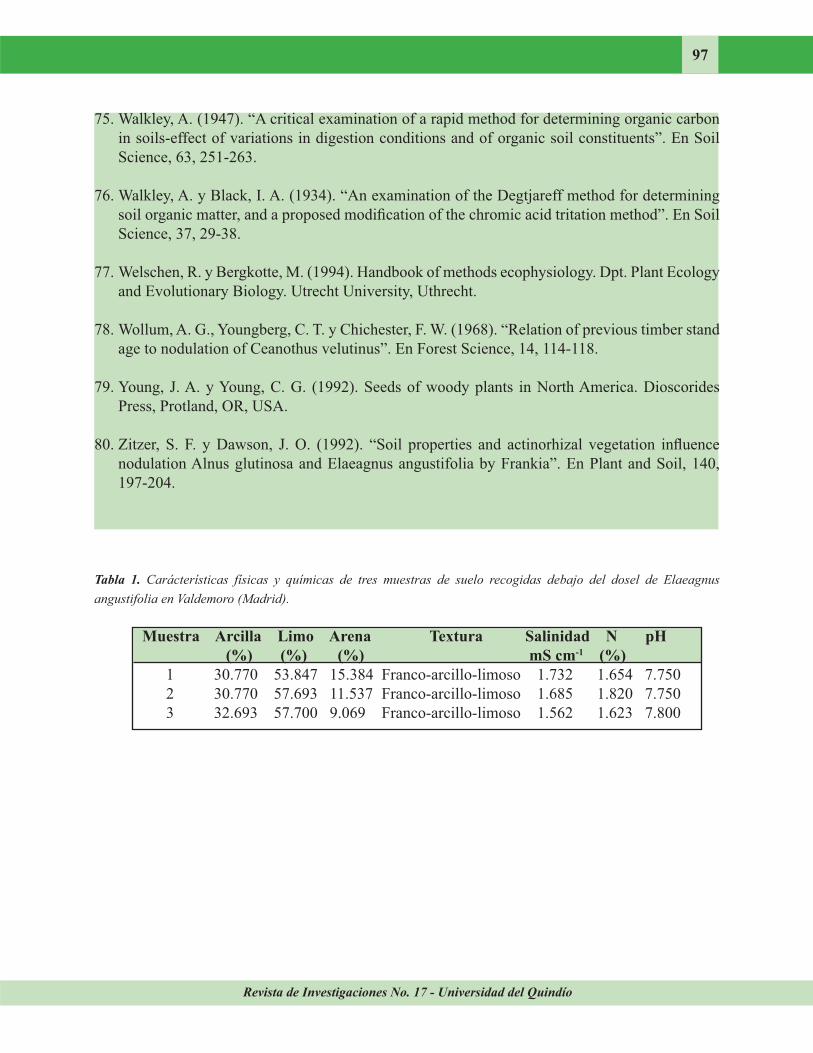
Revista de Investigaciones No. 17 - Universidad del Quindío
97
Tabla 1. Carácterísticas físicas y químicas de tres muestras de suelo recogidas debajo del dosel de Elaeagnus
angustifolia en Valdemoro (Madrid).
Muestra Arcilla Limo Arena Textura Salinidad N pH (%) (%) (%) mS cm-1 (%) 1 30.770 53.847 15.384 Franco-arcillo-limoso 1.732 1.654 7.750 2 30.770 57.693 11.537 Franco-arcillo-limoso 1.685 1.820 7.750 3 32.693 57.700 9.069 Franco-arcillo-limoso 1.562 1.623 7.800
Walkley, A. (1947). “A critical examination of a rapid method for determining organic carbon 75. in soils-effect of variations in digestion conditions and of organic soil constituents”. En Soil Science, 63, 251-263.
Walkley, A. y Black, I. A. (1934). “An examination of the Degtjareff method for determining 76. soil organic matter, and a proposed modification of the chromic acid tritation method”. En Soil
Science, 37, 29-38.
Welschen, R. y Bergkotte, M. (1994). Handbook of methods ecophysiology. Dpt. Plant Ecology 77. and Evolutionary Biology. Utrecht University, Uthrecht.
Wollum, A. G., Youngberg, C. T. y Chichester, F. W. (1968). “Relation of previous timber stand 78. age to nodulation of Ceanothus velutinus”. En Forest Science, 14, 114-118.
Young, J. A. y Young, C. G. (1992). Seeds of woody plants in North America. Dioscorides 79. Press, Protland, OR, USA.
Zitzer, S. F. y Dawson, J. O. (1992). “Soil properties and actinorhizal vegetation influence 80. nodulation Alnus glutinosa and Elaeagnus angustifolia by Frankia”. En Plant and Soil, 140, 197-204.
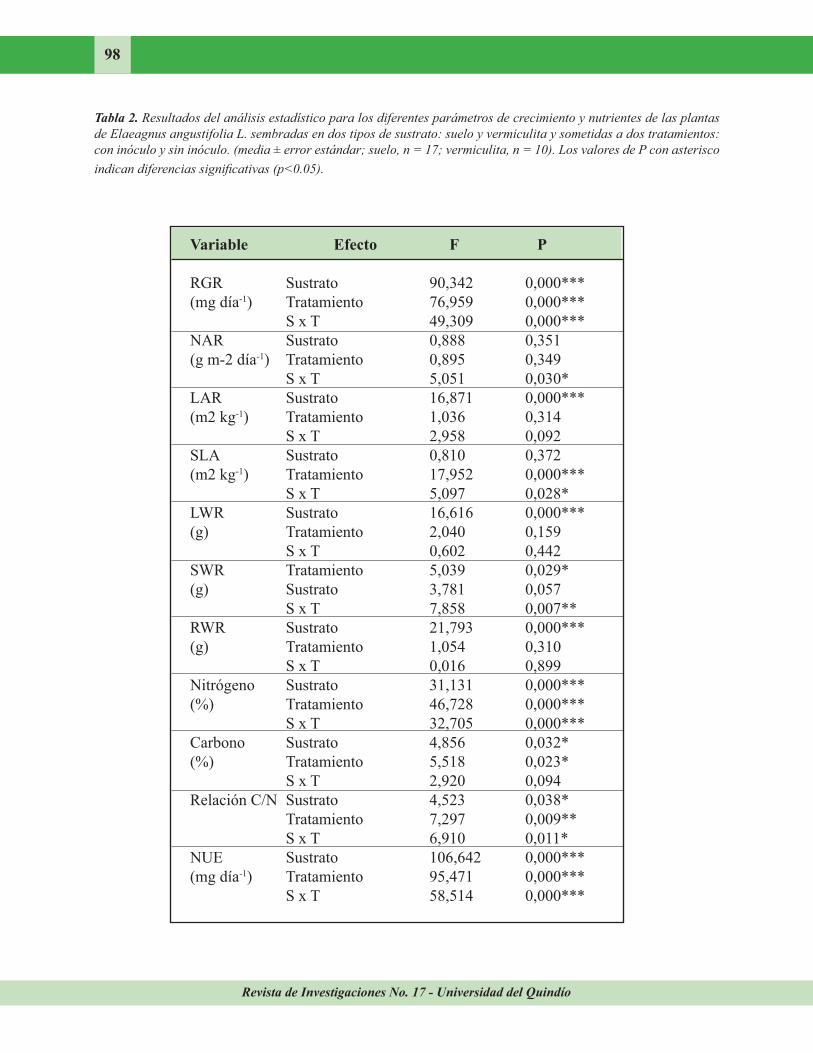
Revista de Investigaciones No. 17 - Universidad del Quindío
98
Tabla 2. Resultados del análisis estadístico para los diferentes parámetros de crecimiento y nutrientes de las plantas
de Elaeagnus angustifolia L. sembradas en dos tipos de sustrato: suelo y vermiculita y sometidas a dos tratamientos:
con inóculo y sin inóculo. (media ± error estándar; suelo, n = 17; vermiculita, n = 10). Los valores de P con asterisco
indican diferencias significativas (p<0.05).
Variable Efecto F P
RGR Sustrato 90,342 0,000*** (mg día-1) Tratamiento 76,959 0,000*** S x T 49,309 0,000*** NAR Sustrato 0,888 0,351 (g m-2 día-1) Tratamiento 0,895 0,349 S x T 5,051 0,030* LAR Sustrato 16,871 0,000*** (m2 kg-1) Tratamiento 1,036 0,314 S x T 2,958 0,092 SLA Sustrato 0,810 0,372 (m2 kg-1) Tratamiento 17,952 0,000*** S x T 5,097 0,028* LWR Sustrato 16,616 0,000*** (g) Tratamiento 2,040 0,159 S x T 0,602 0,442 SWR Tratamiento 5,039 0,029* (g) Sustrato 3,781 0,057 S x T 7,858 0,007** RWR Sustrato 21,793 0,000*** (g) Tratamiento 1,054 0,310 S x T 0,016 0,899 Nitrógeno Sustrato 31,131 0,000*** (%) Tratamiento 46,728 0,000*** S x T 32,705 0,000*** Carbono Sustrato 4,856 0,032* (%) Tratamiento 5,518 0,023* S x T 2,920 0,094 Relación C/N Sustrato 4,523 0,038* Tratamiento 7,297 0,009** S x T 6,910 0,011* NUE Sustrato 106,642 0,000*** (mg día-1) Tratamiento 95,471 0,000*** S x T 58,514 0,000***
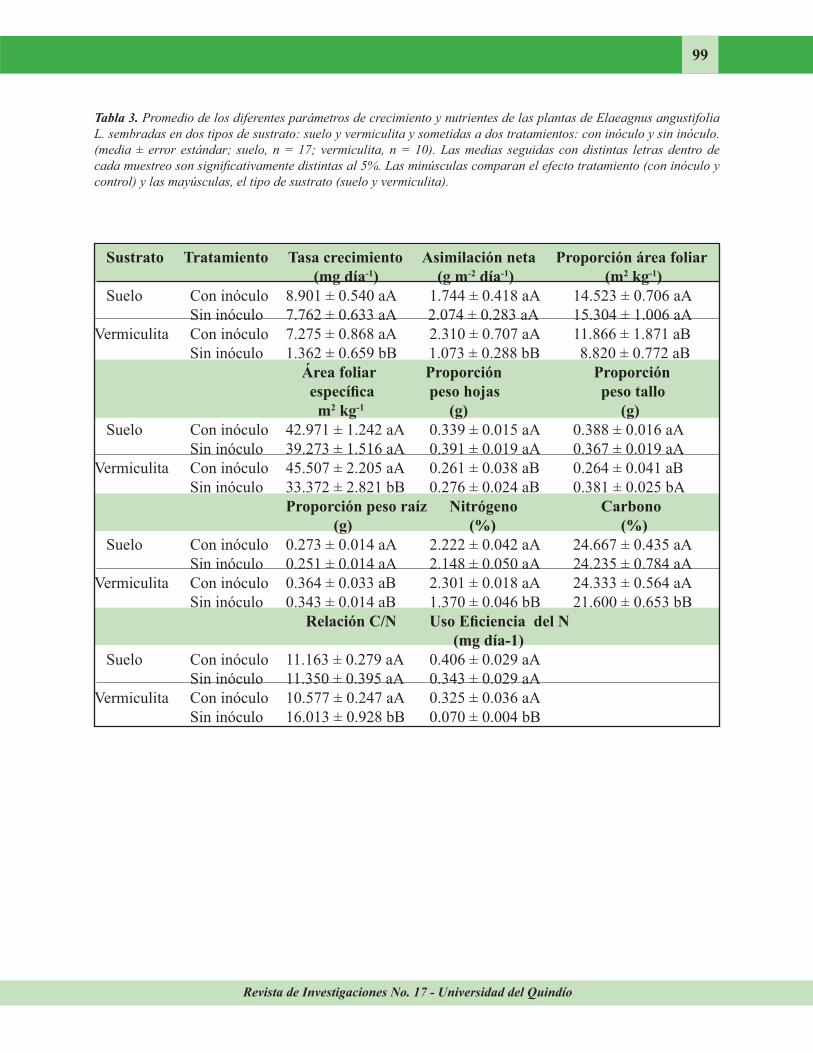
Revista de Investigaciones No. 17 - Universidad del Quindío
99
Tabla 3. Promedio de los diferentes parámetros de crecimiento y nutrientes de las plantas de Elaeagnus angustifolia
L. sembradas en dos tipos de sustrato: suelo y vermiculita y sometidas a dos tratamientos: con inóculo y sin inóculo.
(media ± error estándar; suelo, n = 17; vermiculita, n = 10). Las medias seguidas con distintas letras dentro de
cada muestreo son significativamente distintas al 5%. Las minúsculas comparan el efecto tratamiento (con inóculo y
control) y las mayúsculas, el tipo de sustrato (suelo y vermiculita).
Sustrato Tratamiento Tasa crecimiento Asimilación neta Proporción área foliar (mg día-1) (g m-2 día-1) (m2 kg-1) Suelo Con inóculo 8.901 ± 0.540 aA 1.744 ± 0.418 aA 14.523 ± 0.706 aA Sin inóculo 7.762 ± 0.633 aA 2.074 ± 0.283 aA 15.304 ± 1.006 aAVermiculita Con inóculo 7.275 ± 0.868 aA 2.310 ± 0.707 aA 11.866 ± 1.871 aB Sin inóculo 1.362 ± 0.659 bB 1.073 ± 0.288 bB 8.820 ± 0.772 aB Área foliar Proporción Proporción específica peso hojas peso tallo m2 kg-1 (g) (g) Suelo Con inóculo 42.971 ± 1.242 aA 0.339 ± 0.015 aA 0.388 ± 0.016 aA Sin inóculo 39.273 ± 1.516 aA 0.391 ± 0.019 aA 0.367 ± 0.019 aAVermiculita Con inóculo 45.507 ± 2.205 aA 0.261 ± 0.038 aB 0.264 ± 0.041 aB Sin inóculo 33.372 ± 2.821 bB 0.276 ± 0.024 aB 0.381 ± 0.025 bA Proporción peso raíz Nitrógeno Carbono (g) (%) (%) Suelo Con inóculo 0.273 ± 0.014 aA 2.222 ± 0.042 aA 24.667 ± 0.435 aA Sin inóculo 0.251 ± 0.014 aA 2.148 ± 0.050 aA 24.235 ± 0.784 aAVermiculita Con inóculo 0.364 ± 0.033 aB 2.301 ± 0.018 aA 24.333 ± 0.564 aA Sin inóculo 0.343 ± 0.014 aB 1.370 ± 0.046 bB 21.600 ± 0.653 bB Relación C/N Uso Eficiencia del N (mg día-1) Suelo Con inóculo 11.163 ± 0.279 aA 0.406 ± 0.029 aA Sin inóculo 11.350 ± 0.395 aA 0.343 ± 0.029 aA Vermiculita Con inóculo 10.577 ± 0.247 aA 0.325 ± 0.036 aA Sin inóculo 16.013 ± 0.928 bB 0.070 ± 0.004 bB
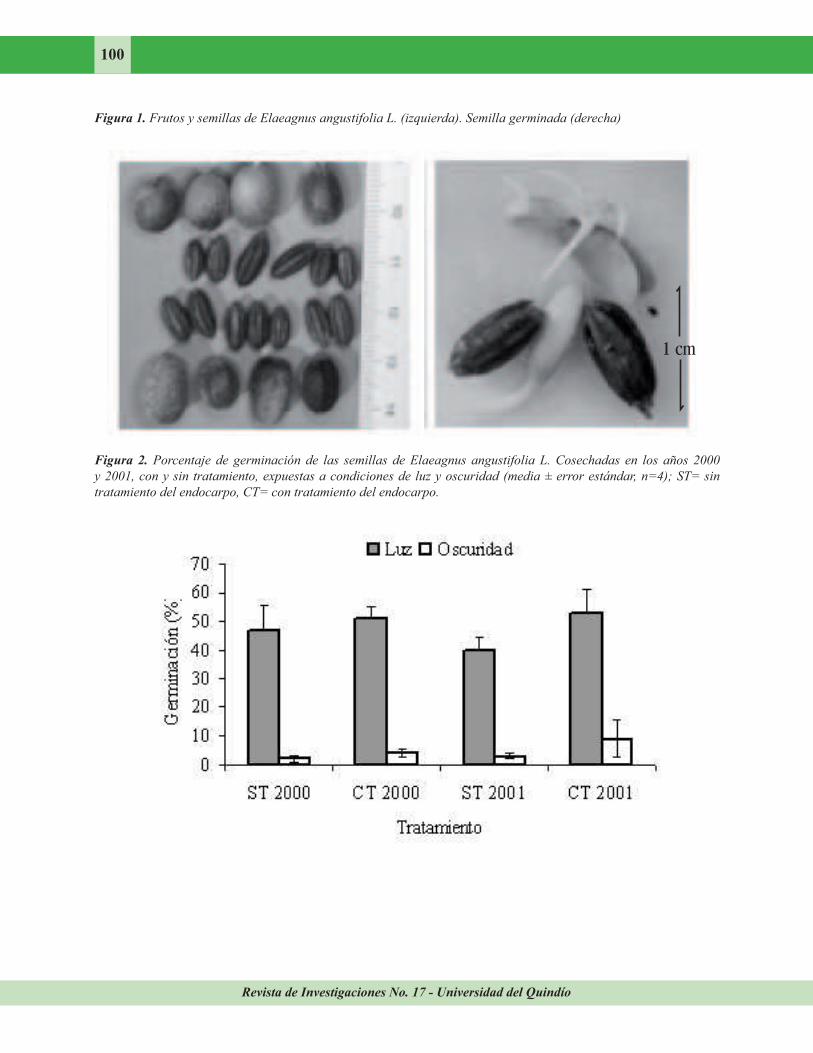
Revista de Investigaciones No. 17 - Universidad del Quindío
100
Figura 1. Frutos y semillas de Elaeagnus angustifolia L. (izquierda). Semilla germinada (derecha)
1 cm
Figura 2. Porcentaje de germinación de las semillas de Elaeagnus angustifolia L. Cosechadas en los años 2000
y 2001, con y sin tratamiento, expuestas a condiciones de luz y oscuridad (media ± error estándar, n=4); ST= sin
tratamiento del endocarpo, CT= con tratamiento del endocarpo.

Revista de Investigaciones No. 17 - Universidad del Quindío
101
Figura 3. Raíces de plántulas de Elaeagnus angustifolia L. con racimos nodulares, los cuales aparecen encerrados
en un círculo rojo.
Figura 4. Plántulas de Elaeagnus angustifolia L. el ejemplar de la izquierda fue inoculado en el suelo y el de la
derecha corresponde a uno de los controles en suelo.
5 cm{

Revista de Investigaciones No. 17 - Universidad del Quindío
102
Figura 5. Plántulas de Elaeagnus angustifolia L. el ejemplar de la izquierda fue inoculado en vermiculita y el de la
derecha corresponde a uno de los controles en vermiculita.
5 cm{