análise da interação dos protozoários parasitas leishmania (l.)...
TRANSCRIPT

Débora Barreiros Petrópolis
Análise da interação dos protozoários parasitas Leishmania (L.) amazonensis e Tritrichomonas
foetus com a glicoproteína da matriz extracelular laminina-1.
Dissertação submetida ao Instituto de Biofísica Carlos Chagas Filho, da
Universidade Federal do Rio de Janeiro, visando a obtenção do grau em Mestre em Ciências Biológicas (Biofísica)
Universidade Federal do Rio de Janeiro Centro de Ciências da Saúde Instituto de Biofísica Carlos Chagas Filho 2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Petrópolis, Débora Barreiros Análise da Interação dos protozoários parasitas Tritrichomonas foetus e Leishmania (L.) amazonensis com a glicoproteína da matriz extracelular laminina-1 / Débora Barreiros Petrópolis – Rio de Janeiro: UFRJ / IBCCF, 2007. xi, 90f.: il.; 31 cm. Orientador: Fernando Costa e Silva Filho Dissertação (mestrado) – UFRJ, IBCCF, Pós-graduação em Ciências Biológicas (Biofísica), 2007. Referências bibliográficas: 78-92 1. Matriz extracelular. 2. Tritrichomonas foetus. 3.
Leishmania (L.) amazonensis. 4. Laminina-1. 5. Relação parasita / hospedeiro
I. Silva-Filho, Fernando Costa. II. Universidade Federal do Rio de Janeiro, Instituto de Biofísica Carlos Chagas Filho. III. Título

iii
Este trabalho foi realizado no Laboratório de Biologia da Superfície Celular do Instituto de
Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, sob orientação do Prof.
Dr. Fernando Costa e Silva Filho, com o apoio das seguintes entidades:
• Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
• MCT - Programa Núcleos de Excelência (PRONEX)
• Fundação Universitária José Bonifácio (FUJB / UFRJ)
• Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ).
• MCT- Nanotecnologia e Materiais Avançados
• Programa “Bolsa nota 10” (FAPERJ)

iv
Esta Dissertação estará sendo submetida à publicação segundo o anexo à folha 91

v
AGRADECIMENTOS Aos meus pais, Ricardo e Célia e ao meu irmão Diogo por acreditarem cegamente em tudo o que faço e por todo o carinho que funcionaram como impulsores para mim. Aos meus padrinhos e avós por todo o apoio e credibilidade incondicionais, devo a vocês uma parte. Ao meu orientador Fernando, que foi meu orientador no real sentido da palavra. Obrigada por me ajudar a aprender a pensar cientificamente. Obrigada também pela credibilidade que me permitiu pensar por conta própria e desenvolver meu senso crítico. Sem dúvida o maior laboratório fica dentro de nossas mentes e você me ajudou a desenvolver um pouco mais o meu. A Juliany por toda a grande disponibilidade e atenção com o meu trabalho, devo a você tudo o que consegui com as pequeninas (Leishmania spp). Ao Bruno, pela amizade, pela orientação cientifica e não cientifica, pelas ajudas práticas do dia-a–dia e por todos os “calma Débora” que podiam até não me acalmar realmente, mas me davam a certeza de não estar sozinha. Ao Gustavo, pela ajuda sempre muito sincera, e pela companhia que se tornou amizade. Foi muito legal perceber que mesmo trabalhando em áreas aparentemente tão diferentes podíamos ajudar um ao outro, e você foi, sem dúvida, de grande ajuda para mim. Como mesmo tão quieto foi capaz de fazer tanta falta dentro do laboratório? A Jéssica, Karina Tatiana e Liliam que vieram aumentar o contingente feminino do laboratório. Obrigada pelo companheirismo e amizade, a convivência com vocês foi sem dúvida animadora. A amiga Mariane, que mesmo estando longe continua me ajudando sem nem saber. Obrigada pela versão colorida das T. foetus e diversas outras ajudas computacionais que me pouparam bastante tempo. Ao Renato, por criar um ambiente de trabalho muito mais organizado do que conseguiríamos sozinhos. A sua presença dentro do laboratório foi sem dúvida de grande ajuda e me poupou bastante tempo. Obrigada! A todos do Laboratório de Microscopia Eletrônica e Ultraestrutura Celular por toda colaboração em especial as amigas Juliany, Joana, Emile e Sarah por toda a paciência e ajuda. A professora Narcisa, por todo o material emprestado, desde Potter até ECL. A colaboração com vocês foi mais do que essencial, muito obrigada! A professora Regina, por toda a colaboração e diversas alíquotas de LMN-1 emprestadas na mais pura confiança. Sua ajuda me salvou alguns importantes meses de experimento, muito obrigada!

vi
Ao Leandro por toda a ajuda amiga dentro do biotério do Laboratório de Microscopia. Sem falar nas inúmeras caronas que me permitiram, de forma segura, chegar cedo e sair tarde do Fundão. As minhas amigas não biólogas Cíntia e Priscilla por toda a amizade, incentivo e ajuda incondicional durante todos esses muitos anos de convivência e não convivência. As minhas amigas biólogas Patrícia e Fabiana por todo o apoio científico e não científico durante todo o período do mestrado. A amizade de vocês me dá energia para seguir sempre adiante. A Joana, minha amiga e companheira de mestrado que desde o período de seleção vem se mostrando ser de grande ajuda. Obrigada por todos os pequenos e grandes favores que você fez por mim durante esses últimos dois anos. A Alessandra, por toda a amizade sincera e disponibilidade mesmo nos seus momentos mais difíceis. A sua amizade é algo que confio e desejo ter sempre comigo. Ao meu namorado Bruno por toda a paciência comigo nestes últimos meses onde estive bastante ocupada. Obrigada por toda companhia, carinho e atenção. Ao meu amigo Diogo que permitiu o meu acesso aos periódicos da Capes do computador da minha casa. Muita obrigada, não só por isso, mas principalmente por toda a amizade. Obrigada vizinho! Ao meu amigo Pablo, atual residente de Houston, por todas as conversas científicas e troca de
idéias. Valorizo muito as nossas conversas intermináveis. Uma pena que agora sejam conversas
com taxa de DDI. Saudades!
Aos meus amigos Arthur e Christiano também por toda a amizade e ajuda.

vii
“Tristes tempos os nossos, em que é mais fácil desintegrar um átomo do que um
preconceito."(Albert Einstein)
“ Não é preciso consenso
nem arte,
nem beleza ou idade:
a vida é sempre dentro
e agora.
(A vida é minha
para ser ousada).
A vida pode florescer
numa existência inteira.
Mas tem de ser buscada, tem de ser
conquistada.”
(Lya Luft / Perdas & Ganhos)

viii
Resumo
A interação de patógenos com a matriz extracelular de seus hospedeiros é uma etapa crítica no processo de infecção, e ainda pouco descrita para a maior parte dos parasitas. Membrana basal é um tipo especializado de matriz extracelular encontrada na região baso-lateral do epitélio, composta principalmente por polímeros de LMN-1 associadas a redes de Colágeno IV, via Entactina. A membrana basal constitui o primeiro contato de muitos parasitas com a matriz extracelular de seus hospedeiros. Tritrichomonas foetus é o protozoário parasita causador da tricomonose bovina, uma doença sexualmente transmissível de grande relevância veterinária. T. foetus é um parasita extracelular e, portanto, a fixação no ambiente extracelular é de extrema importância para a sua sobrevivência. Leishmania (L.) amazonensis é o protozoário parasita causador da leishmaniose cutânea, doença relacionada com a formação de úlceras na região de inóculo do parasita. No hospedeiro humano, L. amazonensis é considerada um parasita intracelular capaz de induzir a sua fagocitose por macrófagos e então sobreviver e se multiplicar dentro de fagolisossomos. Apesar de ser um parasita intracelular, o contato com a matriz extracelular também ocorre durante o processo de infecção por L. amazonensis. Tanto as formas promastigotas metacíclicas (infectivas) quanto as formas amastigotas (reprodutivas) deste parasita entram em contato com o ambiente extracelular de seus hospedeiros. O objetivo deste trabalho foi analisar de forma comparativa o mecanismo de interação destes protozoários parasitas, extracelular (T. foetus) e intracelular (L. amazonensis), com LMN-1. Para isto, ensaios de interação parasito-matriz imobilizada foram realizados, demonstrando que tanto T. foetus quanto as formas promastigotas e amastigotas de L. amazonensis são capazes de se associar a LMN-1 imobilizada em lamínulas de vidro. Demonstramos também que essa associação à LMN-1 causa drásticas mudanças morfológicas em T. foetus que passa a exibir predominantemente uma morfologia amebóide, espraiada. Também através de ensaios de associação demonstramos a participação de cátions divalentes no processo de reconhecimento de LMN-1 por ambos patógenos estudados. Através de ensaios de “Dot Blot” demonstramos a presença de proteínas ligantes de LMN-1 em T. foetus e L. amazonensis. Identificamos então a presença de cinco proteínas reativas a LMN-1 em T. foetus, e quatro em formas promastigotas de L. amazonensis. Verificamos que o reconhecimento de LMN-1 se faz através dos sítios de ligação A13, C16, AG73 e A208 contidos nesta glicoproteína. Por último, analisamos o papel funcional da interação destes patógenos com LMN-1. T. foetus em contato com LMN-1 apresenta exacerbação de citopatogenicidade. Por outro lado, LMN-1 parece funcionar como ponte entre L. amazonensis e sua célula hospedeira uma vez que o número de promastigotas associados a macrófagos é significativamente aumentado na presença de LMN-1.

ix
Abstract
The interaction between pathogens and the host extracellular matrix is an important task
for the infection success. Nerveless, little is still known on such subject. The basal membrane is an specialized form of extracellular matrix which is mostly composed of Laminin-1 and type IV Collagen. The extracellular parasitic protozoan Tritrichomonas foetus is the causative agent of bovine trichomonosis, an urogenital disease of veterinary relevance. Since it is an extracellular parasite its recognition of the host milieu is crucial to the parasite surveillance into host. The intracellular parasitic protozoan Leishmania (L.) amazonensis is the causative agent of cutaneus leishmaniosis, disease characterized by the formation of epithelial ulcers in the site of the vector bite. L. amazonensis is a intracellular parasite adaptated to survive and multiply inside of the acidic vacuoles of macrophages. Promastigotes and amastigotes forms of L. amazonensis have to pass trough the extracellular matrix of the host during the infection course. This work is concerning analyses of the interaction between T. foetus or L. amazonensis and the major glycoprotein of the basal membrane laminin-1 (LMN-1). Attachment assays were carried out in the presence of immobilized LMN-1. Either T. foetus or each one of the both forms of L. amazonensis were capable to recognize LMN-1. Following parasite-LMN-1 interaction we did observed a drastic morphological transformation among T. foetus which became mostly ameboid. A requirement of divalent cations to the interaction of both protozoa parasites to the immobilized LMN-1 was observed. The involvement of laminin binding protein (LBP) on the surface of T. foetus and L. amazonensis was also strongly suggested by dot blot assay. T. foetus and L. amazonensis promasigotes and amasitogotes seems to recognized LMN-1 through the A208, A13, C16 and AG73 LMN-1 related peptides. Five laminin binding molecules seem to exist in T. foetus. On the other hand, promastigotes of L. amazonensis seem to possess four laminin binding molecules.Related to the role played by the binding of each one of the parasites to immobilized LMN-1 we demonstrated that such interactive process enhanced T. foetus citophatogenicity. In the case of L. amazonensis, LMN-1 seems to work as a molecular bridge between promastigotes and macrophages.

x
Lista de Abreviaturas
BSA albumina sérica bovina (do ingles “bovine serum albumin”)
FN fibronectina
GAGs glicosaminoglicanas
Gp63 glicoproteína de 63kDa
GPI glicosil fosfatidil inositol
HeLa linhagem celular de carcinoma uterino humano
HEPES ácido N-2-hidroxietilpiperazina-N’-2-etanosulfônico
LBM “laminin binding molecule” – molécula ligante a laminina
LBP “laminin binding protein” – proteína ligante a laminina
LG laminin globular domain – domínio globular da laminina
LMN –1 isoforma 1 da Laminina
LPG lipofosfoglicana
MB membrana basal
MDBK Madin-Darby Bovine Kidney
MDCK Madin-Darby Canine Kidney
MEC matriz extracelular
MEV microscopia eletrônica de varredura
MP membrana plasmática
PBS solução salina tamponada com fosfato - “Phosphate buffer solution”
PPG proteofosfoglicana
PTGs proteoglicanas
SDS dodecil sulfato de sódio – “sodium dodecil sulphate”
TYM “trypticase - yeast extract - maltose”
VP vacúolo parasitóforo

xi
Sumário Introdução 1) A Matriz Extracelular _______________________________________________ 12) As Lamininas ____________________________________________________ 3 2.1) Isoformas ____________________________________________________ 5 2.2) Isoforma 1 ___________________________________________________ 6 2.3) Seqüências de reconhecimento ___________________________________ 6 2.4) Os papeis funcionais ___________________________________________ 9 2.5) O reconhecimento celular _______________________________________ 93) Interação de patógenos com a MEC __________________________________ 114) Tritrichomonas foetus 4.1) Aspectos clínicos e patológicos __________________________________ 14 4.2) Aspectos gerais ______________________________________________ 16 4.3) A Interação parasita-hospedeiro _________________________________ 18 4.3.1) Reconhecimento e citoadesão ________________________________ 4.3.2) Citotoxicidade _____________________________________________ 4.3.3) Interação com o sistema imune do hospedeiro ____________________ 4.3.4) Interação com a MEC________________________________________
19202223
5) Leishmania (L.) amazonensis 5.1) As leishmanioses _____________________________________________ 25 5.2) Aspectos gerais ______________________________________________ 28 5.3) A Interação parasita-hospedeiro __________________________________ 5.3.1) Reconhecimento, ligação e internalização pela célula hospedeira_____ 5.3.2) A interação com o sistema imune do hospedeiro__________________ 5.3.3) A interação com a MEC do hospedeiro _________________________
323536
Racional e Objetivos ________________________________________________ 38 Metodologia________________________________________________________ 39 Resultados ________________________________________________________ 47 Discussão _________________________________________________________ 64 Conclusão _________________________________________________________ 74 Referências Bibliográficas ___________________________________________ 75 Anexo ____________________________________________________________ 91

1
Introdução
1) A Matriz Extracelular.
Grande parte do volume tecidual animal é composto por uma complexa rede de
macromoléculas. Tal rede, de textura gelatinosa, é comumente referida como Matriz Extracelular
(MEC). Esta é composta por proteínas e polissacarídeos os quais, por sua vez, são secretados por
diversos tipos celulares que compõem os tecidos. Os diferentes tecidos não se diferenciam apenas
em relação aos tipos celulares constituintes, mas também por apresentarem drásticas diferenças
em relação à composição química de suas MECs.
Atualmente, sabe-se que tanto a constituição química quanto a estrutura da MEC possui um
papel determinante para diversas funções celulares. As células e a MEC estão em constante
comunicação, de forma que através do reconhecimento do ambiente extracelular por receptores,
modificações funcionais são geradas na célula, que por sua vez, torna-se capaz de remodelar seu
ambiente extracelular [Bissell & Barcellos-Holff, 1987; Schenk & Quaranta, 2003; Vogel et al,
2001]. As células monitoram seu ambiente extracelular via informações processadas a partir de
ligações receptor-ligante e outras de natureza física, originadas na tribologia (topologia e
topografia) do ambiente extracelular através da percepção da textura e forças de tensão. A
percepção do ambiente é, obviamente, essencial à sobrevivência celular.
1.1) Composição
A MEC animal é composta por glicosaminoglicanas (GAGs) e por proteínas fibrosas
sendo fibronectinas (FN), lamininas (LMN), elastina e colágenos as mais comumente
encontradas.
As GAGs geralmente ocorrem covalentemente ligadas a “esqueletos” protéicos, formando
complexos conhecidos por proteoglicanas (PTGs) [Ruoslahti & Engvall 1994]. Agrecana,
sindecana, decorina, perlecana e betaglicana são exemplos de proteoglicanas. As GAGs são
cadeias polissacarídicas não-ramificadas, compostas de unidades dissacarídicas repetidas, sendo
um dos açúcares aminado (N- acetilglucosamina ou N-acetilgalactosamina); na maioria das vezes

2
também sulfatado. Devido à presença de grupos sulfato ou carboxil, as GAGs apresentam-se
eletronegativamente carregadas. Quatro grupos distintos de GAGs foram identificados e
classificados de acordo com (a) seus resíduos de açúcar, (b) tipos de ligação, (c) o número e (d)
localização dos grupamentos sulfato: ácido hialurônico, condroitin sulfato (e dermatan sulfato),
heparan sulfato (e heparina) e queratan sulfato [Alberts et al, 2002].
Proteínas fibrosas são especialmente abundantes na MEC. Muitas dessas proteínas logo
que são secretadas se estruturam sob a forma de redes ou longas fibrilas. A família dos colágenos
forma a mais abundante rede fibrilar encontrada em tecidos animais (25% em mamíferos), sendo
que o colágeno tipo IV apresenta a capacidade de formar redes [Timpl et al, 1981]. Colágeno
tipo IV, assim como laminina, contém em sua seqüência primária informações necessárias para se
auto-arranjar e interagir entre si e com outros componentes da MEC [Aumailley et al, 1989]. Essa
característica não é encontrada em todos os constituintes da MEC: perlecana e
nidogênio/entactina, por exemplo, não são capazes de se auto-arranjarem. Contudo, a interação
entre colágeno e laminina é facilitada por nidogênio/entactina e perlecana [Sasaki et al, 2004].
Entactina ou nidogênio são, na verdade, designações diferentes para uma mesma glicoproteína de
aproximadamente 150KDa [Aumailley et al, 1989].
1.2) A Membrana Basal.
Lâmina ou Membrana Basal (MB) é um tipo especializado de MEC que se caracteriza
tanto por sua alta densidade óptica, quanto pelo fato de estar sempre em estreita associação com
células. A MB é geralmente encontrada na região baso-lateral do epitélio, do endotélio ou ao
redor de axônios periféricos neuronais, células adiposas e musculares. Os principais componentes
da MB são colágeno tipo IV, LMN, nidogênio/entactina e algumas GAG sulfatadas do tipo
heparina. Em menores proporções também podem ser encontrados agrina, osteopontina, fibulinas
e colágenos, tipos XV e XVIII [Kalluri, 2003]. No entanto, existem diferentes isoformas de
LMN, colágeno, PTG e de nidogênio/entactina. Tais isoformas são expressas diferencialmente
nos diversos tecidos. Portanto, cada MB, assim como a MEC em geral, é composta pela interação
de diferentes isoformas moleculares formando complexos com propriedades distintas e, assim
sendo, são capazes de determinar funções celulares diferentes [Kalluri, 2003].

3
2) As Lamininas:
Lamininas representam uma família de moléculas abundantemente encontradas na MB de
diferentes tecidos. Trata-se de uma família de glicoproteínas cruciformes, flexíveis e
multidomínio, de aproximadamente 850KDa. A LMN é heterotrimérica, formada por uma cadeia
polipeptídica pesada (200-400KDa) denominada α e duas leves (150-200KDa) denominadas β e
γ [Burgeson et al 1994]. As diferentes cadeias α, β e γ de LMN mantêm-se unidas por interações
“coiled-coil” entre os domínios I e II de cada cadeia e, ainda, por associações covalentes entre os
domínios I (Figura 1). As ligações “coiled-coil” formadas via pontes dissulfeto entre as três
cadeias resultam numa longa estrutura estável, assimetricamente entrecruzada (Figura 1) [Ekblon
et al, 2003].
Isoformas: β1γ1 = laminina -1 (LMN-111) α1 β2γ1 = laminina-3 (LMN-122) β1γ1 = laminina-2 (LMN-211) α2 β2γ1 = laminina-4 (LMN-221) β1γ3 = laminina-12 (LMN-213) β1γ1 = laminina-6 (LMN-311) α3 β2γ1 = laminina-7 (LMN-321)
β3γ2 = laminina-5 (LMN-332)
β1γ1 = laminina-8 (LMN-411) α4 β2γ1 = laminina-9 (LMN-421)
β2γ3 = laminina-14 (LMN-423) β1γ1 = laminina-10 (LMN-511) α5 β2γ1 = laminina-11 (LMN-521)
β2γ3 = laminina-15 (LMN-523)
Figura1: Estrutura e domínios presentes na isoforma 1 de laminina. Adaptado de
http://profile.myspace.com/index.cfm?fuseaction=user.viewprofile&friendID=196181691
LMN é capaz de se auto-organizar de forma a construir grandes polímeros, o que é de
grande importância para a formação da MEC. Foi demonstrado que cátions divalentes são
fundamentais ao auto-arranjo da LMN, assim como para sua conformação funcional. Cálcio é o
Cadeia α
Cadeia β Cadeia γ
I
II
III III IV
V VI
IVa
IVb
VI
VI V
IV
IIIa
IIIb
V

4
mais eficiente cátion divalente a mediar a polimerização de LMN [Yurchenco & Shan Cheng,
1993] sugerindo, portanto, ser crucial sua presença para a conformação funcional da molécula.
Assim como cálcio, o pH ácido parece atuar como indutor da autopolimerização de LMN. De
fato, a acidificação do meio induz a mudanças conformacionais que preservam a estrutura
terciária da LMN favorecendo sua rápida polimerização in vitro [Freire & Coelho-Sampaio
2000].
2.1) As isoformas
Atualmente são conhecidas 15 isoformas de laminina (LMN-1 a LMN-15) as quais são
determinadas por diferentes combinações entre as 5 isoformas de cadeia α, as 3 de cadeia β e
outras 3 de cadeia γ conhecidas até o momento (Figura 1) [Yurchenco et al 2004]. Cada isoforma
de LMN apresenta característica, função e uma distribuição tecidual particular. Por isso é
prudente especificar onde e quando (dentro do desenvolvimento do organismo) cada isoforma
pode ser encontrada.
Laminina-2 (α2β1γ1) pode ser encontrada em tecidos musculares esqueléticos e cardíacos, e
no sistema nervoso periférico. Alguns trabalhos sugerem importantes papéis biológicos para
LMN-2 no tecido muscular, já que mutações no gene para a cadeia α2 são a principal causa de
distrofia muscular congênita em humanos e camundongos [Jones et al, 2001]. Receptores do tipo
integrina (α6β1 e α3β1) reconhecem LMN-2.
Laminina-5 (α3β3γ2), também conhecida como epiligrina, calinina ou niceína, apresenta suas
três cadeias truncadas, o que parece estar relacionado a sua incapacidade de se auto - polimerizar.
LMN-5 é um importante componente da adesão epitelial à MB, devido a sua capacidade de ser
reconhecida pelas integrinas α6β4 e α3β1. [Niessen et al, 1994; Rousselle & Aumailley, 1994;
Rousselle et al, 1995]. Uma doença genética rara conhecida como epidermólise bolhosa
congênita apresenta como principal sintoma a formação de bolhas na pele e mucosas, devido a
ineficiência de ancoragem por parte das células epiteliais à LMN presente na membrana basal.
Esta doença está relacionada a mutações nos genes responsáveis pela expressão das cadeias que
formam LMN-5, demonstrando a importância desta isoforma para a adesão epitelial [Muhle et al,
2005]. LMN-5 é a única isoforma atualmente conhecida a apresentar a cadeia γ2. Esta cadeia foi

5
relatada como podendo ser sintetizada e secretada sem a presença das outras cadeias formadoras
da LMN-5 [Koshikawa et al, 1999].
LMN-7, assim como outras LMNs que contêm a cadeia α3, são encontradas na MB de
junções dermo-epiteliais. Laminina-8 e 10 são os componentes majoritários da MB vascular e
foram detectadas na MEC da medula óssea [Gu et al, 2003]. Funcionalmente, estas isoformas
foram descritas como substratos adesivos para células progenitoras hematopoéticas, através do
reconhecimento celular feito por integrinas β1 presente nestas células [Siler et al, 2000]. Novas
isoformas como as LMN-12 (α2β1γ3), LMN-14 (α4β2γ3) e LMN-15 (α5β2γ3) foram descritas
sem, no entanto, se conhecer seus papéis fisiológicos.
2.2) A Isoforma 1
A primeira LMN descrita e por isso conhecida como isoforma 1 (LMN-1), foi purificada a
partir da MB de tumores Engelbreth-Holm-Swarm (EHS). Composta pela ligação entre as cadeias
α1, β1 e γ1, trata-se da isoforma mais bem estudada e bem descrita [Timpl & Brown, 1994].
LMN-1 contêm vários domínios de ligação, muitos deles contidos nas cadeias β1 e γ1, os quais
também se encontram em outras 11 isoformas (Figura 1). No entanto, a região carboxi-terminal
da cadeia α1 contém 5 domínios globulares que a distingue das demais isoformas. Essa região da
cadeia α1 parece ter papel fundamental na adesão celular [Rudenko et al, 2001]. Os domínios
globulares carboxi terminal LG da LMN-1 são também conhecidos por LNS devido a sua
similaridade com domínios encontrados em neurexinas e na globulina do hormônio sexual
(SHBG). No entanto, acredita-se que este domínio seja muito comum entre proteínas da MEC, já
que análises por cristalografia sugerem a existência de domínios LG em pentraxinas, algumas
lectinas e na cadeia α2 presente em outras isoformas de LMN [Rudenko et al, 2001].
2.3) As Seqüências de Reconhecimento
Para facilitar a referência por domínios (estruturas dentro de uma cadeia protéica com
conformação independente) dos componentes da MEC [Timpl & Brown 1994] foi proposta uma
nomenclatura de enumeração dos domínios de LMN, como demonstrado na figura 1, e a

6
utilização das letras A, B, C e LG quando se referem aos domínios da cadeia α, β, γ e globular
carboxi terminal, respectivamente.
Fragmentos resultantes de degradação proteolítica, proteínas recombinantes e peptídeos
sintéticos têm sido utilizados para identificar e caracterizar funcionalmente os mais diversos
domínios contidos na molécula de LMN-1. Desta forma, muitos sítios de ligação presentes na
cadeia α1 tais como A-10, A-13, A-55, A-64, A-167, A-203, A-208, entre outros, estão sendo
alvo de estudos com os mais variados tipos celulares [Nomizu et al, 1998]. A seqüência IKVAV
presente na posição A-208 (Figura 2) da região carboxi – terminal está relacionada a funções
como, por exemplo, de promoção de adesão celular, indução de expansão neuronal, indução de
atividade de colagenase IV, angiogênese, multiplicação celular e tumoral [Tashiro et al., 1989].
Devido a sua atividade promotora de adesão celular o peptídeo IKVAV é bastante estudado e
utilizado como componente adesivo para biomateriais. Presente no domínio globular VI na região
amino terminal da cadeia α-1 (Figura 2), o peptídeo A-13 (RQVFQVAYIIIKA) demonstrou
promover adesão celular mediada por receptores do tipo integrinas (α5β1e αvβ3) e parece estar
relacionado a atividades biológicas como indução de migração, angiogênese e metástase [Nomizu
et al., 2001]. Este domínio VI, também conhecido como amino terminal da cadeia α (LN), está
presente também nas cadeias α2, α3 e α5, o que sugere grande importância fisiológica deste sítio
de reconhecimento [Nomizu et al., 2001]. Presente em outras proteínas da MEC, o tripeptídeo
RGD é um conhecido sítio de ligação para integrinas, como as α3β1 e α6β1, e que foi descoberto
presente também na cadeia α-1 de LMN (Figura 2). No entanto, nessa isoforma de LMN esse tal
sítio se encontra oculto pelo domínio globular IV encontrado ao seu lado [Schulze et al., 1996].
Todavia, a presença de 2 tripeptídeos RGD no domínio IV, descobertos na cadeia α5, são capazes
de interagir com a integrina αvβ3 [Sasaki & Timpl 2001].
Os fragmentos E3 e E8 (Figura 2) são assim chamados por serem fragmentos de digestão por
elastases. Ambos são sítios de ligação marcantes tanto para receptores celulares quanto para
componentes de MEC. Posteriormente descrito como domínio LG, demonstrou-se que um dos
seus peptídeos sintéticos (α1LG4) é capaz de alterar a morfogênese epitelial em alguns sistemas,
e provavelmente está envolvido na ligação a algumas proteoglicanas [Engvall & Wewer, 1996].
Diversos peptídeos presentes no domínio LG têm sido descritos como sítios de ligação
importantes para diversas funções biológicas. Estudos realizados com o peptídeo de posição AG-

7
73 (RKRLQVQLSIRT) mostraram que este pode ser fortemente reconhecido por grande espectro
de receptores [Nomizu et al., 1995]. Este peptídeo está relacionado à indução da diferenciação de
células salivares, estimulação de expansão neuronal e secreção de metaloproteinases. AG-73
parece não ser reconhecido por receptores do tipo integrina mas, por outro lado, já foi descrito
como importante sítio de ligação para receptores contendo heparinas sulfatadas, como a
sindecana-1. Regiões homólogas a LG4-5 foram encontradas nas cadeias α2, α3, α4 e α5,
sugerindo tratar-se de uma região fortemente conservada [Engvall & Wewer, 1996].
Figura 2: Principais sítios de ligação presentes na isoforma 1 de laminina. Adaptado de Ekblom et al., 2003.
Trabalhos similares realizados com as cadeias β e γ comprovaram a existência nessas cadeias
de outros importantes sítios de ligação celular. O peptídeo YIGSR presente na cadeia β promove
adesão e migração celulares, e inibe angiogênese e metástases [Nomizu et al., 2000]. Na cadeia
γ1 os principais sítios de ligação encontrados foram C-16, C-28, C-64, e C-68, todos localizados
no domínio globular amino-terminal (LN) (Figura 2). Este domínio LN, assim como toda a
A-208
C-16
A-13
AG-73
RGD YIGSR

8
cadeia γ é bastante conservada dentro da família das LMNs [Nomizu et al., 1997], estando
presente na maioria das isoformas de LMN. Isso pode ser explicado pela participação desta
cadeia na interação com nidogênio/entactina, a qual é muito importante para a estruturação da
MB. A interação entre LMN e nidogênio/entactina se dá através de repetições de domínios ricos
em cisteínas conhecidos como EGF (fatores de crecimento epitelial), localizados próximos a
região amino–terminal da molécula de LMN. Entre os peptídeos da cadeia γ melhor estudados, C-
16 (KAFDITYVRLKF) é aquele que apresenta maior atividade de ligação a células, e que se
revelou inibido por EDTA (quelante de calcio), sugerindo participação de cálcio para sua
conformação funcional. No entanto, C-16 parece não ser reconhecido por receptores do tipo
integrinas [Nomizu et al., 1997].
2.4) Papéis funcionais.
Lamininas possuem grande importância fisiológica uma vez que estão diretamente envolvidas
na adesão [Terranova et al 1980], proliferação [Kikkawa et al, 2004] e migração [Gu et al, 2003]
celulares, assim como na expansão de neuritos [Richard et al., 1996], em metástases [Patarroyo et
al, 2002], na regulação do desenvolvimento embrionário [Yurchenco et al, 2004] e na
diferenciação celular [Adams & Watt 1993], entre outras. LMN-1 desempenha papel primordial
no arranjo estrutural da MB assim como na adesão de células epiteliais [Sasaki et al, 2004].
Durante as primeiras etapas do desenvolvimento embrionário, LMN-1 pode ser detectada junto
ao epitélio em desenvolvimento. Em tecidos adultos, a expressão de LMN-1 passa a ser restrita à
MB de alguns tecidos epiteliais [Yurchenco et al, 2004]. Além disso, LMN-1 atua como
importante proteína adesiva para vários tipos celulares, o que a tornou um agente de
sobrevivência para o cultivo in vitro de muitas células. LMN-1 é também capaz de induzir
diferenciação celular e de induzir a produção de proteínas do leite por células da glândula
mamária [Boudreau et al, 1995].
2.5) Reconhecimento Celular.
Como anteriormente descrito, devido ao seu amplo espectro de reconhecimento LMN é capaz
de induzir várias funções. São diversos os tipos celulares capazes de reconhecer e de se ligar a

9
LMN, assim como são várias as possibilidades de ocorrência de complexos receptores para LMN
em um mesmo tipo celular. Por outro lado, um mesmo complexo receptor pode apresentar
diferentes afinidades por diferentes sítios de reconhecimento presentes em uma mesma isoforma
de LMN. Tal afinidade diferencial pode levar à indução de funções celulares variadas [Mercúrio,
1995].
Os receptores para LMN presente na superfície de eucariotos melhor estudados são as
integrinas e a α-distroglicana. Integrinas fazem parte de uma grande família de proteínas
transmembrana heterodiméricas (subunidades α e β) que atuam principalmente como receptores
para a seqüência RGD presente em vários componentes de MEC, fazendo a conexão com o
citoesqueleto e moléculas de sinalização intracelular. Inúmeras integrinas, principalmente da
família β1, foram relatadas como capazes de interagir com diversas isoformas de LMN: α1β1,
α2β1, α3β1, α6β1, α7β1, α5β1, α9β1, αvβ3, αvβ5 e α6β4 [Plow et al, 2000 e Kikkawa et al.,
2004]. LMN-1 é reconhecida pelas integrinas α1β1, α2β1, α6β1, αvβ3, α5β1, α7β1 e α6β4,
mas o papel da ligação com α1β1 e α2β1 na adesão parece ser mínimo e a real função desta
ligação ainda não se encontra bem descrita. Células podem utilizar mais de uma integrina para
interagir com uma mesma isoforma de LMN, sugerindo que uma mesma isoforma de LMN pode
induzir a diferentes funções celulares. Por outro lado, uma mesma integrina associada aos
mesmos sítios de ligação de LMN-1 pode sinalizar diferentes funções. Isto se deve ao fato de que
o sítio de ligação das integrinas é dependente de conformação e pode ter sua afinidade alterada
para um ou outro ligante [Mercúrio, 1995]. A integrina α6β1, por exemplo, possui duas variações
(A e B) que modulam diferentemente a força de adesão, morfologia e a migração celulares [Shaw
& Mercúrio, 1994]. Diferentes afinidades para LMN-1 também foram relatas entre diferentes
variações da integrina α7β1 [Nishiuchi et al., 2006]
Além das integrinas, outras moléculas de superfície celular também fazem ligações com
LMN: α-distroglicana [Gee et al, 1993], receptores para LMN de 32kDa e 67kDa (receptor
elastina-laminina), galectina-1 [Van der Brûle et al, 1995], lectina de 14kDa [Cooper et al, 1991]
e β1,4-galactosiltransferase (60kDa) [Shur, 1993]. Sozinhas ou em combinação (co-receptor)
com integrinas, essas proteínas ligantes de LMN (LBPs) contribuem para a formação ou a
regulação do complexo de interação de LMN com a superfície celular.

10
LMN apresenta uma alta composição de carboidratos, entre 13 a 15% da sua composição total
[Chung et al., 1979]. LMN-1, por exemplo, apresenta 40 N-oligossacarídeos, de acordo com os
12 a 14 potenciais sítios aceptores (Asn-Xaa-Thr) presentes nas cadeias α, β e γ. A estrutura
básica compreende diferentes tipos de complexos dentre os quais unidades oligolactosídicas que
são diversificadas por terminações de resíduos α-galactose e ácido siálico [Fjiwara et al., 1988;
Arumugham et al., 1986]. Os sítios N-oligossacarídeos não se encontram randomicamente
distribuídos pela molécula de LMN, o que sugere distintas funções e a presença de controle
topológico desses domínios glicídicos [Fjiwara et al., 1988]. Além de aumentar a estabilidade
desta proteína contra proteases, a porção oligossacarídica de LMN é também utilizada como sítio
de ligação para diversos receptores do tipo lectina. Galectina –1, por exemplo, é uma proteína do
tipo lectina específica para resíduos β-galactosídeos, capazes de se associar com LMN [Van der
Brûle et al, 1995; Moiseeva et al., 2003]. L-14, lectina ligante a resíduos de lactose, está presente
em tecidos musculares e é capaz de reconhecer LMN e inibir a interação de LMN com a integrina
α7β1 [Gu et al, 1994]. Outras moléculas receptoras de LMN, do tipo lectina, foram encontradas
em vários tipos celulares [Elola et al., 2005; Sparrow et al., 1987; Woo et al., 1990].
LMNs também podem se comportar como lectinas, reconhecendo porções oligosacarídicas de
outras moléculas, como descrito para heparinas e α-distroglicana. Distroglicana é o segundo mais
comum receptor de LMN, composto por uma subunidade α extracelular e outra β
transmembrana, a qual é o componente central do complexo de glicoproteínas associadas a
distrofina. A ligação de distroglicana com LMN é dependente de sítios encontrados no domínio
α1LG4 (Figura 1). AG-73 não é um sítio de ligação para distroglicanas, mas se encontra bem
próximo a ele. No entanto, AG73 parece estar relacionado a ligação de LMN a algumas
proteoglicanas de superfície [Ekblom et al. 2003].
A maior parte das moléculas da MEC é mais ativa para o reconhecimento in vitro, quando
estão imobilizadas sobre um substrato. Assim, grande parte dos dados publicados sobre as
interações células-MEC foi obtida utilizando-se matrizes em duas dimensões (2D) onde
componentes de MEC são associados a substratos inertes (plástico ou vidro, principalmente). No
entanto, o uso de matrizes tri-dimensionais (3D) é muito importante para análises mais próximas
à realidade in vivo sobre os efeitos da interação célula-MEC [Grinnell, 2003].

11
3) Interação de patógenos com a MEC.
Patógenos são capazes de monitorar e responder ao ambiente extracelular resultando no
estabelecimento de processos infecciosos [Petrópolis et al., 2007]. O reconhecimento do
ambiente extracelular do hospedeiro é uma etapa importante no processo infeccioso, tanto por
patógenos extracelulares quanto por aqueles ditos intracelulares [Nde et al., 2006]. Moléculas
ligantes de componentes na MEC tem sido descritas em diversos organismos patogênicos
(bactérias, fungos e protozoários). Um grupo de moléculas receptoras a componentes de MEC,
bastante conhecida em patógenos são as MSCRAMMs (do inglês, microbial surface components
recognizing adhesive matrix molecules). Estas são moléculas ditas adesivas encontradas
originalmente na superfície de bactérias. Entretanto, até o momento ainda não se identificou
nenhum processo de sinalização decorrente da ligação de MSCRAMMs. Assim, acredita-se
serem as MSCRAMMS receptinas ao invés de receptores propriamente ditos. Receptinas, por sua
vez, são proteínas microbianas com ligação específica para certas moléculas da MEC mas sem
desencadear efeitos intracelulares secundários diretos [Kronvall & Jonsson 1999].
Moléculas tipo-integrina (“integrin-like molecules”) foram identificadas na superfície de uma
série de patógenos, como por exemplo, o receptor para fibronectina identificado na superfície de
Trypanosoma cruzi [Ouaissi et al, 1986] e o receptor similar a integrina β1 identificado na
superfície de Entamoeba histolytica e que está relacionado a ligação do parasita a fibronectina e
colágeno tipo I [Sengupta et al, 2001]. Um receptor tipo integrina de 120kDa foi também
identificado na superfície de Mycobacterium avium [Rao et al, 1992]. Outras moléculas
receptoras para componentes de MEC, cátions-dependentes, foram descritas em parasitos [Wang
et al., 1994]. Receptores do tipo lectina reconhecendo oligosacarídeos de glicoproteínas e
proteoglicanas também foram encontrados em patógenos [Hoire et al., 2005; Saarela et al.,
1996]. A maioria dos receptores para LMN descritos em patógenos até o momento parecem ser
glicoproteínas, onde o sítio de reconhecimento pode se encontrar tanto na porção protéica [Hoire
et al., 2005] como na porção glicídica da molécula [Ferreira et al., 2006]. Portanto, várias são as
estruturas utilizadas por organismos patogênicos para reconhecer a MEC de seus hospedeiros. Tal
como relatado em células de mamíferos, parasitos também podem possuir receptores para mais
de um componente da MEC, e ainda, mais de um receptor para uma mesma molécula da matriz
de forma a permitir não só ancoragem, mas também, o monitoramento do ambiente extracelular.

12
Moléculas da MEC podem funcionar para patógenos como guias durante o processo de
invasão tissular, levando-os até seus sítios preferenciais ou nichos de infecção [Pradel et al.,
2002]. A presença de uma determinada combinação de isoformas de LMN e colágenos, por
exemplo, pode localizar o sítio de infecção de um determinado patógeno sendo, portanto, este
reconhecimento de extrema importância para a sobrevivência deste patógeno. A interação
parasita-MEC parece proporcionar uma permanente troca de sinais tanto de “fora para dentro”
(MEC-parasita) como de “dentro para fora” (parasita-MEC). Tal “comunicação” bidirecional
capacita o parasita a “decidir” onde infectar [Petrópolis et al., 2007]. Através da ação de
receptores verdadeiros, alguns protozoários parasitas são capazes de modular suas funções em
decorrência do reconhecimento inicial de componentes da MEC. Entamoeba histolytica, por
exemplo, possui receptor “integrina-like” capaz de ativar cascata de sinalização a qual culmina na
indução de polimerização de actina e formação de fibras de estresse importantes para o processo
de adesão [Hernandez-Ramirez et al., 2000; Flores-Robles et al., 2003; Talamás-Rohana & Rios,
2000]. Além da promoção da adesão, em Entamoeba histolytica, a ligação de seus receptores com
proteínas da MEC modula o seu potencial invasivo e patogênico [Franco et al., 2002; Li et al.,
1995]. Muitos parasitas utilizam MMPs (metaloproteases de matriz) para a promoção de invasão
tecidual [Lauwaet et al, 2000]. MMPs são proteínas transmembranas ou secretadas e que
possuem um sítio catalítico dependente de zinco, sendo responsáveis pela proteólise da MEC.
Pseudomonas aeruginosa, por exemplo, possui uma MMP capaz de degradar LMN facilitando
sua disseminação pelo hospedeiro [Heck et al, 1986]. A modulação do potencial invasivo por
ligação à MEC pode ser observada também em modelos de patógenos intracelulares como
Toxoplasma gondii, capaz de modificar adesividade de macrófagos infectados a MEC [Da Gama
et al., 2004]. A presença de determinadas proteínas da MEC também parece estar fortemente
relacionada com a capacidade de alguns parasitos de passarem através da barreira hematocefálica
[Silva et al., 1999; Masocha et al., 2004], etapa importante na formação de quadros clínicos mais
graves. A associação de receptores da superfície de patógenos com componentes da MEC pode
funcionar como sinalizador para a promoção de adesão, diferenciação, invasão tecidual e celular,
entre outras importantes atividades do processo infeccioso [Petrópolis et al., 2007; Li et al., 1995;
Silva-Filho et al., 1988]. É importante enfatizar que grande parte dos dados sobre interação
parasito-MEC foi obtida utilizando-se matrizes em 2D. No entanto, o uso de matrizes 3D é muito

13
importante para análises mais completas sobre os reais efeitos da interação parasito-MEC no
processo infeccioso [Grinnell, 2003; Behnsen et al., 2007; Hansen et al., 2006].
Atualmente é vasto o conhecimento de microorganismos capazes de se associar a MEC.
Diversas bactérias como Staphylococcus aureus, Streptococcus pyogenes, Escherichia coli entre
outras, tiveram suas receptinas para LMN descritas [Kronvall & Jonsson, 1999]. Da mesma
forma, é conhecido o reconhecimento de LMN por parte de alguns protozoários parasitas como
Plasmodium berghei [Arrighi & Hurd, 2002], Entamoeba histolytica [Li et al, 1995],
Trichomonas vaginalis e Tritrichomonas foetus [Silva Filho et al, 1988], Leishmania donovani
[Ghosh et al, 1996] e Acanthamoeba polyphaga [Gordon et al, 1993], entre outros.
É importante mencionar o papel de componentes da MEC na modulação do sistema imune e,
portanto, realçar a determinação do sucesso ou fracasso do processo infeccioso pela existência de
um desequilíbrio entre as modulações que levam a ativação do sistema imune e as modulações
que levam ao seu silenciamento. Mindina é o exemplo mais bem conhecido de molécula da MEC
funcionando como “armadilha” para organismos patogênicos. Esta proteína se liga diretamente a
moléculas antigênicas como LPS, comumente encontrada na superfície de diversos
microorganismos, funcionando como opsonina para macrófagos [He et al., 2004; McDonald &
Nuñez, 2004]. Outros componentes da MEC podem funcionar como promotores de inflamação,
recrutando células inflamatórias e regulando sua ativação e desativação [Morwood & Nicholson,
2006] no entanto, pouco se sabe sobre os mecanismos envolvidos neste processo.
Está cada vez mais evidente a importância da regulação da interação de patógenos com a
MEC hospedeira para o sucesso da infecção. Assim, alguns autores propõem o uso de peptídeos
sintéticos que contenham seqüências de aminoácidos de componentes de MEC como ferramentas
quimioterápicas para uso na clínica médica [Flock, 1999].

14
4) Tritrichomonas foetus
4.1) Aspectos clínicos e patológicos
Tritrichomonas foetus é o protozoário parasita causador da tricomonose bovina, doença
sexualmente transmissível que apresenta distribuição cosmopolita e pode causar grandes
prejuízos em criações destinadas ao corte e a produção de leite. T. foetus habita o trato
reprodutivo de touros e vacas, principalmente a região mucosal. Em touros os parasitas são
encontrados especialmente na cavidade prepucial, apesar de também estarem presentes no pênis.
A infecção raramente se manifesta, podendo persistir por anos sem nenhum sintoma clínico
detectável [Skirrow & BonDurant, 1988]. Mesmo quando se manifesta a infecção é geralmente
branda, revelando-se como uma inflamação no tecido prepucial associada a uma descarga
mucopurulenta [Parsonson et al, 1974]. Geralmente não são observadas lesões significativas e a
infecção não afeta a qualidade do sêmen ou o comportamento sexual do animal. Portanto, em
geral, os touros infectados atuam como carreadores assintomáticos de T. foetus [Anderson et al.,
1994]. Nas fêmeas, o parasita pode ser encontrado colonizando a vagina, o cérvice, o útero e os
ovidutos. O quadro patológico é geralmente mais grave, podendo variar de infecções
imperceptíveis até casos clínicos mais severos, onde o parasita se alastra causando vaginite,
endometrite aguda e infertilidade transiente. O endométrio e o tubo uterino são as regiões mais
afetadas pela infecção por T. foetus; duas regiões determinantes na concepção e gestação. Fêmeas
grávidas infectadas podem apresentar quadros de aborto provocados pela destruição das ligações
placentárias pelo parasita e quadros de endometrite crônica que pode tornar o animal
permanentemente estéril [Parsonson et al, 1976].
O diagnóstico da infecção por T. foetus é baseado na detecção do parasita em secreções
vaginais ou prepuciais, sendo a sensibilidade destes métodos variável: entre 58,7% com amostras
de muco vaginal a 81,6% para amostras de esmegma prepucial. A baixa eficiência e a demora de
confirmação dos resultados por estes métodos têm estimulado a busca de novas técnicas para o
diagnóstico da tricomonose bovina. Não existe até o momento uma terapia comprovada e
aprovada contra tricomonose bovina. Derivados de imidazol têm sido amplamente testados no
tratamento de bovinos portadores de tricomonose. Dentre estes, o dimetridazol parece ser a droga
mais eficiente; no entanto, sua utilização não é permitida em muitos países [Corbeil, 1994].

15
Diversos estudos têm buscado o desenvolvimento de vacinas para o controle da doença. Vacinas
produzidas com frações de membrana ou extrato total de T. foetus parecem levar novilhos
imunizados a produzirem resposta imune contra T. foetus, resultando na diminuição do tempo de
eliminação da infecção e da taxa de aborto [Cobo et al., 2002]. No entanto, essa vacina ainda se
encontra em fase experimental e não estando, portanto, disponível comercialmente. Na ausência
de imunoprofilaxia e terapêutica eficientes, o controle da infecção é feito por meio de
gerenciamento do gado, tanto por isolamento de touros infectados quanto por técnicas de
inseminação artificial. Além de dispendiosos, estes métodos são pouco eficientes, fazendo com
que a tricomonose seja responsável por importantes perdas econômicas em atividades
agropecuárias. Em 1999 estimava-se que cada touro infectado gerasse uma perda anual de
4.800,00 dólares, e que a tricomonose bovina resultasse em um prejuízo anual de 6,1 milhões de
dólares nos Estados Unidos [Rae et al., 1999].
Pouco se sabe sobre os aspectos bioquímicos e moleculares envolvidos no
estabelecimento da infecção por T. foetus e, portanto, é comum a correlação com dados obtidos
em estudos que abordam a tricomoníase humana. Esta patologia é causada por Trichomonas
vaginalis, parasita que guarda semelhanças morfológicas, fisiológicas e evolutivas com o agente
causador da doença bovina. Ao contrário da tricomonose bovina, a infecção humana é passível de
tratamento. Desde 1959, derivados de nitroimidazóis vêm sendo utilizados no tratamento de
pacientes portadores de T. vaginalis, sendo metronidazol o composto mais comumente utilizado.
No entanto, já foram relatados casos de pacientes com infecções resistentes ao tratamento [Petrin
et al., 1998].

16
4.2) Aspectos gerais de T. foetus
Tritrichomonas foetus possui a seguinte identidade sistemática (Cavalier-Smith, 1993):
Reino: Protozoa
Sub-Reino: Dictyozoa
Filo: Parabasalia
Classe: Zoomastigophora
Ordem: Trichomonadida
Família: Trichomonadidae
Gênero: Tritrichomonas
Figura 3: Modelo esquemático da ultraestrutura de Tritrichomonas foetus. A: axóstilo; C: costa; CI: corpo
infracinetosomal; FA: flagelos anteriores; FP: filamentos parabasais; FR: flagelo recorrente; G: complexo de Golgi;
Gl: Grânulos de glicogênio; H: hidrogenossomos; L: lisossomo; MO: membrana ondulante; N: núcleo; P: pente; Pe:
pelta. Adaptado de Benchimol & Engelke (2003).
O Filo Parabasalia é caracterizado por organismos multiflagelados que não apresentam
mitocôndria, peroxissomas ou glicossomas, mas apresentam hidrogenossomas envoltos por
membrana dupla como organela produtora de energia; presença de complexo de Golgi
desenvolvido e axóstilo. A mitose é fechada, com fuso exonuclear, e os ribossomos são 70S,
típicos de procariotos [Cavalier-Smith, 1993]. A família Trichomonadidae compreende
L
N
G
A
P e
F P F P
F A
G l
F R
M O
PC I
C
H

17
protozoários flagelados divididos em duas sub-famílias: Trichomonadinae, à qual pertence os
patógenos humanos Trichomonas vaginalis e Pentatrichomonas homini, e Tritrichomonadinae,
que inclui T. foetus [Kirby, 1951].
T. foetus possui quatro flagelos, sendo três anteriores e um recorrente associado à
superfície celular, formando uma estrutura conhecida como membrana ondulante (figura 3). Os
movimentos dos três flagelos e da membrana ondulante explicam a grande motilidade deste
organismo. T. foetus é uma célula polimórfica podendo variar de piriforme à fusiforme. Todavia,
em condições de estresse, verifica-se a presença de formas arredondadas com flagelos
internalizados. Esta forma é chamada por alguns autores de pseudocisto [Granger et al, 2000].
Em média apresentam um corpo celular alongado, medindo de 15 a 22µm de comprimento e
entre 4,5 e 10µm de largura [Honigberg, 1978].
T. foetus apresenta um núcleo localizado na região anterior do corpo celular, cercado por
um envelope nuclear poroso típico dos eucariontes [Petrin et al., 1998]
O citoesqueleto dos tricomonadídeos é composto por tubulina e fibras de actina, sendo
que diferentes tipos de tubulina são encontrados dentro de uma mesma célula e a actina presente
difere daquela encontrada em mamíferos. É marcante a presença do axóstilo, estrutura
microtubular que corta longitudinalmente a célula, definindo o eixo antero-posterior do parasita
[Honigberg, 1978]. Nos tricomonadídeos, o axóstilo está associado aos hidrogenossomos,
retículo endoplasmático e grânulos de glicogênio [Benchimol, 2000] e sua principal função está
relacionada à divisão celular, uma vez que esta estrutura participa da constrição do núcleo
durante a cariocinese [Ribeiro et al, 2000]. A costa é outra importante estrutura citoesquelética
dos tricomonadídeos. Trata-se de uma estrutura fibrilar estriada, associada ao corpúsculo basal do
flagelo o que sugere participação no batimento flagelar da membrana ondulante [Honigberg,
1978]. Os demais flagelos se originam nos corpúsculos basais que fazem parte do aparelho
parabasal, estrutura que se encontra envolta por uma extensão do axóstilo em forma de lua
crescente denominada pelta (figura 3).
In vitro, o metabolismo energético dos tricomonadídeos é baseado na degradação de
carboidratos. In vivo, no hospedeiro bovino, T. foetus encontra pouca disponibilidade de
carboidratos. Assim, o parasita parece também se utilizar da via de síntese de poliaminas, tendo
arginina como precursor, para produzir energia [Yarlett, 1988]. O metabolismo de açúcares em
T. foetus (que não possui mitocôndrias) é fermentativo, localizado em duas regiões diferentes da

18
célula: no citoplasma, onde ocorre a via glicolítica, e no hidrogenossomo, onde ocorre o
“metabolismo hidrogenossomal” [Lindmark & Muller, 1973]. Hidrogenossomos, portanto, são
organelas de grande importância metabólica para este protozoário.
T. foetus é considerado “aerotolerante” [Coombs & Muller, 1995], ou seja, não necessita de
oxigênio para sua sobrevivência e multiplicação uma vez que não o utiliza como aceptor final de
elétrons em seu metabolismo energético. Apesar de ser capaz de viver na presença de oxigênio,
este protozoário é extremamente sensível a elevadas concentrações de oxigênio [Mack & Muller,
1978]. Crescimento ótimo in vitro de T. foetus é obtido em cultivos em baixas concentrações de
oxigênio e na presença de 5% CO2 [Mack & Muller, 1978].
T. foetus possui um ciclo de vida bastante simples, monogenético, e apenas uma forma
evolutiva conhecida como “trofozoíta”, a qual se reproduz assexuadamente por mitose fechada
[Kofoid & Swezy, 1915; Ribeiro et al., 2002]. O tipo de mitose encontrada em T. foetus é
denominado fechado uma vez que o envelope nuclear não é rompido e os microtúbulos do fuso e
do cinetócoro são extranucleares [Alberts et al., 2002]. Os hospedeiros naturais são os bovinos
Bos taurus e Bos indicus, mas ocasionalmente esse protozoário pode ser encontrado em suínos,
eqüinos e felinos [Skirrow & BonDurant, 1988].
4.2) A interação parasita - hospedeiro.
O sucesso da infecção de T. foetus depende de várias adaptações que levem o parasita a
reconhecer e a colonizar o hospedeiro, ser capaz de competir com outros organismos e sobreviver
ao ataque do sistema imunológico e a variações ambientais. Para T. foetus, o hospedeiro é acima
de tudo sua fonte de nutrientes. Devido à carência de diversas vias metabólicas os
tricomonadídeos em geral dependem diretamente de seus hospedeiros para sobreviver. Por isso é
importante a captação de nutrientes no sítio de infecção por parte do parasita. Os processos de
citoadesão, atividade enzimática e evasão do sistema imune são considerados importantes fatores
de virulência para T. foetus.
Em Trichomonas vaginalis é descrita uma maior quantidade de moléculas envolvidas com a
patogênese. Além disso, existem descrições da interação do parasita com a flora vaginal do
hospedeiro que fornecem importantes informações sobre a ecologia do protozoário [Petrin et al.,
1998]; conhecimento este ainda inexistente em T. foetus.

19
4.2.1) Reconhecimento e citoadesão.
O primeiro contato de T. foetus com seu hospedeiro ocorre na camada mucosa que recobre
todo o epitélio genital. Para a colonização do epitélio subjacente esse protozoário deve primeiro
ser capaz de degradar mucinas para, em seguida, reconhecer a célula epitelial. A adesão de T.
foetus ao muco cervical bovino parece ser feita as custas de lectinas específicas para ácido siálico
[Babál & Russel, 1999]. Em T. foetus foram descritas glicosidases, como a β-N-acetil
galactosaminidase, que possui como substrato grupos N-acetil galactosamina predominantes na
composição de mucinas [Connaris & Greenwell, 1997]. Além disso, foi demonstrado que
tricomonadídeos apresentam um repertório de glicosidases, como galactosidases e glucosidades,
indicando que o parasita pode degradar a mucina e outros complexos glicídicos de maneira que
além de fazer parte do processo patológico da infecção, a mucina pode se tornar fonte de energia
para o parasita [Connaris & Greenwell, 1997].
Foi demonstrado em T. foetus que a citoadesão é o primeiro passo para o processo de
destruição de monocamadas epiteliais [Silva-Filho & de Souza, 1988] e que seu efeito citotóxico
pode ser inibido por pré-tratamento do parasita com periodato (agente oxidante de glicídeos),
indicando a participação de glicoconjugados de superfície na destruição de células hospedeiras
[Singh et al, 1999].
Quando em contato com células epiteliais T. foetus sofre uma grande transformação
morfológica: de piriforme a amebóide, totalmente espraiada. Sabe-se que T. foetus é capaz de
utilizar a projeção posterior do axóstilo no processo de citoadesão, apesar de adesão mediada
pelos flagelos anteriores também ter sido descrita [Silva-Filho & de Souza, 1988]. Além disso,
participação de feixes de actina no processo de adesão celular foi demonstrada. Nota-se também
que a relação parasita-célula hospedeira se dá por interações protéicas já que parasitas pré-
tratados com tripsina aderem significativamente menos a monocamadas de células epiteliais
[Silva-Filho & de Souza, 1988]. A interação de T. vaginalis e células hospedeiras envolve a
participação de associações do tipo receptor ligante e dependente da presença de cátions
divalentes e pH [ Alderete & Garza, 1985; Silva-Filho et al., 1987]
Foram identificadas moléculas de superfície em T. foetus que parecem estar relacionadas
com o processo de citoaderência. A adesina Tf190, um glicoconjugado de 190 kDa com

20
complexa constituição glicídica, formada por fucose, galactosamina, glucosamina, galactose,
glucose e manose foi descrita em T. foetus. Verificou-se que Tf190 apresenta afinidade à lectina
Concavalina A, indicando papel de resíduos terminais de manose no reconhecimento molecular
feito pelo parasita [Shaia et al, 1998]. Outro glicoconjugado (Tf1.17) foi descrito e relacionado
com a adesão de T. foetus à células vaginais bovinas (BVEC) [Corbeil et al, 1989]. Tf1.17 parece
estar relacionada não só a adesão ao epitélio vaginal mas também sua inibição acarreta na
imobilização dos parasitas e na sua lise mediada por complemento [Corbeil, 1994]. Análises
bioquímicas mostraram que Tf1.17 é um glicoconjugado de aproximadamente 60kDa de peso
molecular [Hodgson et al., 1990]. Singh e colaboradores (1999) isolaram e caracterizaram
parcialmente uma lipofosfoglicana da superfície de T. foetus denominada TF-LPG. Trata-se de
uma glicoconjugado bastante representativo na superfície do parasita (2-3. 106 cópias por
organismo) o qual está envolvido na adesão de T. foetus à células epiteliais bovinas. Sabe-se
também que células hospedeiras apresentam na sua superfície moléculas de reconhecimento para
ácido siálico, as quais estão presentes em altas concentrações na superfície de T. foetus [Silva-
Filho et al., 1982]. É importante mencionar que nenhuma molécula isolada de T. foetus até o
momento é única responsável pela adesão do parasita a células hospedeiras, reforçando a idéia de
que este processo de adesão é multifatorial.
Ferro iônico atua como regulador negativo do processo de citoaderência, uma vez que a
aderência de T. foetus à monocamadas de células HeLa é aumentada em condições de baixa
concentração de ferro no meio de cultivo [Melo-Braga et al, 2003].
4.2.2) Citotoxicidade
Diversos autores monstraram que T. foetus pode induzir a danos em células epiteliais in
vitro, porém as bases moleculares deste processo ainda não são bem conhecidas. A atividade
citotóxica de T. foetus foi evidenciada utilizando-se diferentes modelos epiteliais, incluindo
células HeLa, MDCK [Silva-Filho et al., 1982; Burgess et al., 1990] e MDBK [Singh et al.,
1999].
A adesão de T. foetus à monocamadas formadas por células MDCK pode ser o primeiro
passo para o processo citotóxico que se segue [Silva-Filho & de Souza, 1988]. Assim, como
demonstrado em T. vaginalis, a citoaderência pode disparar processos de sinalização intracelular,

21
ativando uma série de outras moléculas fundamentais ao processo de citopatogênese [Alderete et
al., 1995]. Desta forma, o efeito citotóxico deste parasita pode ser considerado dependente de
contato [Singh et al., 1999], apesar do envolvimento de fatores líticos secretados por parasitas
não aderidos à células epiteliais [Silva-Filho & de Souza, 1988; Burgess et al., 1990; Kennett &
Hook, 2002]. Utilizando T. foetus pré-incubado com anticorpos monoclonais contra a adesina
Tf190 Burgess e McDonald (1992), inibiram parcialmente o efeito citotóxico. Neste mesmo
trabalho, observou-se que populações do parasita com altas taxas de adesão apresentaram alto
efeito citotóxico contra células HeLa, sugerindo uma correlação positiva entre citoadesão e
citotoxicidade.
O evento de citotoxicidade aqui comentado parece ser saturável pela densidade de
parasitas, podendo ser inibido pelo pré-tratamento dos mesmos com periodato, indicando o papel
de glicoconjugados acídicos não só na adesão mas, também, na destruição de células hospedeiras
[Bonilha et al., 1995; Singh et al., 1999].
A incubação de T. foetus e células MDCK na presença de inibidores de protease diminui o
dano causado pelo parasita a células MDCK [Silva-Filho & De Souza, 1988], sugerindo
envolvimento de proteases na toxicidade de T. foetus a tais células. Além disso, o efeito
citotóxico causado por sobrenadantes de cultura de T. foetus à células MDCK e HeLa foi também
evidenciado. Serino e cisteíno proteases parecem ser os principais tipos de enzimas envolvidas na
citotoxicidade de T. foetus [Burgess et al., 1990; Melo-Braga et al., 2003]. T. foetus parece
também se utilizar de glicosidases como neuraminidases, para danificar células in vitro [Silva-
Filho & de Souza, 1988].
Embora a literatura aponte para o papel fundamental da secreção de proteases na
citopatogenicidade de T. foetus, Kennett & Hook (2002) responsabilizam uma citotoxina não
enzimática e solúvel de 40KDa como fundamental no processo.
Extratos de T. foetus, parasitas inteiros ou ainda suas secreções podem lisar hemácias
bovinas. É importante mencionar que o efeito hemolítico de tais secreções é mediado por fatores
não proteolítcos, uma vez que inibidores de proteases não são capazes de inibir tal processo
[Burgess et al., 1990]. A hemólise é de grande importância a sobrevivência de T. foetus já que
hemácias representam para o parasita fontes de nutricionais de ferro e lipídeos [Felleisen, 1999].
É extremamente difícil imputar-se a uma única classe de moléculas presente em T. foetus
papel preponderante na sua interação com células in vitro, visto que in vivo o nicho ocupado pelo

22
parasita encontra-se co-habitado com vírus, bactérias e fungos os quais podem, diretamente ou
indiretamente, influenciar no metabolismo do protozoário de forma a modular sua citotoxicidade.
É, portanto, importante termos em mente que a tricomonose bovina apresenta-se sempre como
infecção múltipla [Petrin et al., 1998].
4.2.3) Interação do parasita com o sistema imune do hospedeiro
T. foetus é normalmente eliminado do trato reprodutivo de fêmeas após alguns meses de
infecção, diferentemente da infecção persistente geralmente encontrada em machos [Skirrow &
BonDurant, 1988]. Também se observa certo grau de resistência temporária de bovinos pré-
parasitados à reinfecções [Skirrow & BonDurant, 1988]. No entanto, tanto os mecanismos que
medeiam essa resposta imune protetora como a forma pela qual T. foetus é eliminado, ainda são
pouco conhecidos [Felleisen, 1999].
Cisteíno proteases secretadas e/ou associadas à superfície do parasita revelam-se
fundamentais ao escape do parasita a resposta imune do hospedeiro, uma vez que tais enzimas
são capazes de clivar in vitro imunoglobulinas bovinas [Talbot et al, 1991]. Outro mecanismo de
escape reside na internalização de anticorpos aderidos à superfície do parasita [Granger &
Warwood, 1996]. Por último, T. foetus também é capaz de remodelar sua superfície induzindo,
assim, a variação antigênica dificultando à imunocompetência por parte do hospedeiro. Tal
variabilidade antigênica é detectada em Tf190, TF-LPG e TF1.17 [Shaia et al., 1998; Singh et al.,
2001]. Além disso, verificou-se que T. foetus secreta um antígeno solúvel (SGA) que também se
apresenta imunogenicamente diferente e cujo papel na etiologia da tricomonose ainda é
desconhecido [Singh et al., 2001].
4.2.4) Interação do parasita com a MEC.
O trato reprodutivo de fêmeas bovinas consiste, basicamente, em uma cavidade envolta por
camadas de células epiteliais estratificadas e escamosas revestidas por muco. Logo abaixo do
epitélio vaginal se encontra a MB. Mecanismos de adesão do parasita à células e a MEC são
primordiais para a persistência num ambiente em constante transformação, devido ao longo ciclo
menstrual. Desta forma, assume-se que após digerir mucinas, destruir o epitélio de revestimento

23
via citólise ou rompimento de suas junções oclusivas, T. foetus entre em contato a MEC e ali
possa se associar.
Diversas proteínas plasmáticas, incluindo fibronectina (FN), podem ser encontradas
associadas à superfície de T. vaginalis [Peterson & Alderete,1982]. A ligação de FN com T.
vaginalis parece ser mediada por proteínas de superfície, uma vez que o pré-tratamento deste
parasita com tripsina inibe em até 85% tal associação [Crouch & Alderete, 1999]. Tanto T.
vaginalis como T. foetus são capazes de degradar FN, mas a relevância fisiológica de tal
atividade ainda não está totalmente esclarecida [Talbot et al, 1991].
Interação específica e saturável com LMN-1 foi também observada tanto em T. vaginalis
quanto em T. foetus [Silva Filho et al, 1988; Silva Filho et al, 2002]. No caso do patógeno
humano esta interação foi melhor caracterizada, demonstrando-se que o reconhecimento de
LMN-1 por T. vaginalis ocorre preferencialmente pelo peptídeo YIGSR [Silva Filho et al., 1998].
Foi também identificada na superfície de T. vaginalis uma proteína de 118KDa capaz de se ligar
a LMN-1 [Silva Filho et al, 1988]. No entanto, carboidratos de superfície parecem também estar
envolvidos na interação de T. vaginalis com LMN-1, uma vez que o pré-tratamento do parasita
com periodato reduz em 55% a capacidade de associação deste parasita com LMN-1 imobilizada
[Crouch & Alderete, 1999]. T. foetus apresenta uma maior ingestão de partículas de poliestireno
quando estas se encontram recobertas por LMN-1 ou FN o que corrobora a existência de
glicoconjugados reconhecedores destas moléculas na superfície de T. foetus [Benchimol et al.,
1990].
A associação de T. vaginalis à LMN-1 imobilizada predominantemente resulta em drásticas
modificações morfológicas deste parasita, o qual passa a apresentar forma amebóide, espraiada
sobre o biofilme de LMN-1, sugerindo a ocorrência de adesão estável [Crouch & Alderete, 1999;
Silva Filho et al, 1988]. Acredita-se que este processo de transformação morfológica esteja
relacionado a sinalização intracelular decorrente da ligação LMN-1 à superfície do parasita. No
entanto, além da participação de proteínas G, pouco se sabe sobre os processos moleculares
envolvidos [Lal et al., 2006].
Componentes da MEC podem influenciar a interação parasito-célula hospedeira de diversas
formas. Em T. foetus foi demonstrado que na presença de LMN-1 a capacidade do parasita de se
aderir a células epiteliais é significativamente aumentada [Silva-Filho et al., 1988]. Foi também
observado que a associação de T. vaginalis a LMN-1 imobilizada induz a expressão de uma

24
atividade de metaloprotease, a qual, por sua vez é similar à metaloproteases de matriz [Silva
Filho et al., 1999], sugerindo que a ligação de T. vaginalis ao biofilme de LMN-1 induza à
degradação ou remodelamento da MEC promovendo, assim, ao menos potencialmente, à invasão
tissular no hospedeiro.
Crouch e Alderete (1999) além de indicarem que a interação de T. vaginalis com FN ou
LMN-1 parece ser mediada por mecanismos distintos, demonstraram que os “receptores” de
superfície envolvidos são diferentes daqueles identificados como adesinas no processo de
citoadesão.

25
4) Leishmania (L.) amazonensis
5.1) As leishmanioses
O táxon Leishmania inclui protozoários parasitas causadores de um grupo de zoonoses
denominadas leishmanioses e possuem como reservatórios pequenos mamíferos (roedores,
marsupiais e canídeos).
As leishmanioses consistem de um amplo espectro clínico de enfermidades que afligem a
espécie humana e que se revelam como manifestações clínicas divididas em dois grupos:
Leishmaniose visceral ou calazar. Neste caso, os parasitas apresentam acentuado tropismo
pelo sistema fagocítico mononuclear do baço, do fígado, da medula óssea e dos tecidos linfóides.
A forma clínica da doença varia desde quadros assintomáticos até casos graves que, quando não
tratados, levam o paciente à morte. A sintomatologia clássica da doença é caracterizada por febre,
hepatoesplenomegalia, anemia, leucopenia e caquexia.
Leishmaniose tegumentar. Grupo de enfermidades onde ocorre a infecção de células do
sistema fagocítico mononuclear da pele e mucosas. Os agentes etiológicos da leishmaniose
tegumentar americana são espécies pertencentes aos complexos mexicana e brasiliensis (tabela1).
Dependendo da Leishmania spp envolvida e do estado imunológico do paciente podem ser
observadas diferentes formas clínicas:
- Leishmaniose cutânea localizada (LCL). Os parasitos infectam os macrófagos no sítio
de inoculação e sua intensa multiplicação resulta numa destruição celular e reação
inflamatória local. A inflamação evolui para necrose, formando ulceras indolores, rasas
ou fundas, de bordas salientes onde se concentram os parasitos, e fundo granuloso com
ou sem exudação. A cicatrização é lenta, podendo ocorrer num período entre 6 meses a
vários anos.
- Leishmaniose cutânea disseminada (Borderline). Se inicia com o aparecimento de
lesões epitelias típicas de LCL, no entanto, após um tempo de aparente cura (meses até
ano) observa-se a formação de lesões satélites devido a ocorrência de disseminação
linfática ou hematogênica dos protozoários.
- Leishmaniose mucocutânea ou cutâneomucosa. Neste caso ocorre o comprometimento
das mucosas. Inicialmente, tem-se a formação de lesões características da leishmaniose
cutânea, mas devido a uma resposta imune celular exarcebada do paciente surgem

26
lesões desfigurantes, com baixa densidade de parasitos. É comum a ocorrência de
invasão hematogênica ou linfática, geralmente para a região oral ou nasofaríngea. O
paciente pode apresentar dificuldades de respirar, falar e de se alimentar, podendo
chegar ao óbito.
- Leishmaniose cutânea difusa. Após o surgimento da lesão inicial surgem várias lesões
metastásicas, devido principalmente à disseminação linfática. A forma cutânea difusa
se manifesta em pacientes que apresentam uma resposta imune celular anérgica ou com
histórico de infecção com calazar. Ao invés de lesões ulcerosas são observados nódulos
ricos em parasitos. Esta forma da doença tem um curso crônico progressivo e é
extremamente refratária à quimioterapia.
Nos últimos anos foi registrado mundialmente um aumento significativo de áreas endêmicas
para leishmaniose, acompanhado por aumento marcante no número de casos documentados da
doença, atingindo aproximadamente 12 milhões de pessoas espalhadas em 88 paises, dos quais 72
são países em desenvolvimento [Desjeux, 2004]. A importância da doença também cresce à
medida em que seu caráter oportunista em pacientes aidéticos vem sendo descrito, sendo o Brasil
possuidor do maior número de casos de co-infecção HIV- Leishmaniose (Boletim anual da
OMS).
Existem cerca de 20 espécies de Leishmania já identificadas como causadoras de infecções
humanas (Tabela 1).
Posição sistemática e classificação do gênero Leishmania, adaptado de Mendonza-León e
colaboradores (1996). Esta classificação agrupando as espécies em complexos foi proposta por
Lainson e Shaw, em 1972:
- Tabela 1 –
*termo sem valor taxonômico.
Gênero: Leishmania Sub-gêneros: Leishmania (L.) Viannia (V.) Complexos*: L. donovani L. tropica L. mexicana L. braziliensis
Espécies:
L. (L.) donovani L. (L.) tropica L. (L.) mexicana L. (V.) brasiliensis L. (L.) infantum L. (L.) aethiopica L. (L.) amazonensis L. (V.) peruviana L. (L.) chagasi L. (L.) major L. (L.) pifanoi L. (V.) panamensis L. (L.) archibaldi L. (L.) garhami L. (V.) guyanensis L. (L.) venezuelensis L. (L.) venezuelensis L. (V.) lainsoni L. (V.) naiffi L. (V.) shawi L. (V.) colombiensis

27
O diagnóstico da leishmaniose é bastante amplo, envolvendo técnicas laboratoriais como (a)
coloração de Giemsa e posterior visualização ao microscópio óptico, de material obtido de
punção ou raspado da borda de lesões cutâneas ulceradas; (b) cultivo in vitro ou inoculação em
animais; (c) reação de Montenegro (teste intradérmico), também conhecido como teste da
leishmanina, onde o antígeno proveniente de formas promastigotas mortas é injetado na face
anterior do antebraço, sendo a reação positiva indicada pelo aparecimento de uma pápula ou
nódulo ou ulceração; (d) sorologia [Grevelink & Lerner, 1996]; (e) hibridização in situ e PCR.
No caso de regiões endêmicas, geralmente o diagnóstico é baseado em sinais clínicos.
Poucos avanços foram realizados no tratamento das leishmanioses, o qual é muitas vezes
ineficaz, associado a graves efeitos colaterais, e extremamente doloroso. As drogas de primeira
escolha são os antimoniais pentavalentes, como Glucantime® (antimoniato de N-metil-glucamina,
Rhône Poulene), e Pentostan® (estiboglucanato de sódio, Wellcome Foundation), que foram
introduzidos como quimioterápicos na década de 40 (Rath et al., 2003). O inconveniente no uso
destes medicamentos é a necessidade de tratamentos prolongados, o que resulta na acumulação
de metabólitos tóxicos, além de favorecer o surgimento de cepas resistentes e de ser injetável.
Outras drogas utilizadas no tratamento das leishmanioses são pentamidina, anfotericina B,
paromomicina e miltefosina [Rath et al., 2003]. Apesar de pacientes tratados apresentarem uma
cura clínica, acredita-se que não exista cura real para a leishmaniose. Parasitos viáveis podem ser
recuperados a partir de biópsias de lesões cicatrizadas por alguns anos, e o perfil nutricional e
imunológico dos pacientes pode permitir a manutenção de infecções sub-clínicas por mais de 30
anos [Saravia et al., 1990; Aebischer et al., 1993].
Como profilaxia, há a necessidade de se fazer um controle dos vetores e animais
reservatórios em regiões endêmicas, o que se constitui uma tarefa árdua, por ser dispendioso
financeiramente e trazer riscos ao equilíbrio ecológico.

28
5.2) Aspectos Gerais.
Leishmania (L.) amazonensis possui a seguinte identidade sistemática:
Reino: Protozoa
Filo: Sarcomastigophora
Super Classe: Mastigophora
Ordem: Kinetoplastida
Família: Trypanosomatidae
Gênero: Leishmania
Figura 4: Identidade sistemática de Leishmania (L.) amazonensis. Modelo esquemático da forma promastigota de L.
amazonensis. Esquema feito por Márcia Attias.
A Família Trypanosomatidae compreende um grande grupo de protozoários monoxênicos
(Angomonas, Blastocrithidia, Crithidia, Herpetomonas, Leptomonas, Rhynchoidomonas e
Wallaceina) e heteroxênicos (Endotrypanum, Phytomonas, Sauroleishmania e Trypanosoma),
sendo que o gênero Leishmania pertence, como discutido anteriormente, ao grupo dos
digenéticos. O gênero Trypanosoma, de extrema importância médica assim como o gênero
Leishmania, possui como principais representantes às espécies Trypanosoma cruzi e
Microtúbulos subpeliculares
Bolsa flagelar
Cinetoplasto

29
Trypanosoma brucei, agentes etiológicos da doença de Chagas e da doença do sono,
respectivamente.
Atualmente o gênero Leishmania subdivide-se em 2 subgêneros: Viannia e Leishmania. O
subgênero Viannia compreende apenas o complexo Leishmania braziliensis, encontrado nas
Américas tropical e subtropical, enquanto que o subgênero Leishmania é constituído pelos
complexos Leishmania donovani, Leishmania mexicana e Leishmania tropica, encontrados tanto
no Velho Mundo como no Novo Mundo (tabela 1). Esta classificação agrupando as espécies em
complexos foi proposta por Lainson e Shaw em 1972. É importante ressaltar que os complexos
não são considerados unidades taxonômicas formais, mas são muito úteis para agrupar diferentes
espécies de Leishmania que são semelhantes e apresentam algumas características peculiares.
Uma espécie de Leishmania de grande importância médica no Novo Mundo trata-se de espécie
Leishmania chagasi, pertencente ao complexo Leishmania donovani, sendo a espécie responsável
por todos os casos de leishmaniose visceral.
A espécie utilizada no presente estudo, Leishmania (L.) amazonensis pertence ao
complexo L. mexicana e é uma espécie causadora da leishmaniose cutânea localizada,
disseminada e difusa [Silveira et al., 2005]. Assim como os outros componentes do gênero,
apresenta duas formas evolutivas distintas: promastigotas, (células tipicamente alongadas,
flageladas e bastante móveis) e amastigotas (formas arredondadas, imóveis, intracelulares,
responsáveis pela manutenção da infecção no hospedeiro vertebrado) [Alexander &
Russel,1992].
A transmissão da leishmaniose é feita por insetos (vetores) do gênero Phlebotomus no Velho
Mundo, enquanto no Novo Mundo a transmissão ocorre principalmente por insetos dos gêneros
Lutzomyia e Psychodopygus. O parasita desenvolve-se extracelularmente no trato alimentar do
inseto vetor na forma promastigota, onde são reconhecidos 5 morfotipos distintos: promastigotas
procíclicos, nectomonas, haptomonas, paramastigotas e promastigotas metacíclicos [Bates, 1994].
O primeiro evento que ocorre no intestino médio do inseto vetor é a transformação das
amastigotas, ingeridas durante o repasto sanguíneo, em formas promastigotas procíclicos. Este
morfotipo é elíptico, com aproximadamente 6-8 μm de comprimento e divide-se rapidamente no
espaço delimitado pela membrana peritrófica do intestino médio. Gradualmente, os promastigotas
procíclicos vão se alongando e sofrendo modificações morfológicas passando pelos diferentes
morfotipos, finalmente se diferenciando para promastigotas metacíclicos. Este processo de

30
diferenciação em formas promastigotas metacíclicas é denominado de metaciclogênese. A forma
promastigota metacíclica é o estágio infectivo do parasito que será transmitido pelo inseto vetor
durante o seu repasto sanguíneo, esta forma é capaz de sobreviver às mudanças de ambiente
durante a infecção do hospedeiro vertebrado [Alexander et al., 1999]. Promastigotas metacíclicas
são encontradas no aparelho bucal, numa posição que facilita a transmissão na hora da picada. As
formas promastigotas metacíclicas irão infectar preferencialmente os macrófagos, fagócitos
profissionais do sistema fagocítico mononuclear, onde se diferenciarão em formas imóveis,
denominadas amastigotas, que se multiplicarão no interior de vacúolos parasitóforos
(fagolisossomo). Após sucessivos ciclos de divisão celular, o excesso de carga parasitária leva ao
rompimento da célula hospedeira e os amastigotas liberados vão infectar novos macrófagos
(Figura 5) [Handman & Bullen, 2002]. Atualmente se conhece a capacidade de L. amazonensis
infectar outros tipos celulares como células dendríticas, fibroblastos e até neutrófilos, mas a
célula hospedeira preferencial é o macrófago.
Figura 4: Ciclo de Vida de L. amazonensis. Adaptado de Vannier-santos et al, 2002.
Hospedeiro Invertebrado
Hospedeiro Vertebrado
Proliferação de procíclicos no intestino.
Migração de metacíclicos para partes do aparelho bucal.
Ligação inicial e fagocitose
Diferenciação para amastigotas no fagolisossomo
Proliferação dos amastigotas
Lise do macrófago
Re-infecção

31
Do ponto de vista morfológico, a membrana plasmática de L. amazonensis possui aspecto
trilaminar com espessura de aproximadamente 9nm e pode ser dividida em três regiões distintas:
(1) a região do próprio corpo celular, (2) da bolsa flagelar e, (3) do flagelo [Benchimol & de
Souza, 1980]. Existe uma diferença considerável na composição protéica da membrana
plasmática (MP) de formas infectivas e não-infectivas, além da forma amastigota. Pimenta e De
Souza (1983) demonstraram que a superfície de L. amazonensis contém tanto cargas positivas
quanto negativas e que durante a transformação amastigota-promastigota ocorre modificações na
carga de superfície do parasita. No caso de Leishmania e outros tripanosomatídeos, bem como
em fungos, não são capazes de sintetizar o colesterol sendo, portanto, o ergosterol o esterol
presente em suas membranas.
A Leishmania, assim como outros representantes da família Trypanosomatidae, possui logo
abaixo da sua membrana plasmática, uma camada de microtúbulos subpeliculares, extremamente
estáveis, e que confere aos representantes desta família uma eficaz proteção contra o estresse
mecânico. Estes microtúbulos são estruturas ocas compostas por treze protofilamentos, sendo que
cada microtúbulo conecta-se com seu adjacente através de pequenos filametos [Hunt & Ellar,
1974; Linder & Stalhelin, 1977; Dwyer, 1980].
A presença de microfilamentos de actina em formas promastigota e amastigota de
Leishmania foi caracterizada em diferentes sítios do parasito como falgelo, bolsa flagelar, núcleo
e cinetoplasto, bem como em diferentes faces das membranas celulares e também associada aos
microtúbulos subpeliculares, indicando que a actina está espalhada por toda célula
[Sahasrabuddhe et al., 2004].
O flagelo da Leishmania, localizado na região anterior do corpo celular, surge a partir de
um corpúsculo basal que conecta-se a membrana da mitocôndria próximo ao cinetoplasto, através
de pequenos filamentos, e emerge de uma invaginação da membrana plasmática chamada de
bolsa flagelar [De Souza, 1984]. É pelo flagelo, principalmente pela sua extremidade, que o
parasito irá se aderir no intestino do inseto vetor e preferencialmente na célula hospedeira. As
formas amastigotas possuem um curto flagelo internalizado na bolsa flagelar. Por isto esta forma
é imóvel; diferente do que observamos na promastigota, onde o flagelo confere motilidade ao
parasito.
Assim como outros membros da família Trypanosomatidae, a Leishmania amazonensis
possui uma mitocôndria única que se estende ao longo do corpo celular próxima à membrana

32
plasmática [De Souza & Benchimol, 1984]. Na matriz desta mitocôndria, localizada próxima ao
corpúsculo basal, apresenta um complexo arranjo de fibrilas de DNA em forma de bastão
conhecida como cinetoplasto. Esta estrutura é utilizada para definir todos os protozoários da
ordem Kinetoplastida.
Os megassomas são compartimentos lisossomais com a função de digerir material
extracelular internalizado seja por endocitose ou autofagia. O termo megassoma foi proposto por
estes autores tendo em vista a dimensão destas organelas, que muitas vezes pode ser maior que o
núcleo da célula, entretanto organelas com dimensões variadas também podem ser encontradas
[Alexander & Vickerman, 1975; Pupkis et al., 1986; Antoine et al., 1988]. As leishmanias, bem
como outras espécies do gênero, possuem também retículo endoplasmático liso e rugoso,
ribossomos, complexo de Golgi, formando seu sistema de síntese e glicosilação de proteínas e
lipídeos. O seu núcleo possui uma organização estrutural semelhante as das células eucarióticas
em geral.
5.3) A Interação Leishmania - Hospedeiro.
5.3.1) Reconhecimento, ligação e internalização pela célula hospedeira:
Leishmania sp, assim como outros parasitas intracelulares, utilizam-se de estratégias que
asseguram a sua sobrevivência em ambientes hostis, seja adaptando-se ao novo ambiente, seja
evitando tal ambiente. A adesão do parasita à superfície da célula hospedeira envolve vários
sistemas ligante-receptor. Esta adesão pode ocorrer através de moléculas adsorvidas na superfície
do parasito como anticorpos, fibronectina e de receptores para manose-6-fosfato e manose-fucose
[Vannier-Santos et al., 2002]. As formas promastigotas apresentam na sua superfície moléculas
como a glicoproteína de 63kDa (gp63), a qual é uma metaloprotease [Davies et al., 1990;
Pimenta et al., 1991], e o lipofosfoglicano (LPG), principal glicoconjugado de membrana
[Descoteaux & Turco, 1999, 2002], ambos desempenhando importante papel durante a interação
parasita - célula hospedeira [Rittig & Bogdan, 2000]. A gp63 é incorporada à membrana
plasmática das formas promastigotas por uma cauda de fosfatidilinositol e aproximadamente 10%
da superfície do parasito é ocupada por esta glicoproteína. O LPG é um glicolipídeo composto de
repetidas unidades de dissacarídeos ligados a uma cauda de fosfatidilinositol através de um

33
núcleo hexassacarídico, ocupando 25% da superfície total do parasito [Pimenta et al., 1989;
Descoteaux & Turco, 1999]. O LPG é estágio e espécie específico.
O reconhecimento parasito-célula hospedeira ocorre principalmente através de receptores
para o terceiro componente do sistema complemento, presentes na superfície do macrófago (CR1
e CR3). Estes são capazes de interagir diretamente com gp63 [Soteriadou et al., 1992], LPG
[Talamás-Rohana et al., 1990; Descoteaux & Turco, 2002] e proteofosfoglicanas (PPG) [Ilg,
2000], ou ainda, podem reconhecer os fragmentos C3b e C3bi, depositados na superfície do
parasito em função da ativação da cascata do complemento por glicoconjugados da superfície da
Leishmania [Mauël, 1996; Alexander et al., 1999; Cunningham, 2002]. Segundo Wright e
Silverstein (1983), a utilização dos receptores do sistema complemento não leva à indução da
explosão respiratória, favorecendo a sobrevivência intracelular do parasita. No caso de formas
amastigotas, as quais apresentam pouca ou nenhuma expressão de LPG, a interação parasita-
macrófago parece ser mediada por PPGs, moléculas ligantes de heparina e glicoesfingolipídeos
de superfície [Love et al., 1993; Strauss et al., 1993; Ilg, 2000]. O reconhecimento se dá através
de receptores para porção Fc de imunoglobulinas do tipo IgG (FcγR), além de CR3 e receptores
de manose do macrófago [Guy & Belosevic, 1993; Peters et al., 1995] Possivelmente receptores
do tipo Toll-like, primeiramente descritos em Drosophila [Medzhitov et al., 1997],
posteriormente encontrados em células de mamífero como macrófago e células dendríticas,
possam estar envolvidos na adesão da Leishmania aos macrófagos do hospedeiro vertebrado.
Estes receptores estão envolvidos na resposta imune inata e numa resposta imune não específica.
São fortes as possibilidades de Leishmania usar também estes receptores para se ligar aos
macrófagos e então levar a uma regulação de genes da resposta imunológica inicial, um
mecanismo ainda não elucidado [Awasthi et al., 2004].
A internalização de Leishmania se dá principalmente por fagocitose do tipo ‘zipper’, porém
são também observados eventos de fagocitose do tipo ‘coiling’ e macropinocitose. Os
protozoários então se mantêm em compartimentos acídicos limitados por membrana, de origem
fagolisossomal, denominados vacúolos parasitóforos [VPs – Chang, 1983]. As formas
amastigotas, adaptadas à vida intracelular, apresentam metabolismo acidófilo e, a presença de
sistemas de transporte de metabólitos com atividade ótima em pH ácido na membrana destes
microrganismos, garante ao parasito acesso aos nutrientes presentes no VP [Burchmore & Barret,
2001; Chang et al., 2003]. Provavelmente gp63 auxilia a forma amastigota a viver no ambiente

34
acídico do vacúolo parasitóforo produzido pela fusão com os lisossomos. Além disso, a
sobrevivência no ambiente inóspito do fagolisossoma é possível graças à capacidade do
protozoário de alterar a fisiologia da célula hospedeira em benefício próprio [Alexander et al.,
1999; Cunningham, 2002]. Apesar de apresentarem diversas características em comum, os VPs
que abrigam as diferentes espécies de Leishmania apresentam algumas diferenças fenotípicas. A
maioria das espécies de Leishmania, incluindo L. donovani e L. major, se mantêm em vacúolos
pequenos, ditos apertados, contendo um ou poucos parasitos. Já as espécies do complexo
mexicana L. amazonensis e L. mexicana induzem a formação de VPs espaçosos, ditos frouxos,
que podem conter dezenas de parasitos [Antoine et al., 1998]. Segundo Peters e colaboradores
(1997), a formação destes vacúolos largos por L. mexicana envolve a participação de
proteofosfoglicanas polianiônicas secretadas pelos amastigotas. Nos últimos 30 anos uma série de
trabalhos tem demonstrado que os VPs contendo Leishmania apresentam uma alta capacidade
fusogênica, característica que favorece a captação de metabólitos pelo parasita. Ligantes
internalizados por endocitose via receptor que geralmente se acumulam em lisossomas são
encaminhados para o VP em macrófagos infectados, indicando a capacidade destes vacúolos de
fundirem com vesículas da via endossomal/lisossomal [Alexander & Vickerman, 1975; Courret
et al., 2002].
Dentre as funções celulares alteradas no macrófago como conseqüência da infecção por
Leishmania spp. podemos citar: (i) inibição de diversas cascatas de sinalização intracelular
[Olivier et al., 1992] (ii) inibição da produção de agentes microbicidas, tais como radicais livres
de oxigênio [Descoteaux et al., 1992], radicais superóxido [Buchmüller-Rouiller & Mauël, 1987]
e óxido nítrico [Proudfoot et al., 1995]; (iii) aumento da produção de prostaglandinas e
leucotrienos, importantes imunomoduladores celulares [Belley & Chadee, 1995; Matte et al.,
1999]; (iv) redução da capacidade de apresentação de antígenos [Alexander et al., 1999]; (v)
redução marcante na expressão gênica dos macrófagos [Buates & Matlashewski, 2001]; (vi)
bloqueio do processo de apoptose [Moore & Matlashewski, 1994]; (vii) modulação da resposta
celular a citocinas [McDowell & Sacks, 1999; Ray et al., 2000].

35
5.3.2) A interação do parasito com o sistema imune do hospedeiro.
Aparentemente, o sistema imune do hospedeiro desempenha um papel regulatório na
expressão da doença, uma vez que a ativação dos macrófagos infectados pode provocar a morte
dos parasitos através da produção de espécies reativas de oxigênio e/ou óxido nítrico. A resposta
imune denominada celular relacionada com a ativação da população de linfócitos T (Th1) e a
produção de citocinas primárias como IL-12 e IFN-γ que em conjunto com outras citocinas levam
a ativação dos macrófagos que passam a apresentar antígeno e a produzir IL-12 [Croft &
Yardley, 2002]. A ativação dos macrófagos é o primeiro mecanismo para eliminar a Leishmania
via produção de metabólitos de oxigênio, num processo conhecido como explosão respiratória,
onde se observa a produção de ânions superóxido (O-2), peróxido de hidrogênio (H2O2) e óxido
nítrico (NO) [Assreuy et al., 1994].
Em camundongos, observou-se que duas populações de células T “helper”, tipo 1 (Th1) e
tipo 2 (Th2), parecem estar envolvidas na indução da resistência ou susceptibilidade à infecção.
As células Th1 produzem IL-12 e IFN-γ e são responsáveis pelo aumento da resposta imune
mediada por células através da ativação dos macrófagos, levando ao desenvolvimento de um
quadro clínico brando. Por outro lado, se houver um aumento da atividade das células Th2 que
secretam IL-4, IL-5 e IL-10, há um aumento da resposta denominada humoral e inibição de
algumas respostas mediadas por células, resultando quase sempre em infecção disseminada
[Grevelink & Lerner, 1996].
Evidências clínicas apontam para a ocorrência de destruição óssea e comprometimento
das articulações em pessoas infectadas com L. amazonensis com processo inflamatório
exarcebado, no entanto, os mecanismos relacionados ainda não são conhecidos [Costa et al.,
2006]
Tanto gp63 quanto LPG protegem as leishmanias do sistema imunológico. A
glicoproteína gp63 é capaz de degradar imunoglobulinas, fatores do complemento bem como
proteínas lisossomais [Chaudhuri & Chang, 1988]. As bases moleculares destes mecanismos de
proteção ainda não foram elucidadas. Quanto a participação do LPG na inibição da resposta
imune primeiramente sabe-se que ele é liberado da membrana da forma promastigota durante a
transformação para a forma amastigota, migrando então para a superfície dos macrófagos
infectados. Sabe-se que ele é capaz de inibir a explosão respiratória, inibir a atividade hidrolítica

36
de enzimas lisossomais, possivelmente por quelar o cálcio, bem como inibir a proteína cinase C
[Turco & Descoteaux, 1992].
Evidências apontam para a capacidade de L. amazonensis alterar diversas vias de
sinalização intracelular do macrófago hospedeiro. Macrófagos contendo amastigotas
internalizadas apresentam ativação de MAPK e ERK1/2 [Yang et al., 2007]. Essa ativação está
relacionada com a indução de secreção de IL-10 que por sua vez está relacionada com a inibição
da resposta celular e a casos clínicos mais severos [Karp et al., 1993]. É importante mencionar
que a ativação de MAPK e ERK1/2 pela interação com amastigoas de lesão não é suficiente para
induzir a produção de IL-10. Em combinação deve haver outro estimulo inflamatório como a
presença de ácido hialurônico de baixo peso molecular [Yang et al., 2007]. Ácido hialurônico
existe na MEC como fragmentos de alto peso molecular, no entanto, após uma injúria tecidual
pequenos fragmentos desta molécula podem ser encontrados levando a ativação de células do
sistema imune [Hodge-Dufour et al., 1997].
5.3.3) A interação com a MEC do hospedeiro.
A MEC é uma barreira para a infecção por Leishmania spp já que formas promastigotas
inoculadas na pele do hospedeiro vertebrado precisam ultrapassar a barreira epitelial e MB até
encontrar sua célula hospedeira, o fagócito mononuclear. Da mesma forma, os amastigotas
liberados pelo rompimento de macrófagos encontram a MB durante a procura por novos
macrófagos aos quais devem se ligar e promover internalização. Assim sendo, é de grande
importância para ambas as formas evolutivas apresentarem mecanismos específicos de interação
com componentes da MEC.
Regiões da derme infectadas com L. amazonensis apresentam perda da matriz do tecido
conjuntivo local. Abreu-Silva e colaboradores (2004) demonstraram que estas alterações na
organização da MEC são causadas diretamente pela degradação promovida por formas
amastigotas durante a re-infecção de macrófagos. Essas observações estão de acordo com a
atividade proteolítica típica de metaloprotease de matriz recentemente descoberta em gp63 [Mc
Gwire et al, 2003]. Tanto as formas de gp63 ancoradas na MP como as liberadas possuem
atividade de degradação de Colágeno tipo I e FN. No entanto, LMN parece ser resistente à
digestão por gp63. Acredita-se que essa atividade de degradação de componentes da MEC atue

37
como facilitador para a migração pela MB [McGwire et al, 2003], no entanto, a possibilidade de
estar ocorrendo uma remodelação da MEC não pode ser descartada, assim como seus efeitos
sobre as células do tecido e do sistema imune do hospedeiro.
Atividade de ligação a heparina foi também identificada na superfície de formas amastigotas
de L. amazonensis. No entanto, acredita-se que esta atividade esteja associada ao reconhecimento
de PTG do tipo heparan sulfato, bastante presente na superfície dos macrófagos [Love et al,
1993].
FN foi o primeiro componente da MEC a ser identificado como possível ligante para L.
amazonensis. Formas promastigotas e amastigotas reconhecem e se ligam a FN e essa associação
facilita o processo de fagocitose pelos macrófagos [Wyler et al, 1985]. Acredita-se que a FN
funcione como uma ponte molecular entre o macrófago e o parasito, uma vez que ambos
possuem receptores específicos para esta glicoproteína. O efeito de FN , assim como LMN, como
estimulante para o processo de fagocitose por macrófagos de eritrócitos recobertos com IgG ou
complemento já é bem conhecido [Bohnsack et al., 1985].
Promastigotas de L. mexicana são capazes de aderir especificamente a fibrilas de colágeno
tipo I por onde, aparentemente, se alinham e se movem com facilidade [Lira et al., 1997].
O reconhecimento de LMN foi descrito para L. donovani (protozoário causador da
leishmaniose visceral no Velho Mundo) [Ghosh et al., 1996]. Uma LBP de 67kDa foi
identificada e isolada da superfície de formas promastigotas deste parasita [Ghosh et al., 1999].
Trabalhos de caracterização desta associação demonstraram indícios de participação de cátions
divalentes e estimulação da ligação pela presença de zinco. O sítio de reconhecimento da LMN
utilizado pela LBP de 67kDa parece ser o pentapeptídeo YIGSR [Bandyopadhyay et al, 2002].
Esta LBP que parece ser uma proteína integral transmembrana teve seu domínio ligante de LMN
localizado na região extracelular N-terminal da proteína; a terminação citosólica C-terminal
também foi demonstrada [Bandyopadhyay et al., 2003]. A associação L. donovani - LMN via
LBP de 67kDa parece ser de grande importância para o processo infeccioso [Bandyopadhyay et
al., 2001]. Essa associação induz a ativação de tirosina quinases, sugerindo a indução de eventos
de sinalização secundários. Além disso, foi demonstrado que na presença de LMN o número de
parasitos por macrófago é significativamente aumentado [Bandyopadhyay et al., 2001].

38
Racional e Objetivos
O que tem nos chamado a atenção nos últimos anos é o fato de protozoários parasitas intra
e extracelulares interagirem com LMN tomarem destinos diferentes. Tanto o tropismo quanto as
invasões celular e tissular apresentados por protozoários parasitas pode ser, ao menos em parte,
devido a forma com que interagem com glicoproteínas de matriz extracelular (MEC): (a) sítios
reconhecidos no(s) ligante(s); (b) tipo(s) de complexo(s) receptor empregado(s) no
reconhecimento; (c) via(s) de processamento intracelular da associação complexo receptor-MEC
e (d) respostas dos parasitas tanto à química quanto à topografia apresentada pelo ligante (LMN
no presente caso).
No presente trabalho, abordamos algumas dessas etapas: o reconhecimento de LMN
imobilizada por parte de T. foetus e L. amazonensis, a caracterização inicial do(s) receptor(es) e
as respostas morfológicas dos parasitas à associação com LMN, de forma a comparar estratégias
desenvolvidas por estes protozoários extra e intra-celulares decorrentes de suas interações com
LMN-1. Portanto, este trabalho tem como objetivo analisar de forma comparativa a interação de
T. foetus (modelo extracelular) e L. amazonensis (modelo intracelular) com a glicoproteína
majoritária da MB LMN-1.
Especificamente, os objetivos deste trabalho são:
• caracterizar o processo de interação entre T. foetus e formas amastigotas e promastigotas
de L. amazonensis com LMN-1 imobilizada.
• caracterizar o(s) receptor(es) para LMN presentes nos protozoários parasitas.
• investigar a influencia da ligação à LMN-1 na citopatogenicidade de T. foetus e
• investigar a influência da associação de L. amazonensis à LMN-1 na interação de formas
promastigotas com macrófagos peritoniais.

39
Metodologia
1) Parasitas.
1.1) Tritrichomonas foetus
Utilizamos microorganismos da cepa K de T. foetus [Silva Filho et al, 1982], isolada pelo Dr.
Hélio Guida (Projeto de Patologia Animal, EMBRAPA), do trato urogenital de um touro
infectado. Formas trofosoítas foram cultivados axenicamente em meio TYM (Diamond, 1957),
suplementado com 10% de soro fetal bovino (SFB), a pH 6.8, por 24 horas e a 37 °C com 5%
CO2.
1.2) Leishmania (L.) amazonensis
L. amazonesis da cepa WHOM/BR/75/Josefa, foi mantida por infecções sucessivas induzidas
nas patas traseiras de camundongos Balb/C e os parasitas resultantes coletados como previamente
descrito [Rabinovitch et al., 1986].
1.2.1) Formas amastigotas de lesão.
Formas amastigotas foram isoladas de lesões feitas nas patas traseiras de camundongos
Balb/C infectados. A infecção foi feita por inoculação de promastigotas metacíclicos infectivos
na concentração de 105 células/mL, usando seringa descartável de 1 mL no coxim das patas
traseiras. Cerca de um mês após a inoculação as lesões começaram a aparecer. Para isolar as
lesões, os animais foram mortos em câmara de CO2, as lesões retiradas e maceradas em Potter
estéril de forma a provocar o rompimento dos macrófagos e conseqüente liberação das formas
amastigotas; o rompimento foi acompanhado através de microscopia óptica de contraste de fase.
Após o rompimento, o homogeneizado total foi centrifugado por duas vezes a 211g por 10 min 4
°C para a remoção de restos de tecido. O sobrenadante foi então cetrifugado a 1900g por 15 min
a 4 °C, onde no “pellet” foram obtidas as formas amastigotas parcialmente purificadas. As formas
amastigotas foram então isoladas por centrifugação em gradiente de Percoll como adaptado de
Hart e colaboradores (1981) e utilizadas nos experimentos.

40
1.2.2) Formas promastigotas.
As formas promastigotas foram obtidas a partir das formas amastigotas de lesão e cultivadas
axenicamente em meio Warren (BHI, ácido fólico e hemina) suplementado com 10% de SFB, por
4 dias a 28 °C. Todos os ensaios foram feitos com formas consideradas infectivas isto é, até 5
passagens em cultura.
2) Células-alvo.
2.1) HeLa
Células epiteliais da linhagem HeLa foram cultivadas em meio RPMI 1640, acrescido de 10% de
SFB, em garrafas plásticas de cultura à 37°C e 5% de CO2. Após confluência, as células foram
separadas utilizando solução de tripsina 0.2%, EDTA (0,58mM) em tampão fosfato de sódio
(0.01M), por 5 min à 37°C. A atividade enzimática foi interrompida pela adição de SFB e as
células foram, então, coletadas e cultivadas em placas de 96 poços (105células/poço).
2.2) Macrófagos
Macrófagos foram retirados da cavidade peritonial de camundongos BalbC, machos, através da
aspiração de fluido peritoneal previamente injetado com meio RPMI sem soro e, então,
plaqueados em lamínulas redondas de vidro dentro de placas de 24 poços. Após 50 min de
interação em meio RPMI sem soro as células não aderidas foram removidas por lavagens com
PBS e, então, os macrófagos aderidos foram cultivados em meio RPMI, com 10% de SFB, por
24h, 37°C e 5% CO2.
3) Proteína purificada e peptídeos sintéticos.
A Laminina utilizada em todo o trabalho foi a isoforma 1 isolada de tumor EHS de
camundongos (Invitrogen), enquanto que os peptídeos sintéticos (Tabela 2) foram cedidos pelo
Dr. Motoyoshi Nomizu (Universidade de Hokkaido, Japão).

41
Tabela 2: Peptídeos sintéticos de LMN utilizados (Nomizu et al., 1995, 1997, 1998)
4) Incorporação de 3H-timidina por T. foetus
Formas trofosoítas de T. foetus marcados com [3H]-timidina foram obtidos de cultivo feito
em 10mL de meio TYM completo contendo 30μCi [3H]-timidina, “overnight”, a 37°C. As
células foram, então, lavadas para a retirada do marcador radioativo não incorporado e ensaiadas.
5) Associação de T. foetus com LMN-1 imobilizada.
Biofilmes de LMN-1 foram preparados em lamínulas de vidro redondas. Às lamínulas foi
adicionada a solução de LMN-1 (5, 10, 20 e 30μg/ml), por 2 horas, à 37 °C feita em tampão
HEPES free acid 50mM, pH 6.6, acrescido de 10mM de cada um dos seguintes cloretos: CaCl2,
MnCl2, MgCl2. T. foetus previamente marcados com [3H]-timidina (2.105 parasitas/ml) foram
colocados para interagir com LMN-1 imobilizada por 1 hora, a 37 °C, em solução tamponada
com HEPES (10mM), pH 7.2, acrescido de sacarose (125mM) e cátions divalentes (CaCl2,
MnCl2, MgCl2) a 10mM, em placas de 24 poços. Como controle negativo, lamínulas sem a
proteína imobilizada foram utilizadas.
Para a análise da participação de cátions divalente 2.105 parasitas/ml foram colocados
para interagir com lamínulas revestidas com 20μg/ml de LMN-1, como previamente descrito, na
presença ou não de cátions divalentes.
A análise da participação de açúcares na ligação com LMN foi feita por ensaios de adesão
exatamente como descrito acima, em solução tamponada com HEPES (10mM) pH 7.2, acrescido
de cátions divalentes (CaCl2, MnCl2, MgCl2) a 10mM, na presença de β-Lactose (125mM) ou de
sacarose (125mM).
Após 3 lavagens com PBS para a retirada dos parasitos não aderidos, aqueles aderidos às
lamínulas foram solubilizados com SDS 2% e o produto resultante foi ressuspenso em liquido de
cintilação (Cytoscint). A radioatividade resultante, equivalente aos parasitas que se mantiveram
Posição Seqüência Seqüência ativa
A-13 RQVFQVAYIIIKA Não conhecida
A-208 AASIKVAVSADR IKVAV
AG-73 RKRLQVQLSIRT LQVQLSIR
C-16 KAFDITYVRLKF YVRL

42
aderidos às lamínulas, foi medida por cintilação líquida. Os ensaios apresentados representam um
ensaio representativo de três repetições.
6) Associação de formas amastigotas e promastigotas de L. amazonensis a LMN-1 imobilizada.
2.106 promastigotas/poço ou 3.106 amastigotas/poço foram colocados para interagir com o
biofilme de LMN (20μg/ml) por 1 hora, a 37 °C, em solução tamponada com Hepes (50mM) pH
7.2, acrescido de sacarose (125mM) e de cada um dos seguintes cloretos (10mM): CaCl2, MnCl2,
MgCl2. Como controle negativo, o mesmo procedimento foi repetido com lamínulas sem o
biofilme de LMN. Também como controle, parasitos foram pré-incubados com LMN (20μg/ml)
por 30min a 4 °C e então colocados para interagir com o biofilme de LMN. Este ensaio foi
realizado com formas promastigotas vivas ou fixadas em glutaraldeído 2,5%.
Adicionalmente 2.106 promastigotas / poço foram colocados para interagir com lamínulas
revestidas com LMN (20μg/ml), como previamente descrito, na presença ou ausência de cada um
dos cloretos (10mM).
A análise da participação de açúcares na associação de L. amazonensis a LMN foi feita por
ensaios exatamente como descrito acima, na presença de β-Lactose (125mM) ou de sacarose
(125mM) em solução tamponada com Hepes (10mM) pH 7.2, acrescido dos seguintes cloretos
(10mM): CaCl2, MnCl2, MgCl2. Após lavagens em 0,145NaCl tamponado com 0.01M fosfato de
sódio (PBS), para a retirada das células não aderidas, os parasitas são fixados em glutaraldeído
(2,5v/v). A quantificação de parasitas associados a LMN imobilizada foi feita em microscopia
óptica de contraste de fase através de contagem do número de parasitas presentes em 30 campos
randomicamente escolhidos em cada lamínula em triplicata. Os ensaios apresentados são
representativos de três repetições.
7) Microscopia Eletrônica de Varredura (MEV)
Os protozoários (T.foetus e formas promastigotas de L. amazonensis) foram colocados
para interagir com lamínulas recobertas ou não com 20μg/ml de LMN-1, em solução tamponada
(Hepes 50mM) a pH 6.6, acrescido, de 10mM de cátions divalentes (Ca+2, Mn+2 e Mg+2) por 2
horas a 37°C e, então, lavados com PBS, fixados com glutaraldeído 2,5% e preparados para
MEV. Após fixação, as lamínulas foram pós-fixadas com tetróxido de ósmio (2%) e ferrocianeto
de potássio (0,8%) por 40 minutos e, então, lavadas em PBS (4x), desidratadas em concentrações

43
crescentes de etanol, e secas pelo método do ponto crítico (utilizando CO2 como fluido de
transição). Uma camada de ouro de 20nm de espessura foi aplicada às amostras, as quais foram
observadas em microscópio eletrônico de varredura (JEOL JSM-5310). Como controle, parasitos
foram fixados em glutaraldeído (2,5%) e adsorvidos à lamínulas de vidro previamente recobertas
com poli-L-lisina (0.1%).
8) Tratamento de T. foetus com periodato ou tripsina (adaptado de Crouch & Alderete, 1999).
Parasitas previamente incubados com [3H] timidina (30μCi) foram colocados para interagir
com LMN-1 imobilizada, como anteriormente descrito na seção 5, em três condições: (1)
controle, não tratados; (2) pré-tratados com periodato (1mM, por 30 minutos à 4°C em PBS) ou
(3) pré-tratados com tripsina (3mg/ml por 30 minutos à 37°C em PBS). Posteriormente, os
parasitas foram lavados três vezes em PBS e a radioatividade resultante quantificada por
cintilação líquida. Antes de cada experimento, testes de viabilidade (eritrosina B) foram
realizados demonstrando que em todos os tratamentos os parasitos se mantiveram com
viabilidade acima de 90%.
9) Competição com peptídeos sintéticos.
Os parasitos (T. foetus e promastigotas de L. amazonensis) foram pré-incubados com cada um
dos quatro peptídeos sintéticos (Tabela 2), separadamente e por 30min à 4°C e, então, colocados
para interagir com LMN-1 imobilizada por 1h à 37°C. Ensaios de competição foram também
feitos de forma que cada um dos peptídeos foram separadamente co-incubados com o parasita
durante o processo de interação com a LMN imobilizada. A quantificação de parasitas
associados à LMN imobilizada foi feita em microscopia óptica de contraste de fase, através da
contagem do número de parasitas presentes em 30 campos escolhidos aleatoriamente em cada
uma das três lamínulas. Os valores obtidos foram comparados com aqueles resultantes da
associação de parasitas a LMN imobilizada na ausência de competição com os peptídeos e
expressas na forma de porcentagem, segundo:
Porcentagem de associação = parasitas + peptídeos X 100 parasito - peptídeos

44
10) “Dot blot”.
Parasitas (T. foetus e promastigotas de L. amazonensis) foram incubados em tampão de lise
(Tris-HCl 100mM, NaCl 150mM, Glicerol 10% e Triton X-100 0,6%) (Galán et al, 1992) na
presença de um coquetel de inibidores de proteases (10μM de E64, 100μM de TLCK, 1mM de
PMSF, 10mm de EDTA, 10 mM de O-fenatrolina, 1μM de Pepstatina A). Fragmentos de
membrana de nitrocelulose (0,45μm) foram lavadas em tampão Tris 10mM, NaCl 190mM
(TBS). Extratos totais de T. foetus e L. amazonensis (amastigota e promastigota) foram aplicados
às membranas em pequenos pontos de 1μl por vez, em espaços demarcados e separados. As
membranas foram, então, incubadas “overnight” e a 4 °C com tampão TBST de bloqueio (Tris
10mM, NaCl 190mM, Tween 20 0,1% + leite em pó desnatado molico 5% + BSA 0.1%) visando
bloquear sítios de ligação inespecíficos. Em seguida foram lavadas com tampão TBST (Tris
10mM, NaCl 190mM, Tween 20 0,1%) e, então, incubadas com LMN-1 (20μg/ml) por 2 horas
sob agitação suave e em tampão de bloqueio sem Tween 20 e com cátions divalentes (TBS,
1%BSA e MgCl2, MnCl2, CaCl2 a 10mM). Da mesma forma, as membranas foram lavadas e
incubadas com anticorpo primário anti laminina (1:100), diluído em tampão de bloqueio TBST.
Após nova lavagem as membranas foram incubadas com o anticorpo secundário conjugado a
peroxidase (1:8000) por 1 hora também em TBST de bloqueio. Para a retirada de resíduos
contaminantes, as membranas foram sucessivamente lavadas com TBST por 3 horas. Como
controle negativo, as mesmas amostras foram processadas como acima descrito mas sem
proceder a incubação com LMN-1 (20μg/ml). Neste caso, a incubação com o anticorpo primário
e secundário foi feita sem nenhuma modificação.
Extratos totais de T. foetus e de promastigotas de L. amazonensis foram submetidas a
tratamentos com periodato (10, 50 e 100mM) ou tripsina (10, 20, 100mg/ml) e então aplicados às
membranas. As últimas foram sequencialmente incubadas com LMN-1, anticorpo primário e
secundário, exatamente como descrito acima.
A revelação dos produtos associados as membranas foi feita com sensibilização de filme
fotográfico estimulado por ação do ECL (“enhancement of electrochemiluminescence") em
câmara escura.
11) Extração de proteínas e SDS-PAGE

45
Parasitas (T. foetus e promastigotas de L. amazonensis) foram incubados em tampão de lise
(Tris-HCl 100mM, NaCl 150mM, Glicerol 10% e Triton X-100 0,6%) (Galán et al, 1992) na
presença de um coquetel de inibidores de proteases (10μM de E64, 100μM de TLCK, 1mM de
PMSF, 10mm de EDTA, 10 mM de O-fenatrolina, 1μM de Pepstatina A). Ao extrato obtido de T.
foetus foi adicionado ácido tricloacético (TCA, 10%) para a precipitação de proteínas [Grimbleby
e Ntailianas, 1961]. Após a precipitação que se deu por incubação “overnight” a 4 °C, as
proteínas resultantes foram ressuspensas em tampão de dissolução (Tris 16,5mM,
Duodecilsulfato de sódio (SDS; 0,5%), β mercaptoetanol 5%, pH 6,8) e quantificadas segundo
método colorimétrico de Bradford (1976). As amostras (T. foetus e L. amazonensis) foram
ajustadas para 3mg/ml, em tampão de amostra de Laemmli (1970), na ausência de β-
mercaptoetanol e sem ferver para minimizar a desnaturação das proteínas. Em seguida, elas
foram separadas por SDS-PAGE, em géis contendo 7,5% de poliacrilamida.
12) Imunoeletrotransferência.
As proteínas separadas por SDS-PAGE foram transferidas para membranas de
nitrocelulose de 0,45μm (Amershan Life Science, Buckighamshire, England) [Towbin et al,
1979], em cuba de transferência semi-seca (BioRad). Após a transferência eletroforética, as
membranas foram incubadas em tampão de bloqueio (TBST, leite em pó desnatado molico 5% +
BSA 0.1%) a 4 °C, “overnight”. As demais etapas de lavagem, incubações e revelação, seguiram-
se exatamente como descrito para os casos de “Dot Blot”.
13) Co-imunoprecipitação
Extratos totais de culturas de promastigotas de L. amazonensis foram obtidos por lise em
solução apropriada (Nonidet P40 1% + Tris HCl 50mM + NaCl 150mM) acrescida de coquetel
de inibidores de protease (PMF, E64, Pepstaina, O-fenantrolina, TLCK e NaF). Ao extrato total
foi acrescentado LMN-1 (20μg/ml) por 2h, a 4°C. 100μl de proteina G conjugada a Sepharose
(Zimed) foi acrescentada ao extrato, “overnight” e a 4°C (pré-lavagem). Concomitantemente, 10
μl do anticorpo primário previamente descrito foram pré-incubados com 100 μl de proteina G
conjugada a Sepharose, “overnight” e a 4°C (pré-ligação). O sobrenadante da amostra (“pre-
cleaning”) foi incubado com o “pellet” resultante da ligação do anticorpo com a proteína G

46
conjugada a Sepharose (pré-ligação) “overnight” e a 4°C. Em seguida, procedeu-se sucessivas
lavagens em PBS gelado e a amostra foi fervida por 5 min e preparada para SDS-PAGE como
anteriormente descrito no item 11, no entanto, com a utilização de β-mercaptoetanol no tampão
de amostra. As bandas resultantes foram visualizadas após coloração do gel pela prata.
13) Ensaios de citotoxicidade
Culturas confluentes de células HeLa feitas em placas de 24 poços foram incubadas com 4
μCi de [3H]- timidina / poço, “overnight”, a 37°C e 5% CO2 [Singh et al., 1999]. Em seguida as
células foram lavadas por 3 vezes em PBS, a temperatura ambiente, a fim de se retirar o excesso
de traçador. 106 trofoítos de T. foetus /ml foram então ressuspensos em meio RPMI sem soro e
colocados para interagir com as células HeLa, na presença ou ausência de LMN-1 (20µg/ml), por
5 horas [Melo-Braga et al., 2003]. Posteriormente, o meio de interação foi recolhido e a
radioatividade presente foi quantificada por cintilação líquida. As marcações (cpm) resultantes
foram convertidas para valores percentuais comparando-os aos lisados totais de células HeLa que
não entraram em contato com T. foetus. Como controle negativo, foi avaliada a liberação
espontânea do traçador radioativo por células HeLa incubadas em RPMI sem soro por 5 horas.
14) Interação de promastigotas de L. amazonensis com macrófagos peritoniais.
Culturas de macrófagos foram plaqueadas em lamínulas de vidro adsorvidas em placas de
24 poços. A essas culturas foram adicionadas ou não formas promastigotas de L. amazonensis, as
quais haviam sido pré-tratadas ou não com LMN (20μg/ml), por 5min. A quantificação de
parasitos associados aos macrófagos foi feita por contagem em microscópio óptico de campo
claro após fixação e coloração das células com Giensa. Foram contados o numero de
promastigotas associados a cada 100 macrófagos e por lamínula foram quantificados 400
macrófagos. Os valores expressos como resultados, dizem respeito a experimentos feitos em
triplicata e o ensaio demonstrado é representativo de três repetições.

47
Resultados
1) Associação de T. foetus à LMN-1 imobilizada.
Para analisarmos a especificidade do processo e determinarmos a quantidade ideal de
LMN-1 para os ensaios de associação, T. foetus pré-marcados com [3H]-timidina foram
colocados para interagir por 1 hora com lamínulas de vidro recobertas ou não com 5, 10, 20, 30
ou 40 μg/ml de LMN-1. A quantificação de parasitos associados foi, então, aferida pela
quantidade do traçador radioativo presente nas amostras.
A curva de associação resultante apresentou saturação na quantidade de parasitos
associados à LMN-1 imobilizada a ≥ 20 μg/ml. 20 μg/ml de LMN-1 imobilizada foi então
utilizada nos demais ensaios.
Figura 1: associação de T.foetus pré-marcados com [3H]-timidina à lamínulas de vidro recobertas ou não
com LMN-1. A quantidade de parasitas associados à lamínulas contendo LMN-1 imobilizada encontra-se
expressa em cpma (radioatividade detectada por cintilação líquida). Barras representam desvio padrão de
três ensaios feitos em triplicata.
020406080
100120140160180
0 10 20 30 40 50
concentração de LMN (μg/ml)
cpm
a
LMN μg/ml

48
2) Associação de formas promastigotas e amastigotas de L. amazonensis a LMN-1 imobilizada
em lamínulas de vidro.
Formas promastigotas e amastigotas infectivas (Figuras 2 e 3, respectivamente) de L.
amazonensis foram colocadas para interagir por 1 hora com 20μg/ml de LMN-1 previamente
imobilizada em lamínulas de vidro. Tanto os promastigotas vivos colocados em contato com o
biofilme de LMN-1 como aqueles fixados, mortos, apresentaram aumento na associação com a
proteína (Figura 2). Tal aumento foi ainda mais pronunciado em formas amastigotas isoladas que
apresentaram associação quase nula ao vidro não recoberto com LMN-1. Na presença de LMN-1
a quantidade de parasitas associados pôde ser aferida (Figura 3). Buscando entender a
especificidade deste fenômeno, parasitas (promastigotas metacíclicos e amastigotas) foram
colocados para interagir com a proteína imobilizada. Como resultado, a capacidade inicial de
associação dos parasitas a LMN-1 imobilizada foi fortemente inibida, tanto nos ensaios com
promastigotas como naqueles realizados com as formas amastigotas.

49
Figura 2: Ensaio de ligação de formas promastigotas de L. amazonensis com LMN-1. As barras
representam erro padrão ( *, p < 0,05, teste t de Student). Experimento representativo de três repetições.
Figura 3: Ensaio de ligação de formas amastigotas de L. amazonensis com LMN-1. ( *, p < 0,05, teste t de
student). As barras representam erro padrão ( *, p < 0,05, teste t de Student). Experimento representativo
de três repetições.
Fixed parasites
3,475
12,975
28,325
0
5
10
15
20
25
30
35
para
site
s/fie
ldNon-fixed parasites
4,175
15,575
29,475 control withoutLaminin coatedsurface
Laminin coatedsurface
Lamininincubation +Laminin coatedsurface
Fixed parasites
3,475
12,975
28,325
0
5
10
15
20
25
30
35
para
site
s/fie
ldNon-fixed parasites
4,175
15,575
29,475 control withoutLaminin coatedsurface
Laminin coatedsurface
Lamininincubation +Laminin coatedsurface
Controle. Adesão sem o biofilme de LMNAdesão ao biofilme de LMN
Parasitas vivosParasitas fixados
Adesão ao biofilme de LMN c/ pré incubação c/ LMN
Para
sita
s/ca
mpo
*
*
*
*
0
10
20
30
40
50
60
70
Sem LMN-1 LMN-1 pré-incubação comLMN-1
amas
tigot
as p
or c
ampo
*
*

50
3) Morfologia dos parasitas associados a LMN-1.
Com o intuito de analisarmos possíveis alterações morfologicas de T. foetus e formas
prosmatigotas de L. amazonensis adquirida durante interação com LMN-1 monitoramos o
processo por MEV. T. foetus e a forma promastigota de L. amazonensis foram colocados para
interagir com LMN-1 imobilizada e, então, processados para MEV. Como controle, parasitos pré-
fixados foram adsorvidos à lamínulas previamente revestidas com poli-L-lisina. Como pode ser
observado na figura 4, a associação ao biofilme de LMN-1 não induziu nenhuma modificação
morfológica em L. amazonensis. Por outro lado, T. foetus apresentou maior número de formas
arredondadas, espraiadas, sobre lamínulas contendo biofilme de LMN-1 do que à lamínulas não
recobertas com a proteína. Foram contados 10 campos de mesma magnitude (1000X),
diferenciando-se células pouco aderidas (não amebóides) daquelas altamente espraiadas
(amebóides). Como pode ser observado na tabela abaixo, a proporção de T. foetus amebóides
obtidos na presença de LMN-1 foi significativamente maior daquela encontrada nas amostras
controle.
Tabela 3: Proporção de T.foetus pouco associados (não amebóides) e aderidos (amebóides) a lamínulas
revestidas ou não (controle) com LMN-1.
Não amebóide Amebóide
controle 88,40% ± 30,2 11,60% ± 6,2
Laminina 24,30% ± 4 75,70%± 21,7

51
Figura 4: MEV de formas promastigotas de L. amazonensis associadas ao vidro (A) ou a biofilme de LMN-1 (B), e
T. foetus associados ao vidro (C, E) ou aderidos ao biofilme de LMN-1 (D, F).
A B
C D
E F

52
4) Análise da natureza química da ligação T. foetus / L. amazonensis - LMN-1.
Uma vez sugerida a capacidade de T. foetus e L. amazonensis (Figura 1 e 2) de reconhecer
LMN-1, procedemos a identificação da natureza das moléculas de superfície destes parasitas
envolvidas no fenômeno. Adicionalmente observamos que na presença de 10mM de cada um dos
cloretos de CaCl2, MnCl2 e MgCl2 a capacidade de T. foetus de se associar a LMN-1 foi
significativamente aumentada (Figura 5A). Como controle, parasitas foram colocadas para aderir
à LMN-1, em solução tamponada, na presença dos cátions citados e acrescida de 5mM de EDTA.
Nesta condição o numero de parasitas associados a LMN-1 foi parcialmente decrescido. Por
outro lado, a presença de β-lactose (açúcar reconhecido por lectinas ligantes a LMN-1) na
solução de interação não influenciou significativamente a associação de T. foetus à LMN-1
(Figura 5B).
Nos ensaios realizados com a forma promastigota de L. amazonensis observamos que na
ausência dos cátions divalentes (10mM de CaCl2, MnCl2 e MgCl2) a associação do parasita à
LMN-1 foi quase anulada (Figura 6A). Por outro lado, a presença de β-Lactose não influenciou
significativamente a associação dos promastigotas a LMN-1 (Figura 6B).

53
Figura 5: Associação de T. foetus a LMN-1 imobilizada na presença de (A) cátions divalentes (Ca+2, Mg+2
e Mn+2) ou do quelante EDTA + cátions e (B) β-lactose ou sacarose. ( *, p < 0,05, teste t de Student).
Barras representam desvio padrão. Ensaios conduzidos em triplicata.
Adesão de T. foetus ao biofilme de laminina-1
0,001,002,003,004,005,006,007,008,009,00
10,00
controle EDTA Ca, Mg, MnTota
l de
célu
las
ader
idas
(x10
4 )
Adesão de T. foetus ao biofilme de laminina-1
0
1
2
3
4
5
Sacarose galactoseTota
l de
célu
las
ader
idas
(x10
4 )
A
B β- lactose
*
*
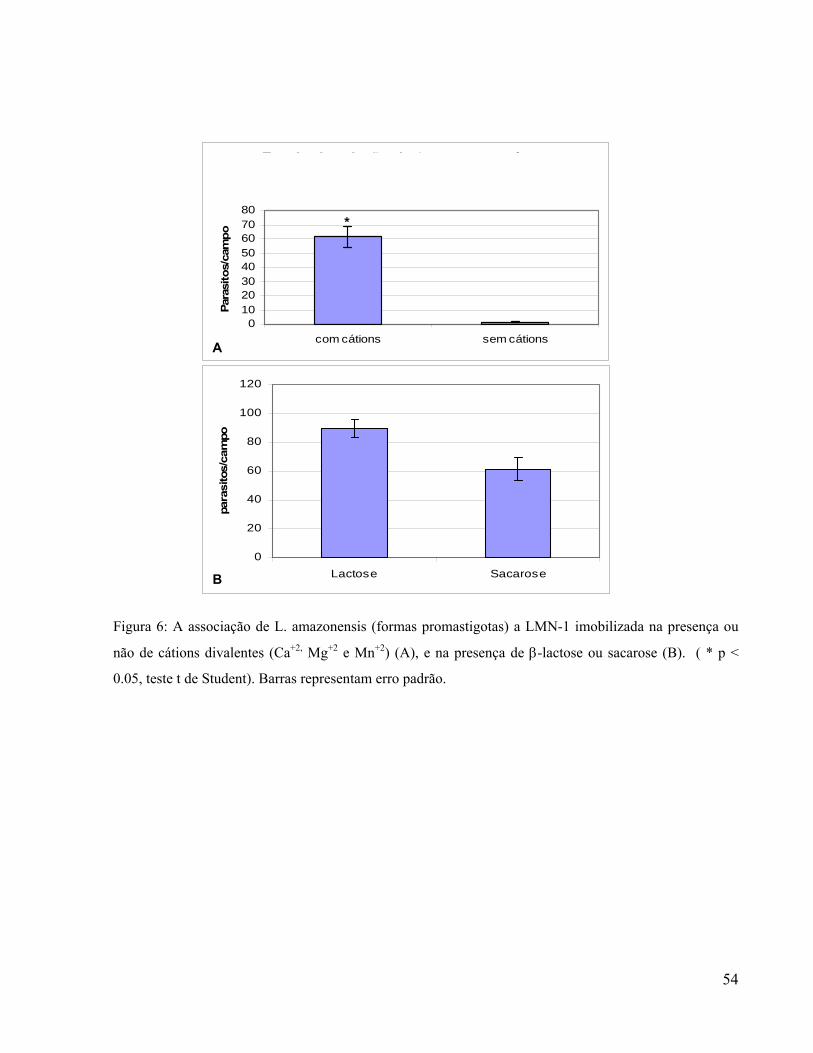
54
Figura 6: A associação de L. amazonensis (formas promastigotas) a LMN-1 imobilizada na presença ou
não de cátions divalentes (Ca+2, Mg+2 e Mn+2) (A), e na presença de β-lactose ou sacarose (B). ( * p <
0.05, teste t de Student). Barras representam erro padrão.
Ensaio de adesão de L. amazonensis ao LMN-1
01020304050607080
com cátions sem cátions
Para
sito
s/ca
mpo
0
20
40
60
80
100
120
Lactose Sacarose
para
sito
s/ca
mpo
A
B
*

55
Passamos a investigar se a associação de T. foetus à LMN-1 ocorria via porções
oligosacarídicas ou protéicas de superfície do parasita. Realizamos ensaios de associação a LMN-
1 com parasitas pré-tratados ou não com periodato de sódio ou tripsina. Como pode ser
observado na figura 7, o tratamento com tripsina reduziu em cerca de 50% a capacidade original
de associação de T. foetus a LMN-1. No entanto, o tratamento com periodato não resultou em
mudança significativa na taxa de associação do parasita ao biofilme de LMN-1.
Figura 7: Associação de T. foetus pré-tratados ou não com 1mM periodato de sódio ou 10mg / ml tripsina
à lamínulas de vidro recobertas com LMN-1. As barras representam desvio padrão ( * p < 0,05, teste t de
Student).
0%
20%
40%
60%
80%
100%
120%
Controle Periodato Tripsina
Ass
ocia
ção
(%)
*

56
5) Ocorrência de receptores para LMN-1 em T. foetus e L. amazonensis.
Ensaios de “Dot Blot” foram feitos utilizando-se extratos totais de parasitos reagidos com
20μg/ml de LMN-1, seguido por incubação seqüencial com anticorpos anti –LMN1 e secundário
conjugado a peroxidase. Como controle positivo, 3μg de LMN-1 foram aplicadas às membranas.
Como pode ser observado na figura 8A, T. foetus e as formas amastigota e promastigota de L.
amazonensis possuem em seus extratos totais moléculas que reagem com LMN-1.
A natureza química dessas moléculas foi ainda investigada a partir de extratos de T. foetus
pré-tratados com tripsina (10, 50 ou 100mg/ml) ou periodato de sódio (10, 20 ou 50mM).
Verificamos que o tratamento com a protease diminuiu progressivamente a marcação original
(Figura 8B). Por outro lado, o pré-tratamento com periodato de sódio não resultou em alteração
da reação.
De forma semelhante àquela realizada com T. foetus, extratos totais de formas
promastigotas de L. amazonensis foram pré-tratados com tripsina (100mg/ml) ou com periodato
de sódio (50mM). Tal qual observado com T. foetus, o tratamento com tripsina levou a anulação
da marcação, enquanto que o tratamento com periodato não resultou em nenhuma alteração na
intensidade dos “spots” (Figura 8B).
A
Laminina (controle)
L. amazonensis (promastigota)
T .foetus
L. amazonensis (amastigota)
Controle

57
Figura 8: “Dot Blot” indireto onde membranas de nitrocelulose contendo amostras em duplicata dos
diferentes parasitas foram sequencialmente incubadas com LMN–1, anticorpo anti LMN-1 e anticorpo
secundário conjugado com peroxidase (A) ou com amostras de formas promastigotas de L. amazonensis
tratadas ou não (CTL) com tripsina (100mg/ml) por 15 min a 37°C ou periodato (50mM) por 30min a 4°C
e com amostras de T. foetus foram tratadas ou não (CTL) com tripsina (100, 50 e 10mg/ml) ou periodato
(5, 10, 20 e 50mM). Todos os controles negativos foram feitos em ausência de pré-incubação com LMN-
1.
B
CTL
Periodato (mM)
100 50 10
Tripisina (mg/ml)
5 10 20 50
Tritrichomonas foetus
Leishmania (L.) amazonensis
CTL Tripsina Periodato

58
6) Seqüências de reconhecimento utilizadas por T. foetus ou L. amazonensis no reconhecimento a
LMN-1 imobilizada.
Com o intuito de identificarmos o(s) sítio(s) de LMN-1 reconhecidos por T. foetus ou L.
amazonensis, realizamos ensaios de competição com peptídeos presentes nas diferentes cadeias
(α, β e γ) de LMN-1. Estes ensaios consistiam em medir a associação dos parasitas ao biofilme
de LMN-1 na presença ou não de cada um dos peptídeos listados na Tabela 2. Os ensaios foram
feitos de duas formas: (a) pré-incubando-se os parasitos com cada um dos peptídeos ou (b)
acrescentando os peptídeos no momento da interação dos protozoários com LMN-1. Tanto as
formas promastigotas infectivas de L. amazonensis (Figura 9) como trofozoítas de T. foetus
(Figura 10) tiveram suas associações ao biofilme de LMN-1 significantemente reduzidas na
presença de cada um dos peptídeos. Tal inibição foi observada tanto quando os parasitas foram
pré-tratados com os peptídeos quanto quando os peptídeos foram adicionados (competição) no
momento da interação com LMN-1.
Figura 9: Ensaio de competição de formas promastigotas L. amazonensis com LMN-1. Os peptídeos
A208, C16, A13 e AGM3 foram utilizados para tratamento dos parasitas (pré-incubação) ou adicionados à
solução de interação (competição). As barras representam erro padrão ( * p < 0,05, teste t de Student).
Ass
ocia
ção
(%)
10,13%
100%
7,53%
18,22% 18,13%
38,25%
20,29%
30,89%
14,71%
0%
20%
40%
60%
80%
100%
120%
controle
AG73
C16
A208
A13
Pré-incubação
Competição
** *
*
*
**
*

59
3,67% 7,33% 9%
100%
12,33%
49%
10,67%
29% 20,33%
0%
20%
40%
60%
80%
100%
120%
para
site
s/Fi
eld
controlAG73C16 A208A13
Figura 10: Ensaio de competição de T. foetus com LMN-1. Os peptídeos A208, C16, A13 e AGM3 foram
utilizados para tratamento dos parasitas (pré-incubação) ou adicionados à solução de interação
(competição). As barras representam erro padrão ( * p < 0,05, teste t de Student).
Pré-incubação
Competição
Asso
ciaç
ão (%
)
*
* * *
*
*
**
e

60
7) Identificação de proteínas ligantes a LMN presentes em T. foetus e em L. amazonensis.
Ensaios de eletrotransferência para membranas de nitrocelulose a partir de géis de SDS-
PAGE foram realizados para este fim. As membranas contendo as bandas dos extratos proteícos
de T. foetus foram incubadas sequencialmente com LMN-1, anticorpo anti-LMN1 e anticorpo
secundário conjugado a peroxidase.
Encontramos 5 bandas referentes a proteínas reconhecedoras de LMN-1: de
aproximadamente 220, 200, 130, 125 e 80kDa (Figura 11 coluna B). Como controle negativo, a
incubação com LMN-1 foi omitida e nenhuma marcação foi encontrada (Figura 11 coluna A).
Figura 11: Imunoeletrotransferência indireta de extratos de T. foetus. Membranas de nitrocelulose
contendo a transferência eletroforética foram incubadas com LMN–1, anticorpo anti LMN-1 e anticorpo
secundário conjugado com peroxidase. Controle foi feito por omissão de incubação com LMN-1 (coluna
A).
A B
250 KDa
150 KDa
100 KDa
75 KDa

61
Para a identificação de moléculas ligantes a LMN-1 em L. amazonensis fez-se necessário uma
co-imunoprecipitação onde o extrato total de formas promastigotas infectivas foram
primeiramente colocados para interagir com LMN-1 e, então, incubados com anticorpo anti-
LMN-1 associada à proteína G conjugada a Sepharose. Dentre as diversas bandas visualizadas na
amostra resultante, apenas 4 não apresentaram massas moleculares compatíveis com as bandas
relativas ao anticorpo associado a proteína G-Sepharose (coluna 1) ou a LMN-1 desnaturada
(coluna 2). Essas 4 bandas são, portanto, possíveis moléculas receptoras de LMN-1 (setas
vermelhas).
Figura 12: Co-imunoprecipitação de extratos totais de promastigotas de L. amazonensis com LMN-1
(coluna 3). Como controle, amostra do “pre-binding” (anticorpo associado a proteína G conjugada a
sefarose) e da LMN-1 foram desnaturados e separados eletroforeticamente (colunas 1 e 2
respectivamente). Padrão de peso molecular está representado pelas setas verdes e as bandas específicas
da amostra de L. amazonensis pelas setas vermelhas.
80KDa
49KDa
28.9KDa
20,6KDa
1 2 3

62
8) Influencia da ligação de LMN-1 à superfície de T. foetus na toxicidade do parasita a células
HeLa.
A fim de investigarmos o papel funcional da ligação de LMN-1 à superfície de T. foetus
na interação do parasita com células de mamíferos, realizamos ensaios de citotoxicidade com
células epiteliais HeLa. Monocamadas celulares foram marcadas com [3H]-timidina e
subsequentemente incubadas com parasitos na presença ou ausência de LMN-1. Após 5 horas de
interação em meio sem soro a lise celular foi avaliada através da liberação do traçador radioativo
no sobrenadante do co-cultivo. Como resultado, observamos que na presença de LMN-1 a
capacidade citopatogênica de T. foetus foi significativamente aumentada (Figura 13). Como
controle, monocamadas radioativamente marcadas e cultivadas nas mesmas condições descritas
acima, porém na ausência T. foetus, tiveram a liberação espontânea de [3H]-timidina analisada.
Figura 13: Ensaio de citotoxicidade de T. foetus a monocamadas de células HeLa marcadas com timidina
tritiada, na presença ou ausência de 20μg/ml de LMN-1. A taxa de citotoxicidade expressa é relativa a
contagem do traçador radioativo liberado no sobrenadante de cultura, como detalhado em Materiais e
Métodos. Barras representam desvio padrão ( * p < 0,05, teste t de Student).
0,00%
5,00%
10,00%
15,00%
20,00%
25,00%
CTL T.foetus T.foetus + LMN
Efei
to c
itotó
xico
*
*

63
9) Influencia da ligação de LMN-1 na associação de L. amazonensis a macrófagos.
O papel funcional da ligação de LMN-1 à superfície de L. amazonensis foi analisado. Para
isto, realizamos ensaios de associação L. amazonensis a macrófagos peritoneais de camundongos
na presença ou ausência de LMN-1. Promastigotas infectivos pré-incubados ou não com LMN-1
foram colocados para interagir com macrófagos (relação parasito-célula de 10:1) e após 5 min de
interação as células foram lavadas, fixadas e coradas. A quantificação dos parasitas associados
aos macrófagos foi feita por contagem em microscópio óptico de campo claro (v. Materiais e
Métodos). A quantidade de parasitos por macrófago mostrou-se significativamente maior na
presença de LMN-1 (Figura 14).
Figura14: Ensaio de associação de formas promastigotas infectivas de L. amazonensis a macrófagos
peritoniais de camundongos na presença (LMN) ou não (CTL) de LMN-1 (20μg/ml). Promastigotas
infectivos foram colocados para interagir com macrófagos (relação parasito-célula de 10:1). Após 5 min
de interação as células foram fixadas, coradas e quantificadas como detalhado em Materiais e Métodos.
Barras representam erro padrão ( * p < 0,05, teste t de Student).
0
0,2
0,4
0,6
0,8
1
1,2
CTL LMN
prom
astig
otas
/mac
rofa
go *

64
Discussão
A cada ano se observa um aumento no número de publicações relacionadas ao
reconhecimento de componentes da MEC por patógenos extra e intracelulares [Lima et al.,
2005]. Evidências de que infecções por L. amazonensis acarretam em modificações na MEC na
região de infecção são fortes indícios da interação in vivo deste patógeno com a MEC de seu
hospedeiro [Abreu-Silva et al., 2004]. No entanto, até o momento pouco se sabe sobre como este
parasita interage com a MEC hospedeira.
No presente trabalho demonstramos que tanto as formas extracelulares (promastigotas)
quanto as intracelulares (amastigotas) de L. amazonensis são capazes de reconhecer LMN-1. A
forma promastigota é a forma mais estudada em relação à interação com a MEC, uma vez que se
trata da forma extracelular infectante no hospedeiro vertebrado e, portanto, supomos, possui
capacidade de atravessar a MB e MEC do tecido conjuntivo onde, então, ocorre o encontro com
macrófagos. Por outro lado, pouco se sabe sobre a interação de formas amastigotas com o meio
extracelular do hospedeiro. Contudo, o processo de manutenção da infecção é realizado por essas
últimas formas, no ambiente da MEC, onde amastigotas “liberadas” por macrófagos devem
interagir com novos macrófagos para o sucesso da infecção.
A forte capacidade de amastigotas de lesão de reconhecerem e de se associarem a LMN-1
imobilizada é o primeiro indício da importância desta interação amastigota-MEC para a
manutenção da infecção no hospedeiro vertebrado. É importante aqui comentar que a forma
amastigota utilizada nesta dissertação foi retirada de lesão em patas de camundongos BalbC e
portanto, trata-se de amastigotas retiradas de macrófagos rompidos. Isto é, amastigotas que
apresentam suas membranas “contaminadas” com restos de membrana do vacúolo parasitóforo.
Portanto, não podemos afirmar que a associação aqui demonstrada é produto da interação de
moléculas da membrana do parasita com LMN-1 ou de moléculas presentes em restos de
membrana do vacúolo parasitóforo que se encontram associadas à membrana do parasito. De
qualquer maneira, o método de obtenção dos parasitas a partir de lesão é a forma de se obter
amastigotas que mais se assemelham com aquelas encontradas in vivo. É justamente com esses
restos de membrana que as formas amastigotas entram em contato com o ambiente extracelular
do hospedeiro.

65
Como esperado para um parasita extracelular que habita a cavidade vaginal onde o
contato com a MB ocorre durante o processo invasivo de destruição da mucosa e barreira
epitelial, T. foetus é capaz de se associar ao componente majoritário da MB, LMN-1 [Silva-Filho
et al., 1988]. O reconhecimento molecular pertinente parece ocorrer de forma específica, isto é,
envolvendo associação receptor-ligante.
A curva de associação de T. foetus com LMN-1 imobilizada apresentou saturação na
quantidade de proteína igual ou maior que 20μg/ml sendo, portanto, esta a concentração ideal de
LMN-1 para os ensaios aqui descritos. Esse resultado está de acordo com Silva-Filho e
colaboradores (1988) onde também se demonstrou que 20μg/ml de LMN-1 foi a concentração
ótima da proteína para ensaios de associação T. foetus- LMN-1.
Morfologicamente, T. foetus apresenta uma drástica transformação quando em contato
com LMN-1 imobilizada. Tal transformação, de formas pirifomes alongadas para formas
amebóides, espraiadas, contendo diversas projeções citoplasmáticas (figura 4 E e F) sugere a
ocorrência de uma adesão molecular de T. foetus a este componente da matriz. Tal fato a nós nos
sugere que T. foetus não apenas reconhece e se ancora ao ligante, mas também re-estrutura seu
citoesqueleto de actina e ocorre disparo de cascata de sinalização. Para formas promastigotas de
L. amazonensis nenhuma mudança morfológica foi observada pelo contato deste microrganismo
com LMN-1. Este resultado nos sugere que a força de adesão de L. amazonensis a MEC é baixa,
resultando num processo simples de associação, e não de adesão ou fixação ao ambiente
extracelular.
A força de fixação ao hospedeiro priorizada pelo patógeno extracelular pode estar
relacionada à necessidade de sobrevivência de T. foetus quando submetido à forças mecânicas e
hidrodinâmicas (fluxo mestrual, por exemplo) o que não parece necessário para parasitas
intracelulares. Tal diferença entre associação e adesão célula-MEC hospedeira deve ser
“percebida” por outros patógenos intracelulares e extracelulares.
A conformação apresentada por LMN-1 imobilizada nas lamínulas de vidro é de grande
relevância na análise da associação LMN-1-parasitas. LMN-1 pode se auto-polimerizar
apresentando estruturas finais diferentes, de acordo com o pH ou presença de cátions na solução
de imobilização [Freire & Coelho-Sampaio, 2000; Yurchenco & Shan Cheng, 1993]. A
conformação de LMN-1 é de extrema importância para ensaios de ligação e adesão uma vez de
acordo com a conformação final adquirida pela proteína polimerizada pode ocorrer a não

66
exposição de sítios de ligação existentes na conformação funcional da molécula ou, por outro
lado, exposição de sítios que naturalmente se encontrariam cripticos. No presente trabalho houve
a preocupação de se fazer a adsorção de LMN-1 em tampão de pH ácido, já que este parece
induzir a auto-polimerização de LMN-1 [Freire & Coelho-Sampaio, 2000 ]. Por outro lado,
cátions divalentes (Ca+2, Mg+2 e Mn+2) foram acrescidos ao tampão de polimerização uma vez
que a presença deles, em especial o Ca+2, foi demonstrada ser de grande importância para a
manutenção da conformação funcional de LMN-1 [Yurchenco & Shan Cheng, 1993]. Ainda
assim, a conformação final de LMN-1 obtida em nossos ensaios pode apresentar importantes
diferenças daquela encontrada in vivo, uma vez que o modo da organização supra-estrutural, isto
é a estrutura final encontrada pela mistura de todos os componentes da MEC, gera epítopos de
ligação à adesinas celulares que não estão presentes na superfície de polímeros feitos com apenas
uma única molécula da MEC. Por outro lado, essa agregação com outros componentes da MEC
pode esconder sítios de ligação encontrados nos polímeros simples de LMN-1[Hansen et al.,
2006].
Uma vez reconhecida a capacidade destes parasitas com vias de infecção tão diferentes de
reconhecer LMN-1 imobilizada, passamos a nos concentrar numa caracterização molecular dos
processos envolvidos no fenômeno. Existe uma grande variedade de receptores para LMN
descritos até o momento [Mercúrio, 1995]: integrinas, lectinas e outras proteínas ligantes a LMN
ou LBPs. Além das integrinas, outras proteínas ligantes a LMN encontradas em patógenos
apresentam conformação dependente de cátion. Seja individualmente ou combinados, cátions
divalentes são requeridos para a funcionalidade de diversos tipos de receptores de componentes
da MEC [Wang et al., 1994; Mercúrio, 1995; Plow et al., 2000]. Esses cátions participam da
ligação tanto de T. foetus como de L. amazonensis à LMN-1. Na presença de 10mM de Ca+2,
Mn+2 e Mg+2 a capacidade de associação de T. foetus a LMN-1 foi significativamente aumentada.
Por outro lado, quando incubado em solução com os cátions divalentes e EDTA, a associação de
T. foetus à LMN-1 imobilizada foi reduzida, mostrando a especificidade do fenômeno. Em
conjunto, estes resultados sugerem a participação de receptor dependente de cátions no
reconhecimento de LMN-1 por T. foetus.
Curiosamente, na ausência dos cátions divalentes a capacidade de adesão de formas
promastigotas de L. amazonensis à LMN foi fortemente inibida. Para promastigotas a presença
dos cátions divalentes, em conjunto ou individualmente, é não apenas importante mas essencial

67
para a funcionalidade do(s) receptor(es) envolvidos no processo de interação do parasita com
LMN-1, sugerindo que funcionalmente, os receptores para LMN-1 encontrados em T. foetus e L.
amazonensis são diferentes. É importante lembrar que a participação dos cátions divalentes pode
estar sendo requerida tanto pelas moléculas receptoras de LMN-1 presentes nestes patógenos,
quanto para a conformação funcional dos sítios de ligação contidos na própria molécula de LMN-
1. O sítio de ligação C16 presente na cadeia gama de LMN-1, por exemplo, depende de Ca+2 para
se apresentar na conformação ativa.
A influência de cátions divalentes pode ser também um indicativo da participação de
receptores do tipo integrina. No entanto, análises mais completas se fazem necessárias para
confirmar a participação de proteínas tipo integrina na associação destes microorganismos a
LMN-1 imobilizada.
Lactose é comumente utilizada para o bloqueio da ligação de receptores de LMN do tipo
galectina (ligates específicos de sítios galactosídicos) ao ligante [Elola et al., 2005; Wood et al.,
1990]. A presença de β-lactose no meio de interação não alterou de forma significativa a
capacidade de associação de T. foetus ou L. amazonensis a LMN-1. Assim, sugerimos a não
participação de lectinas ligantes de sítios galactosídicos dos parasitas no reconhecimento a LMN-
1. Não podemos, no entanto, descartar a possibilidade da participação de lectinas ligantes de
outros sítios na porção glicídica de LMN-1. Contudo, é importante ressaltar que os sítios
galactosídicos e lactosídicos são os mais encontrados neste tipo de glicoproteína.
Uma vez sugerida ser a porção protéica de LMN-1 reconhecida pelos parasitas, passamos
a analisar a natureza química das moléculas ligantes a LMN-1 presentes em T. foetus e L.
amazonensis. Sítios de LMN como o AG73 podem funcionar como lectinas, de forma a
promover associação com receptores glicídicos. O tratamento de T. foetus com periodato de sódio
(agente oxidante de açucares) não resultou em alteração significativa na capacidade de adesão do
parasita à LMN-1 imobilizada. Tal resultado sugere a não participação da porção oligosacarídica
de glicoconjugados de superfície nesta interação. Por outro lado, o tratamento de T. foetus com
tripsina levou a uma inibição da adesão do parasita a LMN-1 demonstrando a participação de
proteínas de superfície de T. foetus na adesão à LMN-1 imobilizada. No entanto, não descartamos
a possibilidade da participação da porção oligossacarídica de glicoproteínas da superfície de T.
foetus na interação com LMN-1, uma vez que a clivagem da porção protéica pela ação da tripina
causaria a clivagem do sítio de ancoragem, impedindo o seu funcionamento mesmo estando o

68
sítio de ligação na porção glicídica da molécula. Além disso, o tratamento com periodato de sódio
se limita a oxidação de cadeias de carboidratos acídicas. Portanto, somente estas podem ser
completamente excluídos de participação no processo de associação de T. foetus a LMN. Cabe
aqui ressaltar a possível não participação neste processo de resíduos de ácido siálico da superfície
de T. foetus sugerida por este ensaio. Tal característica demonstra diferença em relação a
interação de T. vaginalis a LMN-1, onde resíduos de ácido siálico parecem estar envolvidos
[Crouch & Alderete, 1999]. Desta forma, esses resultados mostram que essas duas espécies de
tricomonadídeos apresentam maneiras distintas de interagir com a MEC de seus hospedeiros.
Como tratamentos com periodato de sódio e tripsina podem ser extremamentes tóxicos
para os parasitas, apenas baixas concentrações podem ser utilizadas de forma a não comprometer
a viabilidade dos mesmos. Daí esse ensaio não haver sido realizado com L. amazonensis, uma
vez que não nos foi possível encontrar concentrações não tóxicas de periodato de sódio ou
tripsina a serem utilizadas com este parasita.
A demonstração de que a associação T. foetus e L. amazonensis à LMN-1 não se faz
apenas por ligações eletrostáticas, mas existe o envolvimento de moléculas ligantes de LMN foi
feita por analises bioquímicas de extratos dos microrganismos. Por análises de “dot blot” indireto,
tanto T. foetus quanto as formas amastigotas e promastigotas de L. amazonensis revelaram
possuir em seus extratos moléculas que reconhecem LMN-1. Esse ensaio foi realizado com cinco
amostras diferentes de cada forma evolutiva dos parasitas e aqui apresentamos apenas as
melhores marcações resultantes. Ensaios de “dot blot” foram também importantes para
investigarmos a natureza química das moléculas dos parasitas que reconheciam LMN, uma vez
que podíamos nesses ensaios utilizarmos concentrações mais altas de periodato e tripsina.
Extratos de T. foetus tratados com diferentes concentrações de periodato de sódio não revelaram
alteração de marcação para LMN-1, confirmando a não participação de sítios oligosacarídicos do
parasita no reconhecimento à proteína. Por outro lado, o tratamento dos extratos de T. foetus com
tripsina resultou em ausência de reação à LMN-1. Desta forma, confirmamos o envolvimento da
porção protéica de glicoproteínas de T. foetus no fenômeno de reconhecimento de LMN-1. O
mesmo procedimento conduzido com extratos de formas promastigotas de L. amazonensis
também resultou em ausência de alteração da marcação em extratos tratados com periodato e o
tratamento com tripisina resultou na anulação da marcação.

69
Uma vez determinada a participação de proteínas reconhecedoras de LMN-1 tanto em T.
foetus quanto em L. amazonensis e a evidente participação da porção protéica de glicoproteínas
dos parasitas na interação com LMN-1, passamos a analisar os possíveis sítios de reconhecimento
da molécula envolvidos no fenômeno. LMN-1 é uma proteína multidomínio e vários são,
portanto, os sítios de reconhecimento celular descritos na sua estrutura [Nomizu et al.,1995;
1997; 1998]. Os peptídeos YIGRS, IKVAV (A208), A13, C16, AG73 são os peptídeos mais
estudados como sítios de reconhecimento para diversas células de mamíferos.
O reconhecimento de LMN-1 por promastigotas de L. amazonensis foi significativamente
inibido na presença de cada um destes peptídeos, tanto quando pré-incubados com o parasita
quanto quando presentes no momento da interação. Os mesmos ensaios feitos com T. foetus
resultaram em grande inibição no reconhecimento do parasita a LMN-1. Estes resultados nos
sugerem que LMN-1 pode ser reconhecida por T. foetus e L. amazonensis via cada um dos
peptídeos. Os resultados obtidos pelos ensaios de competição com os peptídeos, isto é, quando
colocados no momento da interação com LMN-1, demonstram que tal inibição não foi fruto de
uma possível aglomeração destes peptídeos no tampão de interação, mas sim de suas associações
com os parasitas. É importante, no entanto, acrescentar que este tipo de ensaio não nos permite
excluir a possibilidade dos peptídeos sintéticos se ligarem a outras porções da LMN-1, de forma a
bloquear a ligação da proteína aos parasitas. No entanto, essa ligação peptídeo-LMN-1 só seria
possível ocorrer em ensaios de competição, e não em ensaios de pré-incubação onde os
microorganismos entram em contato com os peptídeos antes de levados a interagirem com LMN-
1.
O reconhecimento de múltiplos sítios de ligação contidos em regiões distintas de LMN-1
pode permitir a estes microorganismos monitorarem a integridade do ligantes. Através do
reconhecimento de múltiplos sítios em um mesmo ligante de MEC um parasita poderia, também,
ser capaz de reconhecer diferentes isoformas de uma mesma molécula e assim ampliar sua
capacidade de monitoramento do ambiente extracelular. É provável que não somente o
reconhecimento em si, mas as forças de adesão ou afinidades do(s) receptor(es) dos parasitas para
o ligante determinem a ativação de diferentes funções.
Conhecendo a capacidade de T. foetus de reconhecer quatro distintos sítios na molécula de
LMN-1, se fez necessário identificar o(s) receptor(es) do parasita responsáveis por tal
reconhecimento. Através de “imunoblotting” indireto detectamos a ocorrência de cinco bandas de

70
pesos aproximados de 80, 125, 130, 200 e 220KDa como possíveis receptores para LMN-1 em
T. foetus. A presença de múltiplas proteínas ligantes de LMN-1 em T. foetus pode explicar a
capacidade deste parasita de reconhecer múltiplos sítios de ligação na molécula de LMN-1. No
entanto, a presença de uma proteína promíscua ligante a LMN-1, isto é, capaz de se associar a
mais de uma seqüência de reconhecimento não pode ser excluída. A presença de cinco proteínas
reconhecedoras de LMN-1 em T. foetus sugere também a ocorrência de um complexo de ligação
onde receptinas, receptores e co-receptores poderiam atuar em conjunto ou sobre distintas partes
da mesma molécula. Tal complexo estaria envolvido no reconhecimento e na manutenção da
associação do ligante à superfície do parasita, em tempo necessário para que T. foetus
respondesse ao ligante via ativação de diferentes cascatas de sinalização.
No caso de L. amazonensis, a identificação de moléculas reconhecedoras de LMN-1 foi
feita por co-imunoprecipitação uma vez que ensaios de “imunoblotting” indireto não se
revelaram eficientes para este microorganismo. A co-imunoprecipitação permitiu a incubação de
LMN-1 com extratos de promastigotas de L. amazonensis na ausência de substâncias
desnaturantes, como SDS ou β-mercaptoetanol, os quais poderiam prejudicar a conformação
funcional dos sítios de ligação da proteína ligante a LMN-1. Diversas proteínas precipitaram na
associação a LMN-1/IgG/proteína G. Após análise dos controles contendo o “pré-binding”
(proteína G conjugada ao anticorpo anti-LMN-1) e amostras de LMN-1 previamente desnaturada,
nos foi possível identificar quatro bandas como possíveis proteínas reconhecedoras de LMN-1:
três entre 80 e 49KDa, e a quarta entre 28.9 e 20.6KDa. Em L. donovani foi identificada uma
proteína ligante a LMN-1 de 67KDa, semelhante em peso molecular as proteínas aqui
encontradas em L. amazonensis [Ghosh et al., 1996]. Assim como para T. foetus, a presença de
múltiplas proteínas reconhecedoras de LMN pode explicar a capacidade de L. amazonensis de
reconhecer múltiplos sítios de ligação na molécula de LMN-1. No entanto, não descartamos a
possibilidade destas quatro bandas serem, na verdade, subunidades de uma mesma proteína que
atua de forma promíscua, isto é, capaz de se ligar a mais de uma seqüência de reconhecimento em
LMN-1. Para uma análise mais profunda do(s) receptor(es) para LMN-1 presentes em
promastigotas de L. amazonensis, utilizaremos extratos de parasitos metabolicamente marcados,
de forma que, visualizaremos somente as bandas relativas a proteínas sintetizadas pelo parasita.
Uma vez verificada a capacidade de T. foetus de reconhecer e de aderir à LMN-1
imobilizada, passamos a questionar se essa associação teria apenas papel de adesão ou se a

71
ligação a LMN-1 seria capaz de influenciar a interação deste parasita com seu hospedeiro. É
conhecida a capacidade de alguns parasitas de interagirem com componentes da MEC de forma a
estimular seu “potencial” invasivo e patogênico [Franco et al., 2002; Li et al., 1995]. Para
verificar a influência da associação a LMN-1 na citopatogenicidade de T. foetus, ensaios de
citotoxicidade foram realizados na presença e ausência de LMN-1. Na presença da glicoproteína
a capacidade de destruição de células epiteliais HeLa por T. foetus foi significativamente
aumentada. Tal aumento significativo, a nós nos sugere que a associação de T. foetus a LMN-1
pode ser um fator de virulência. No entanto, a capacidade citotóxica de T. foetus não é,
naturalmente, dependente da associação a este componente da MEC. A fim de sobreviver no
hospedeiro T. foetus deve co-optar o ambiente a seu favor, desde seu primeiro contato com
células epiteliais na cavidade vaginal bovina e em seguida a MB subjacente. Assim, nos parece
que o efeito citopatogênico aqui observado é fruto apenas da interação do patógeno com uma
única glicoproteína da MB imobilizada, e nada podemos afirmar sobre este comportamento in
vivo onde, na verdade, este microorganismo encontra uma MEC tri – dimensional, composta por
diversas moléculas que podem sinalizar para várias alterações funcionais. Só em estudos com
MEC complexas, em 3D, poderíamos analisar de maneira detalhada as reais modulações
ocorridas pelo contato deste parasita com a MB do hospedeiro. De qualquer maneira, os
resultados aqui apresentados demonstram que o reconhecimento de LMN-1 por T. foetus pode
modular funções que influenciam a interação do parasita com o hospedeiro. Em outras palavras,
T. foetus é capaz de perceber proteínas do meio extracelular e a elas responder diferentemente.
Toda a interação deste microrganismo com as células epiteliais foi realizada em meio de cultura,
na ausência de soro fetal bovino, para evitar interferência de outras proteínas do soro que
pudessem se associar a LMN-1 ou ao próprio parasita. Como controle, células epiteliais foram
mantidas pelo mesmo tempo em meio sem soro demonstrando que o efeito citotoxico observado
não poderia ser resultante da ausência de soro.
No caso do modelo L. amazonensis procedemos, também, a investigação sobre o papel
funcional da ligação de LMN à superfície deste microorganismo. Na presença de LMN-1 o
número de promastigotas aderidos por macrófago foi significativamente aumentado,
demonstrando que LMN-1 pode atuar como facilitador desta interação. Em L. donovani foi
descrito a influência de LMN-1 como indutor da interação deste parasita com macrófagos
[Bandyopadhyay et al., 2001]. O papel de LMN-1 como facilitador da associação deste

72
microorganismo a sua célula hospedeira é bastante plausível, se considerarmos o fato de que
assim como aqui demonstrado para L. amazonensis, macrófagos também possuem receptor para
LMN-1 [Huard et al., 1986]. Portanto, essa glicoproteína poderia estar funcionando como ponte
molecular entre o parasita e sua célula hospedeira. LMN-1 poderia, também, atuar como indutor
de fagocitose por macrófagos, como descrito por Bohnsack e colaboradores (1985). Portanto, é
possível que pelo menos parte do efeito promotor de LMN-1 na interação entre L. amazonensis e
a célula hospedeira não seja fruto exclusivamente da modulação de funções do parasita, mas de
uma modulação de funções do macrófago em contato com a proteína. Novamente, fica difícil
analisar se tais funções associadas a LMN-1 existem realmente in vivo, ou se isto se trata de um
artefato causado pela utilização da proteína em 2D. De qualquer maneira, o ensaio aqui descrito é
um forte indicativo de como o contato com o meio extracelular pode ser importante no processo
de interação de L. amazonensis com o macrófago [Wyler et al., 1985].
O ensaio de interação de L. amazonensis com macrófagos foi realizado apenas durante
cinco minutos e, portanto, é relativo somente ao processo de reconhecimento e adesão que ocorre
no primeiro contato entre o parasito e a célula hospedeira. Para uma análise mais completa da
cinética de interação se faz necessário a utilização de tempos maiores de incubação parasita-
macrófago, onde poderíamos quantificar os parasitas aderidos e também aqueles internalizados
pelo macrófago. Assim, poderíamos analisar se LMN-1 atua somente como indutor de adesão ou,
também, como agente opsonizante; isto é, induzindo o processo de fagocitose de L. amazonensis
pelo macrófago. É importante também considerar-se que esse processo de indução de fagocitose
de L. amazonensis é um momento chave no processo de infecção, uma vez que dependendo da
maneira como este processo é conduzido pode acarretar na ativação do macrófago levando a
morte do parasita fagocitado. Portanto, é de extrema importância para a infecção por L.
amazonensis que este processo de fagocitose seja realizado de forma silenciosa. Assim, talvez o
papel de LMN-1 na interação L. amazonensis-célula hospedeira possa ocorrer de forma a ajudar a
modulação do silenciamento do macrófago, aumentando a chance de sobrevivência do parasita
internalizado. Portanto, a interação de formas promastigotas e amastigotas de L. amazonensis
com macrófagos na presença de LMN-1 e outros componentes de MEC, imobilizados em 3D,
pode vir a esclarecer detalhes sobre a indução da fagocitose e modulação da ativação do
macrófago.

73
Os ensaios de interação de L. amazonensis com macrófagos na presença de LMN-1 foram
realizados somente com a forma promastigota. Pouco se sabe sobre a influência de LMN-1 na
interação com formas amastigotas. Portanto, é necessária a ampliação das análises de interação
aqui feitas com outros componentes de MEC utilizando formas promastigotas e amastigotas.
A associação à molécula de LMN-1 pode influenciar o comportamento de patogenos de
diversas maneiras. O reconhecimento de LMN-1, assim como o de outro componente da MB,
pode sinalizar a indução de várias funções relacionadas a capacidade infectiva de parasitas como
adesão, migração e quimiotaxia, entre outros. Dessa forma, se faz necessário um estudo mais
amplo sobre como o contato de L. amazonensis ou T. foetus com LMN-1 pode influenciar na
modulação de funções importantes nos processos infecciosos destes parasitas. A capacidade de
invasão de MEC por T. foetus, por exemplo, deve ser analisada de forma a melhor compreender
como este parasita chega ao útero, impedindo a nidação.
É importante também analisar a influência de LMN-1 no contato de parasitas com células
do sistema imune. Assim como as demais células, as do sistema imune também tem suas funções
moduladas por contato com o meio extracelular [Morwood & Nicholson, 2006]. Desta forma, é
plausível considerar que o remodelamento da MEC processado por patógenos pode vir a modular
a expressão fenotípica de células do sistema imune, de forma a cooptar o meio extracelular a
favor dos patógenos. No caso de L. amazonensis tal processo poderia gerar ambientes onde a
produção de citocinas envolvidas na resposta humoral fosse estimulada, em relação a resposta
celular. A modulação do ambiente extracelular é, portanto, de grande importância para o sucesso
de processos infecciosos e pode vir a explicar, pelo menos em parte, a presença de pacientes com
quadros clínicos de severidade bastante diferentes.
Enfatizamos que o trabalho aqui apresentado, assim como a maioria dos dados publicados
até o momento sobre a interação de patógenos com a MEC, advém de experimentos realizados
com componentes de MEC imobilizados em 2D. Entretanto, a utilização de matrizes 3D se faz
necessário para análises mais completas sobre os reais papéis da interação parasito-MEC em
processos infecciosos.

74
Conclusões
Face aos resultados obtidos, concluímos que:
1. Trofozoítas de T. foetus e microorganismos das formas promastigota e amastigota de L. amazonensis associam-se a LMN-1 imobilizada via processo de reconhecimento ligante-receptor;
2. T. foetus e promastigotas e amastigotas de L. amazonensis possuem receptores ou
“sistema receptor” para LMN-1. Esse(s) receptor(es) são de natureza protéica e funcionalmente dependentes de Ca+2, Mg+2 e Mn+2;
3. Enquanto T. foetus apresenta cinco proteínas ligantes a LMN-1, formas promastigotas de
L. amazonensis parecem possuir quatro;
4. Seqüências de aminoácidos presentes nos peptídeos A208, A13, C16 e AG73 de LMN-1 podem ser reconhecidas por T. foetus e L. amazonensis;
5. A ligação de LMN-1 à superfície de T.foetus resulta em ativação de alteração morfológica
e aumento de toxicidade do parasita a células epiteliais HeLa.
6. Para a forma promastigota de L. amazonensis a ligação de LMN-1 estimula a associação do parasita à macrófagos peritoneais de camundongos.

75
Referências Bibliográficas: ABREU-SILVA AL, CALABRESE KS, MORTARA RA, TEDESCO RC, CARDOSO FO, CARVALHO LOP & GONÇALVES DA COSTA SC. 2004. Extracellular matrix alterations in experimental murine Leishmania (L.) amazonensis infection. Parasitology 128: 385-390. ADAMS JC & WATT FM. 1993. Regulation of development and differentiation by the extracellular matrix. Development 117: 1183-1198. AEBISCHER T., MOODY S. F. & HANDMAN E. 1993. Persistence of virulent Leishmania major in murine cutaneous leishmaniasis: a possible hazard for the host. Infection and Immunity. 61: 220-226. ALBERTS B, JOSHNSON A, LEWIS J, RAFF M, ROBERTS K & WALTER P. Molecular Biology of the Cell.2002. ALDERETE JF & GARZA GE. 1985. Specific nature of Trichomonas vaginalis parasitism of host cell surfaces. Infection Immunity 50: 701-8. ALDERETE JF, LEHKER MW & ARROYO R. 1995. The mechanisms and molecules involved in cytoadherence and pathogenesis of Trichomonas vaginalis. Parasitology Today 11: 70-74. ALEXANDER J & VICKERMAN K. 1975. Fusion of host cell secondary lysosomes with the parasitophorous vacuoles of Leishmania mexicana-infected macrophages. Journal of Protozoology 22: 502-508. ALEXANDER J & RUSSELL DG. 1992. The interaction of Leishmania species with macrophages. Advances in Parasitology 31:175-254. ALEXANDER J, SATOSKAR AR. & RUSSELL DG. 1999. Leishmania species: models of intracellular parasitism. Journal of Cell. Science. 112: 2993-3002. ANDERSON ML, BARR BC & CONRAD PA. 1994. Protozoal causes of reproductive failure in domestic ruminants. The Veterinary Clinics of North America Food Animal Practice 10: 439-461. ANTOINE JC, JOUANNE C, RYTER A & BENICHOU JC. 1988. Leishmania amazonensis: acidic organelles in amastigotes. Experimental Parasitology 67: 287-300. ANTOINE, J-C., PRINA, E., LANG, T. & COURRET, N. 1998. The biogenesis and properties of the parasitophorous vacuoles that harbour Leishmania in murine macrophages. Trends in Microbiology 7: 392-401. ARUMUGHAM RG, HSIEH TC, TANZER ML & LAINE RA. 1986. Structures of the asparagine-linked sugar chains of laminin. Biochimica et Biophysica Acta 883:112-26. ARRIGHI RB & HURD H. 2002. The role of plasmodium berghei ookinete proteins in binding to basal lamina components and transformation into oocyts. International Journal of Parasitology 32: 91-98.

76
ASSREUY J, CUNHA FQ, EPPERLEIN M, NORONHA-DUTRA A, O'DONNELL CA, LIEW FY & MONCADA S. 1994 Production of nitric oxide and superoxide by activated macrophages and killing of Leishmania major. Europe Journal of Immunology 24: 672-676. AUMAILLEY M, WIEDEMANN H, MANN K & TIMPL R. 1989. Binding of nidogen and the laminin-nidogen complex to basement membrane collagen type IV. European Journal Biochemistry 184: 241-248. AWASTHI A, MATHUR KR & SAHA B. 2004. Immune response to Leishmania infection. Indian Journal of Medical Research 119: 238-258. BABAL P, & RUSSEL LC. 1999. Sialic acid-specific lectin-mediated adhesion of Tritrichomonas foetus and Tritrichomonas mobilensis. Journal of Parasitology 85: 33-40. BANDYOPADHYAY K, KARMAKAR S, GHOSH A & DAS PK. 2001. Role of 67kDa cell surface laminin binding protein of Leishmania donovani in pathogenesis. Journal of Biochemistry 130: 141-148. BANDYOPADHYAY K, KARMAKAR S, GHOSH A & DAS PK. 2002. High affinity binding between laminin and laminin binding protein of Leishmania is stimulated by zinc and may involve laminin zinc-finger like sequences. European Journal of Biochemistry 269: 1622-1629. BANDYOPADHYAY K, KARMAKAR S, BISWAS A & DAS PK. 2003. Membrane orientation of laminin binding protein. A extracellular matrix bridging molecule of Leishmania donovani. European Journal of Biochemistry 270: 3806-3813. BATES, P. A. 1994. The developmental biology of Leishmania promastigotes. Experimental. Parasitology 79: 215-218. BEHNSEN J, NARANG P, HASENBERG M, GUNZER F, BILITEWSKI U, KLIPPEL N, ROHDE M, BROCK M, BRAKHAGE AA & GUNZER M. 2007. Environmental dimensionality controls the interaction of phagocytes with the pathogenic fungi Aspergillus fumigatus and Candida albicans. PLoS Pathogens 3: 13. BELLEY, A. & CHADEE, K. 1995. Eicosanoids production by parasites: from pathogenesis to immunomodulation. Parasitology Today 11: 327-334. BENCHIMOL, M. & DE SOUZA, W. 1980. Freeze-fracture study of the plasma membrane of Leishmania mexicana amazonensis. Journal of Parasitology 66: 941-947. BENCHIMOL M, BATISTA C & DE SOUZA W. 1990Fibronectin- and laminin-mediated endocytic activity in the parasitic protozoa Trichomonas vaginalis and Tritrichomonas foetus. Journal of Submicroscopy Cytology Pathology 22: 39-45. BENCHIMOL M & ENGELKE F. 2003. Hydrogenosome behavior during the cell cycle in Tritrichomonas foetus. Biology of the Cell 95: 283-93. BERMAN JD. 1997. Human leishmaniasis: clinical, diagnostic, and chemotherapeutic developments in the last 10 years. Clinical Infection Disease 24: 684-703. BISSEL MJ & BARCELLOS –HOFF MH. 1987. The influence of extracellular matrix on gene expression: is structure the message? Journal of Cell Science Supplement 8: 327-343.

77
BOHNSACK JF, KLEINMAN HK, TAKAHASHI T, O´SHEA JJ & BROWN EJ. 1985. Connective tissue and phagocytic cell function. Journal of Experimental Medicine 161: 912-923.
BONILHA VL, CIAVAGLIA MDO C, DE SOUZA W & COSTA E SILVA FILHO F. 1995 involvement of terminal carbohydrates of the mammalian cell surface in the cytoadhesion of trichomonads. Parasitology Research 81: 121-126. BOUDREAU N, MYERS C & BISSELL MJ. 1995. From laminin to lamin: regulation of tissue-specific gene expression by the ECM. Trends in Cell Biology. 5: 1-4. BRADFORD MM. 1976. A rapid and sensitive method for the quantification of microgram quantities of proteins utilizingthe principle of protein-dye binding. Annals of Biochemistry 72: 248-254. BUATES, S. & MATLASHEWSKI, G. 2001. General suppression of macrophage gene expression during Leishmania donovani infection. The Journal of Immunology 166: 3416-3422.
BUCHMULLER-ROUILLER Y & MAUEL J. 1987. Impairment of the oxidative metabolism of mouse peritoneal macrophages by intracellular Leishmania spp. Infection and Immunity 55: 587-593. BURCHMORE, R.J.S. & BARRET, M.P. 2001. Life in vacuoles – nutrient acquisition by Leishmania amastigotes. International Journal of Parasitology 31: 1311-1320. BURGESON RE, CHIQUET M, DEUTZMANN R, EKBLOM P, ENGEL J, KLEINMAN H, MARTIN GR, MENEGUZZI G, PAULSSON M, SANES J, TIMPL R, TRYGGVASON K, YAMADA Y & YURCHENCO P. 1994. A New Nomenclature for the Laminins. Matrix Biology 14: 209-211. BURGESS DE, KNOBLOCK KF, DAUGHERTY T & ROBERTSON NP. 1990. Cytotoxic and hemolytic effects of Tritrichomonas foetus on mammalian cells. Infection and Immunity 58: 3627-32. BURGESS DE & MCDONALD CM. 1992. Analysis of adhesion and cytotoxicity of Tritrichomonas foetus to mammalian cells by use of monoclonal antibodies. Infection and Immunity 60: 4253-4259. CAVALIER-SMITH T. 1993. Kigdom protozoa and its 18 phyla. Microbiological Reviews 57: 953-994. CHANG, K.P. 1983. Cellular and molecular mechanisms of intracellular symbiosis in leishmaniasis. International Review of Cytology 14: 267-305. CHANG, K-P., REED, S.G., MCGWIRE, B.S. & SOONG, L. 2003. Leishmania model for microbial virulence: the relevance of parasite multiplication and pathoantigenicity. Acta Tropica 85: 375-390. CHAUDHURI G & CHANG KP. 1988. Acid protease activity of a major surface membrane glycoprotein (gp63) from Leishmania mexicana promastigotes. Molecular and Biochemical Parasitology 27: 43-52. CHUNG AE, JAFFE R, FREEMAN, IL, VERGNES JP, BRAGINSKI JE & CARLIN B. 1979. Properties of a basement membrane-related glycoprotein synthesized in culture by a mouse embryonal carcinoma-derived cell line. Cell 16: 277-287.

78
COBO ER, CANO D, ROSSETTI O & CAMPERO CM. 2002. Heifers immunized with whole-cell and membrane vaccines against Tritrichomonas foetus and naturally challenged with an infected bull. Veterinary Parasitology 109: 169-184. CONNARIS S & GREENWELL P. 1997. Glycosidases in mucin-dwelling protozoans. Glycoconjugate Journal 14: 879-882. COOMBS GH & MULLER M. 1995. Energy metabolism in anaerobic protozoans. In Biochemistry and Molecular Biology of Parasites. (Marr JJ & Muller M eds) p33-48. Academic Press, New York, N.Y. COOPER DNW, MASSA SM & BARONDES SH. 1991. Endogenous Muscle lectin inhibits myoblast adhesion to laminin. The Journal of Cell Biology 115: 1437-1448. CORBEIL LB, HODGSON JL, JONES DW, CORBEILRR, WIDDERS PR & STEPHENS LR. 1989. Adherence of Tritrichomonas foetus to bovine vaginal epithelial cells. Infection and immunity 57: 2158-2165. CORBEIL LB. 1994. Vaccination strategies against Tritrichomonas foetus. Parasitology Today 10: 103-106. COSTA AA, ABREU AL, GOMES CM, SALDANHA AC, BARRAL A, COSTA JM & CORBETT CE. 2006. Experimental model of chronic osteomyelitis caused by Leishmania (L) amazonensis. Acta Tropica 98: 125-129. COURRET, N., FRÉHEL, C., GOUHIER, N., POUCHELET, M., PRINA, E., ROUX, P. & ANTOINE, JC. 2002. Biogenesis of Leishmania-harbouring parasitophorous vacuoles following phagocytosis of the metacyclic promastigote or amastigota stages of the parasites. Journal of Cell Scienci. 115: 2303-2316. CROFT SL & YARDLEY V. 2002. Chemotherapy of leishmaniasis. Current Pharmaceutical Desing 8:319-342. CROUCH, ML & ALDERETE, JF. 1999. Trichomonas vaginalis interaction if fibronectin and LMN. Microbiology 145: 2834-2843. CUNNINGHAM, A.C. 2002. Parasitic adaptive mechanisms in infection by Leishmania. Experimental and Molecular Pathology 72: 132-141. DA GAMA LM, RIBEIRO-GOMES FL, GUIMARAES U JR & ARNHOLDT AC. 2004. Reduction in adhesiveness to extracellular matrix components, modulation of adhesion molecules and in vivo migration of murine macrophages infected with Toxoplasma gondii. Microbes Infection 6: 1287-1296. DAVIES CE, COOPER AM, PEACOCK C, LANE RP & BLACKWELL JM. 1990. Expression of LPG and gp63 by different development stages of Leishmania major in the sandfly, Phlebotomus papatasi. Parasitology, 101: 337-343. DEDET JP, RYTER A, VOGT E, HOSLI P & PEREIRA DA SILVA L. 1983. Uptake and killing of Leishmania mexicana amazonensis amastigotes by human skin fibroblasts. Annals of Tropical Medicine and Parasitology 77: 35-44.

79
DESCOTEAUX, A., MATLASHEWSKI, G. & TURCO, S.J. 1992. Inhibition of macrophage protein kinase C-mediated protein phosphorylation by Leishmania donovani lipophosphoglycan. Journal of Immunology 149: 3008-3015. DESCOTEAUX, A. & TURCO, S. J. 1999. Glycoconjugates in Leishmania infectivity. Biochimica and Biophysica Acta 1455: 341-352. DESCOTEAUX, A. & TURCO, S.J. 2002. Functional aspects of the Leishmania donovani lipophosphoglycan during macrophage infection. Microbes Infection 4: 975-981. DESJEUX P. 2004. Leishmaniasis: current situation and new perspectives. Comparative Immunology, Microbiology and Infection Diseases 27: 305-318. DE SOUZA, W. & BENCHIMOL, M. 1984. High voltage electron microscopy of critical point dried trypanosomatids. Journal of Submicroscopic Cytology, 16: 237-242. DE SOUZA, W. 1984. Cell biology of Trypanosoma cruzi. International Review of Cytology 86: 197-283. DIAMOND LS. 1957. The estabilishment of various trichomonads of animals and men in axenic culture. Jornal of Parasitology 43: 488-490. DWYER, D. M. 1980. Isolation and partial characterization of surface membranes from Leishmania donovani promastigotes. Journal of Protozoology 27: 176-182. ELOLA TM, CHIESA EM, ALBERTI AF, MORDOH J & FINK EM. 2005. Galectin-1 receptors in different cell types. Journal of Biomedical Science 12: 13-29. ENGVALL E & WEWER M. 1996. Domains of Laminin. Journal of Cellular Biochemistry 61: 493-501. EKBLOM P, LONAI P & TALTS JF. 2003. Expression and biological role of Laminin-1. Matrix Biology 22: 35-47. FELLEISEN RSJ. 1999. Host-parasite interaction in bovine infection with Tritrichomonas foetus. Microbes and Infection 1: 807-816. FERREIRA OE, LOBO LA, PETRÓPOLIS DB, AVELAR KES, FERREIRA MC, SILVA-FILHO FC & DOMINGUES RMCP. 2006. A Bacteroides fragilis surface glycoprotein mediates the interaction between the bacterium and the extracellular matrix component laminin-1. Research in Microbiology 157: 960–966. FLOCK JI. 1999. Extracellular-matrix-binding proteins as targets for the prevention of Staphylococcus aureus infection. Molecular Medicine Today 5: 532-537. FLORES-ROBLES D, ROSALES C, ROSALES-ENCINA JL & TALAMAS-ROHANA P. 2003. Entamoeba histolytica: a beta 1 integrin-like fibronectin receptor assembles a signaling complex similar to those of mammalian cells. Experimental Parasitology 103:8-15. FRANCO E, MANNING-CELA R & MEZA I. 2002. Signal transduction in Entamoeba histolytica induced by interaction with fibronectin: presence and activation of phosphokinase A and its possible relation to invasiveness. Archives of Medical Research 33: 389-397.

80
FREIRE E & COELHO-SAMPAIO T. 2000. Self-assembly of Laminin Induced by Acidic pH. The Journal of Biological Chemistry 275: 817-822. FUJIWARA S, SHINKAI H, DEUTZMANN R, PAULSSON M & TIMPL R. 1988. Structure and distribution of N-linked oligosaccharide chains on various domains of mouse tumour laminin. Biochemistry Journal 252: 453-461. GALÁN J.E., PACE J. & HAYMAN M.J. 1992. Involvement of the epidermal growth factor receptor in the mammalian cells by salmonella typhimurium. Nature 257: 588-589 GEE SH, BLACHER RW, DOUVILLE PJ, PROVOST PR, YURCHENCO PD & CARBONETTO S.1993. Laminin-binding protein 120 from brain is closely related to the dystrophin-associated glycoprotein, dystroglycan, and binds with high affinity to the major heparin binding domain of laminin. Journal of Biological Chemistry 268: 14972-14980. GHOSH A, KOLE L, BANDYOPADHYAY K, SARKAR K & DAS PK. 1996. Evidence of a laminin binding protein on the surface of Leishmania donovani. Biochemical and Biophysical Research Communications 226: 101-106. GHOSH A, BANDYOPADHYAY K, KOLE L & DAS PK. 1999. Isolation of a laminin binding protein from the protozoan parasite Leishmania donovani that may mediate cell adhesion. The Biochemical Journal 337: 551-558. GRANGER BL & WARWOOD SJ. 1996. Rapid internalization and degradation of surface-bound antibodies by Tritrichomonas foetus. Journal of Parasitology 82: 539-549. GRANGER BL, WARWOOD SJ, BENCHIMOL M, & DE SOUZA W. 2000. Transient invagination of flagella by Tritrichomonas foetus. Parasitology Research 89: 699-709. GREVELINK SA & LERNER EA. 1996 Leishmaniasis. Journal of America Academic Dermatology 34: 257-272. GRIMBLEBY FH & NTAILIANAS HA. 1961. Binding of trichloroacetic acid by protein. Nature 189: 835-836. GRINNELL F. 2003. Fibroblast biology in three-dimensional collagen matrices. Trends in Cell Biology 13: 264-269. GORDON VR, ASEM EK, VODKIN MH & MCLAUGHLIN GL. 1993. Acanthamoeba binds to extracellular matrix proteins in vitro. Investigative Ophthamology & Visual Science 34: 658-662. GU M, WANG W, SONG WK, COOPER DNM & KAUFMAN SJ. 1994. Selective modulation of the interaction of α7β1 integrin with fibronectin and laminin by L-14 lectin during skeletal muscle differentiation. Journal of Cell Science 107: 175-181. GU Y, KORTESMAA J, TRYGGVASON K, PERSSON J, EKBLOM P, JACOBSEN SE & EKBOLM M. 2003. Laminin isoform-specific promotion of adhesion and migration of human bone marrow progenitors cells. Blood 101: 877-885.

81
GUY, R.A. & BELOSEVIC, M. 1993. Comparison of receptors required for entry of Leishmania major amastigotes into macrophages. Infection and Immunity 61: 1553-1558. HANDMAN, E. & BULLEN, D. V. R. 2002. Interaction of Leishmania with the host macrophage. Trends in Parasitology 18: 332-334. HANSEN U, HUSSAIN M, VILLONE D, HERRMANN M, ROBENEK H, PETERS G, SINHA B & BRUCKNER P.2006. The anchorless adhesin Eap (extracellular adherence protein) from Staphylococcus aureus selectively recognizes extracellular matrix aggregates but binds promiscuously to monomeric matrix macromolecules. Matrix Biology 25: 252-260. Hart DT, Vickerman K & Coombs GH. 1981. A quick, simple method for purifying Leishmania mexicana amastigotes in large numbers. Parasitology 82: 345-355. He YW, Li H, Zhang J, Hsu CL, Lin E, Zhang N, Guo J, FORBUSH KA & BEVAN MJ. 2004. The extracellular matrix protein mindin is a pattern-recognition molecule for microbial pathogens. Nature Immunology 5: 88-97. HECK LW, MORIHARA K & ABRAHAMSON DR.1986. Degradation of soluble laminin and depletion of tissue-associated basement membrane laminin by Pseudomonas aeruginosa elastase and alkaline protease. Infection and Immunity 54: 149-153. HERNÁNDES-RAMÍREZ VI, ANAYA-RUIZ M, RIOS A & TALÁMAS-ROHANA P. 2000. Entamoeba histolytica: Tyrosine Kinase Activity Induces by Fibronectin through the beta 1- integrin-like Molecule. Experimental Parasitology 95: 85-95. HODGE-DUFOUR J, NOBLE P W, HORTON MR, BAO C, WYSOKA M, BURDICK MD, STRIETER RM, TRINCHIERI G, & PURE E. 1997. Induction of IL-12 and chemokines by hyaluronan requires adhesion-dependent priming of resident but not elicited macrophages. The Journal of Immunology 159: 2492–2500. HODGSON JL, JONES DW, WIDDERS PR & CORBEIL LB. 1990 Characterization of Tritrichomonas foetus antigens by use of monoclonal antibodies. Infection Immunity 58: 3078-3083. HONIGBERG BM. 1978. Trichomonads of veterinary importance. In Parasitic Protozoa. (Kreier JP ed), pp. 164-273. Academic Press, New York, N.Y. HORIE M, MURAKAMI T, SATO T, TARUSAWA Y, NAKAMURA S & TOBA T. 2005. Anaerobic induction of adherence to laminin in Lactobacillus gasseri strains by contact with solid surface. Current Microbiology 51: 275-82. HORWITZ FA. 1997. Integrins and Health. Scientific American 276: 68-75. HUARD TK, MALINOFF HL & WICHA MS. 1986. Macrophages express a plasma membrane receptor for basement membrane laminin. The American Journal of Pathology 123: 365-370. HUNT, R. C. & ELLAR, D. J. 1974. Isolation of the plasma membrane of a trypanosomatid flagellate: general characterization and lipid composition. Biochimica et Biophysica Acta 339: 173-189. ILG, T. 2000. Proteophosphoglycans of Leishmania. Parasitology Today 16: 489-497.

82
JONES JK, MORGAN G, JOHNSTON H, TOBIAS V, OUVRIER RA, WILKINSON I & NORTH KN. 2001. The expanding phenotype of laminin α2 chain (merosin) abnormalities: case series and review. Journal of Medical Genetics 38: 649-657. KALLURI R. 2003. Basement Membranes: structure, assembly and role in tumour angiogenesis. Nature 3: 422- 433. KARP CLSH, EL-SAFI TA, WYNN MM, SATTI AM, KORDOFANI F A, HASHIM M, HAG-ALI FA, NEVA TB, NUTMAN & D. L. SACKS. 1993. In vivo cytokine profiles in patients with kala-azar: marked elevation of both interleukin- 10 and interferon. Journal of Clinical Investigation 91: 1644–1648. KENNETT MJ & HOOK RR. 2002. Tritrichomonas foetus: characterization of isolates and partial purification of a secreted cytotoxin. Experimental Parasitology 102: 1-8. KIKKAWA Y, YO H, GENERSCH E, SANZEN N, SEKIGUSH K, FASSLER R, CAPPELL K & TALTS J. 2004. Laminin isoforms differentially regulate adhesion, spreading,proliferation, and ERK activation of h1 integrin-null cells. Experimental Cell Research 300: 94-108. KIRBY H. 1951 Observations on the trichomonad flagellate of the reproductive organs of cattle. Journal of Parasitology 37: 445-459. KOFOID CA & SWEZY O. 1915. Mitosis in trichomonas. Proceedings of the national Academy of Sciences USA 1: 315-321. KOSHIKAWA N, MORIYAMA K, TAKAMURA H, MIZUSHIMA H, NAGASHIMA Y, YANOMA S & MIYAZAKI K. 1999. Overexpression of laminin gamma2 chain monomer in invading gastric carcinoma cells. Cancer Research 59: 5596-5601. KRONVALL G & JONSSON K. 1999. Receptins: a novel term for an expanding spectrum of natural and engineered microbial proteins with binding properties for mammalian proteins. Journal of Molecular Recognition 12: 38-44. LAEMMLI U.K. 1970. Cleavege of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. LAINSON, R. & SHAW, J. J. 1972. Taxonomy of the new world Leishmania species. Transactions of the Royal Society of Tropical Medicine and Hygiene 66: 943-944. LAL K, NOEL CJ, FIELD MC, GOULDING D & HIRT RP. 2006. Dramatic reorganisation of Trichomonas endomembranes during amoebal transformation: a possible role for G-proteins. Molecular and Biochemical Parasitology 148: 99-102. LAUWAET T, OLIVEIRA MJ, MAREEL M & LEROYA. 2000. Molecular mechanisms of invasion by câncer cells, leukocytes and microorganisms. Microbes and Infection 2: 923-931. LI E, YANG WG, ZHANG T & STANLEY JR SL. 1995. Interaction of laminin with Entamoeba histolytica cysteine proteinases and its effects on amebic pathogenesis. Infection and immunity 63: 4150-4153. LINDER JC. & STAEHELIN LA. 1977. Plasma membrane specializations in a trypanosomatid flagellate. Journal of Ultrastructure Research 60: 246-262.

83
LINDMARK DG & MULLER M. 1973. Hydrogenossome, a cytoplasmic organelle of the anaerobic flagellate Tritrichomonas foetus. The Journal of Biological Chemistry 248: 7724-7728. LIRA R, ROSALES-ENCINA JL & ARGUELLOS C. 1997. Leishmania mexicana: Binding of promastigotes to type I collagen. Experimental Parasitology 85: 149-157. LOCKWOOD BC, NORTH MJ & COOMBS GH. 1984. Trichomonas vaginalis, Tritrichomonas foetus and Trichomitus batrachorum: comparative proteolytic activity. Experimental Parasitology 58: 245-253. LOVE, D.C., ESKO, J.D. & MOSSER, D.M. 1993. A heparin-binding activity on Leishmania amastigotes which mediates adhesion to cellular proteoglycans. Journal of Cell Biology 123: 759-766. MACK SR & MULLER M. 1978. Effect of oxygen and carbon dioxide on the growth of Trichomonas vaginalis and Tritrichomonas foetus. Journal of Parasitology 64: 927-929. MASOCHA W, ROBERTSON B, ROTTENBERG ME, MHLANGA J, SOROKIN L & KRISTENSSON K. 2004. Cerebral vessel laminins and IFN-gamma define Trypanosoma brucei brucei penetration of the blood-brain barrier. Journal of Clinical Investigation 114: 689-694. MATTE, C., MAION, G., MOURAD, W. & OLIVIER, M. 1999. Leishmania donovani-induced macrophages cyclooxygenase-2 and prostaglandin E2 synthesis. European Journal of Immunology 29: 3737-3744. MAUËL, J. 1996. Intracellular survival of protozoan parasites with especial reference to Leishmania spp., Toxoplasma gondii, and Trypanosoma cruzi. Advences in Parasitology 38: 1-51. MCDONALD C & NUÑEZ G. 2004. Midin the fort. Nature Immunology 5:16-18. MCDOWELL, M.A. & SACKS, D.L. 1999. Inhibition of host cell signal transduction by Leishmania: observations relevant to the selective impairment of IL-12 responses. Current Opinion in Microbiology 2: 438-443. MCGWIRE BS, CHANG KP & ENGMAN DM. 2003. Migration through the extracellular matrix by the parasitic protozoan Leishmania is enhanced by surface metalloprotease gp63. Infection and Immunity 71: 1008-1010. MEDZHITOV R, PRESTON-HURLBURT P & JANEWAY CA. 1997. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 6640: 394-397. MENDONZA-LEÓN A., SHAW J.J. & TAPIA F.J. 1996. A guide for the cutaneous leishmaniasis connoisseur. In: Molecular and immune mechanisms in the pathogenesis of cutaneous leishmaniasis. 1st edition, Tapia F.J., Cáceres-Dittmar G. and Sánchez M.A. Eds. Chapman & Hall, New York. MELO-BRAGA MB, ROCHA-AZEVEDO B & SILVA-FILHO FC. 2003. Tritrichomonas foetus the role played by iron during parasite interaction with epithelial cells. Experimental Parasitology 105: 111-120. MERCURIO AM. 1995. Laminin receptors: achieving specificity through cooperation. Trends in Cell Biology 5: 419-423.

84
MOISEEVA EP, WILLIAMS B & SAMANI NJ. 2003. Galectin-1 inhibits incorporation of vitronectin and chondroitin sulfate B into the extracellular matrix of human vascular smooth muscle cells. Biochimica et Biophysica Acta 1619: 125-132. MOORE, K.J. & MATLASHEWSKI, G. 1994. Intracellular infection by Leishmania donovani inhibits macrophage apoptosis. The Journal of Immunology 152: 2930-2937. MORWOOD SR & NICHOLSON LB. 2006. Modulation of the immune response by extracellular matrix proteins. Archives of Immunological Therapy Experimental 54: 367-74. MUHLE C, JIANG QJ, CHARLESWORTH A, BRUCKNER-TUDERMAN L, MENEGUZZI G & SCHNEIDER H. 2005. Novel and recurrent mutationsin the laminin-5 genes causing lethal junctional epidermolysis bullosa: molecular basis and clinical course of Herlitz disease. Human Genetic 116: 33-42. NDE PN, SIMMONS KJ, KLESHCHENKO YY, PRATAP S, LIMA MF, VILLALTA F. 2006 . Silencing of the laminin gamma-1 gene blocks Trypanosoma cruzi infection. Infection and Immunity 74: 1643-1648. NIESSEN CM, HOGERVORST F, JASPARS LH, DE MELKER AA, DELWEL GO, HULSMAN EH, KUIKMAN I & SONNENBERG A.1994. The alpha 6 beta 4 integrin is a receptor for both laminin and kalinin. Experimental Cell Research 211: 360-367. NISHIUCHI R NISHIUCHI R, TAKAGI J, HAYASHI M, IDO H, YAGI Y, SANZEN N, TSUJI T, YAMADA M & SEKIGUCHI K. 2006. Ligand-binding specificities of laminin-binding integrins: a comprehensive survey of laminin-integrin interactions using recombinant alpha3beta1, alpha6beta1, alpha7beta1 and alpha6beta4 integrins. Matrix Biology 25:189-197. NOMIZU M, KIM WH, YAMAMURA K, UTANI A, OTAKA SYA, ROLLER PP, KLEINMAN HK & YAMADA Y. 1995. Identification of cell binding sites in the laminin α1 chain carboxyl-terminal globular domain by systematic screening of synthetic peptides. The Journal of Biological Chemistry 270: 20583-20590. NOMIZU M, KURATOMI Y, PONCE ML, SONG SY, MIYOSHI K, OTAKA A, POWELL SK, HOFFMAN MP, KLEINMAN HK & YAMADA Y. 1997. Identification of cell binding sequences in mouse laminin 1 chain by systematic peptides screening. The Journal of Biological Chemistry 272: 32198-205. NOMIZU M, KURATOMI Y, MALINDA KM, SONG SY, MIYOSHI K, OTAKA A, POWELL SK, HOFFMAN MP, KLEINMAN HK & YAMADA Y. 1998. Cell binding sequences in mouse laminin α1 chain. The Journal of βiological Chemistry. 273: 32491-32499. NOMIZU M, KURATOMI Y, PONCE ML, SONG SY, MIYOSHI K, OTAKA A, POWELL SK, HOFFMAN MP, KLEINMAN HK & YAMADA Y. 2000. Cell adhesive sequences in mouse laminin β1 chain. Archives of Biochemistry and Biophysics. 378: 311-320. NOMIZU M, YOKOYAMA F, SUZUKI N, OKAZAKI I, NISHI N, PONCE ML, KLEINMAN HK, YAMAMOTO Y, NAKAGAWA S & MAYUMI T. 2001. Identification of Homologous biologically active sites on the N-terminal domain of laminin alpha chains. Biochemistry 40: 15310-15317. OLIVIER, M., BAIMBRIDGE, K. G. & REINER, N.E. 1992. Stimulus-response coupling in monocytes infected with Leishmania. Attenuation of calcium transients is related to defective agonist-induced accumulation of inositol phosphates. Journal of Immunology 148: 1188-1196.

85
OUAISSI MA, CORNETTE J & CAPRON A. 1986. Identification and isolation of Trypanosoma cruzi trypomastigote cell surface protein with properties expected of a fibronectin receptor. Molecular and Biochemical Parasitology 19: 201-211. PARSONSON IM, CLARK BL & DUFTY J. 1974. The pathogenesis of Tritrichomonas foetus infection in the bull. Australian Veterinary Journal 50: 421-423. PARSONSON IM, CLARK BL & DUFTY J. 1976. Early pathogenesis and pathology of Tritrichomonas foetus infection in virgin heifers. Journal of Comparative Pathology 86: 59-66. PATARROYO M, TRYGGVASON K & VIRTANEN I. 2002. Laminin isoforms in tumor invasión, angiogenesis and metastasis. Cancer Biology 12: 197-207. PETERS, C., AEBISCHER, T., STIERHOF, Y-D., FUCHS, M. & OVERATH, P. 1995. The role of macrophage receptors in adhesion and uptake of Leishmania mexicana amastigotes. Journal of Cell Science 108: 3715-3724. PETERS, C., STIERHOF, Y-D. & ILG, T. 1997. Proteophosphoglycan secreted by Leishmania mexicana amastigotes causes vacuole formation in macrophages. Infection and Immunity 65: 783-786. PETERSON KM & ALDERETE JF. 1982 Host plasma proteins on the surface of pathogenic Trichomonas vaginalis. Infection and Immunity 37: 755-762. PETRIN D, DELGATY K, BHATT R & GARBER G. 1998. Clinical and microbiological aspects of Trichomonas vaginalis. Clinical Microbiology Review 11: 300-317. PETROPOLIS DB, ROCHA-AZEVEDO B MENEZES GC & SILVA-FILHO FC. 2007. The conversation between ECM molecules and parasitic protozoa. Trends in Parasitology. Aceito para publicação. PIMENTA, P. F. P & DE SOUZA. 1983. Leishmania Mexicana amazonensis: surface charge of amasigotas and promastigotas forms. Experimental Parasitology 56: 194-206. PIMENTA, P. F. P., DA SILVA, R. P. & SACKS, D. L. 1989. Cell surface nanoanatomy of Leishmania major as revealed by fracture lip. A surface meshwork of 44 nm fusiform filaments identifies infective developmental stage promastigotes. European Journal of Cell Biology 48: 180-190. PIMENTA, P. F. P., SARAIVA, E. M. B. & SACKS, D. L. 1991. The fine structure and surface glycoconjugate expression of three life stages of Leishmania major. Experimental Parasitology 72: 191-204. PLOW EF, HAAS TA, ZHANG L, LOFTUS J & SMITH JW. 2000. Ligand binding to integrins. The Journal of Biological Chemistry 275: 221785-221788. PRADEL G, GARAPATY S & FREVERT U. 2002. Proteoglycans mediate malaria sporozoite targeting to the liver. Molecular Microbiology 45: 637-651 PROUNDFOOT, L., O’DONELL, C.C. & LIEW, F.Y. 1995. Glycoinositolphospholipids of Leishmania major inhibit nitric oxide synthesis and reduce leishmanicidal activity in murine macrophages. European Journal of Immunology 25: 745-750.

86
PUPKIS MF, TETLEY L & COOMBS GH. 1986 Leishmania mexicana: amastigote hydrolases in unusual lysosomes. Experimental Parasitology 62: 29-39. RABINOVITCH M, ZILBERFARB V & RAMAZEILLES C. 1986. Destruction of Leishmania Mexicana amazonensis amastigotes within macrophages by lysosomotropic amino acid esters. Journal of Experimental Medicine 163: 520-535. RAE DO, CHENOWETH PJ, GENHO PC, MCINTOSH AD, CROSBY E & MOORE SA. 1999. Prevalence of Tritrichomonas foetus in a bull population and effect on production in a large cow-calf enterprise. Journal of American Veterinary medicine Association 214: 1051-1055. RAO SP, GEHLSEN KR & CATANZARO A. 1992. Identification of a β1 integrin on Mycobacterium avium- Mycobacterium intracellulare. Infection and Immunity 60: 3652-3657. RATH S., TRIVELIN L.A., IMBRUNITO T.R., TOMAZELA D.M., JESÚS M.N., MARZAL P.C., ANDRADE-JUNIOR H.F. & TEMPONE A.G. 2003. Antimonials employed in the treatment of leishmaniaisis: the state of the art. Quím. Nova 26: 550-555. RAY, M. GAM, A.A., BOYKINS, R.A. & KENNEY, R.T. 2000. Inhibition of interferon-γ signaling by Leishmania donovani. Journal of Infection Disease 181: 1121-1128. RIBEIRO KC, MONTEIRO-LEAL LH & BENCHIMOL M. 2000. Contributions of the axostyle and flagella to closed mitosis in the protists Tritrichomonas foetus and Trichomonas vaginalis. Journal of Eukaryotic Microbiology 47: 481-492. RIBEIRO KC, PEREIRA-NEVES A & BENCHIMOL M. 2002. The mitotic spindle and associated membranes in the closed mitosis of trichomonads. The Biology of the Cell 94: 157-172 RICHARD BL, NOMIZU M, YAMADA Y & KLEINMAN HK. 1996. Identification of synthetic peptides derived from laminin α1 and α2 chains with cell type specificity for neurite outgrowth. Experimental Cell Research 228: 98-105. RITTING, M. G. & BOGDAN, C. 2000. Leishmania-host cell interaction: complexities and alternative views. Parasitology Today 16: 292-297. ROUSSELLE P & AUMAILLEY M. 1994. Kalinin is more efficient than laminin in promoting adhesion of primary keratinocytes and some other epithelial cells and has a different requirement for integrin receptors. The Journal of Cell Biology 125: 205-14. ROUSSELLE P, GOLBIK R, VAN DER REST M & AUMAILLEY M. 1995. Structural requirement for cell adhesion to kalinin (laminin-5). Journal of Biological Chemistry 270: 13766-13770. RUDENKO G, HOHENESTER E & MULLER YA. 2001. LG/LNS domains: multiplefunctions - one business end? Trends in Biochemical Sciences 26: 363-368. RUOSLAHTI E & ENGVALL E. 1994. Extracellular matrix components. In: Methods in Enzymology 245. San Diego: Academic press.

87
SAARELA S, WESTERLUND-WIKSTROM B, RHEN M & KORHONEN TK. 1996. The GafD protein of the G (F17) fimbrial complex confers adhesiveness of Escherichia coli to laminin. Infection and Immunity 64: 2857-2860. SAHASRABUDDHE AA, BAJPAI VK & GUPTA CM. 2004. A novel form of actin in Leishmania: molecular characterisation, subcellular localisation and association with subpellicular microtubules. Molecular and Biochemical Parasitology. 134: 105-14. SARAVIA N.G., WEIGLE K., SEGURA O.I., GIANNINI S.H., PACHECO R., LABRADA L.A. & GONÇALVES A. 1990. Recurrent lesions in human Leishmania braziliensis infection--reactivation or reinfection? Lancet 336: 398-402.
SASAKI T & TIMPL R. 2001. Domain IVa of laminin alpha5 chain is cell-adhesive and binds beta1 and alphaVbeta3 integrins through Arg-Gly-Asp. FEBS Letters 509: 181-185. SASAKI T, FASSLER R & HOHENESTER E. 2004. Laminin: the crux of basement membrane assembly. The Journal of Cell Biology. 164: 959-963 SCHENK S & QUARANTA V. 2003. Tales from the crypt[ic] sites of the extracellular matrix. Trends in Cell Biology 7: 366-375. SCHULZE B, MANN K, POSCHL E, YAMADA Y & TIMPL R. 1996. Structural and functional analysis of the globular domain IVa of the alpha 1 chain andits impact on an adjacent RGD site. Journal of Biochemistry. 314: 847-851. SENGUPTA K, HERNANDEZ-RAMIREZ VI, RIOS A, MONDRAGON R & TALAMAS-ROHANA P. 2001. Entamoeba histolytica: monoclonal antibody against the beta1 integrin-like molecule (140 kDa) inhibits cell adhesion to extracellular matrix components. Experimental Parasitology 98: 83-89. SHAIA CI, VOYICH J, GILLIS SJ, SINGH BN & BURGESS DE. 1998. Purification and expression of the Tf190 adhesin in Tritrichomonas foetus. Infection and Immunity 66: 1100-1105. SHAW LM & MERCURIO AM.1994. Regulation of cellular interactions with laminin by integrin cytoplasmic domains: the A and B structural variants of the alpha 6 beta 1 integrin differentially modulate the adhesive strength, morphology, and migration of macrophages. Molecular Biology of the Cell 5: 679-690. SHUR DB. 1993. Glycosyltransferases as cell adhesion molecules. Current Opinion in Cell Biology 5: 854-563. SILER U, SEIFFERT M, PUCH S, RICHARDS A, TOROK-STORB B, MÜLLER CA, SOROKIN L & KLEIN G. 2000. Characterization and functional analysis of laminin isoforms in human bone marrow. Blood 96: 4194-4203. SILVA AA, ROFFÊ E & LANNES-VIEIRA J. 1999. Expression of extracellular matrix components and their receptors in the central nervous system during experimental Toxoplasma gondii and Trypanosoma cruzi infection. Brazilian Journal of Medical Biological Research 32: 593-600. SILVA FILHO F, ELIAS CA & DE SOUZA W. 1982 The surface charge of Tritrichomonas foetus. Journal of Protozoology 29: 551-555.

88
SILVA-FILHO FC, ELIAS CA & DE SOUZA W. 1987. Role of divalent cátions, pH, cytoskeleton components and surface charge on the adhesion of Trichomonas vaginalis to the polystyrene substrate. Memórias do Instituto Oswaldo Cruz 82: 379-384. SILVA-FILHO FC & DE SOUZA W. 1988. The interaction of Trichomonas vaginalis and Tritrichomonas foetus with epithelial cells in vitro. Cell Structure and Function 13: 301-310. SILVA-FILHO FC, DE SOUZA W & LOPES JD. 1988. Presence of laminin-biding proteins in trichomonads and their role in adhesion. Proceedings of the national Academy of Sciences USA 85: 8042-8046. SILVA-FILHO FC, ORTEGA-LOPES J & ARROYO R. 1998. YIGSR is the preferential laminin-1 residing adhesion sequence for Trichomonas vaginalis. Experimental Parasitology. 88: 240-242. SILVA-FILHO FC. 1999. Integrins in the Trichomonas vaginalis. Memórias do instituto Oswaldo Cruz 94 (Suppl.II): 48. SILVA-FILHO FC, KASAI S, NOMIZU M, LÓPEZ LB, MELO-BRAGA MB, ROCHA-AZEVEDO B, PETRÓPOLIS DB & HORBACH IS. 2002. How laminin-1 can be recognized by the protozoan parasite Tritrichomonas foetus: possible role played by the extracellular matrix glycoprotein in both cytoadhesion and cytotoxicity exerted by the parasite. Parasitology International 51: 305-307. SILVEIRA FT, LAINSON R & CORBETT CE. 2005. Further observations on clinical, histopathological and immunological features of borderline disseminated cutaneous leishmaniasis caused by Leishmania (L) amazonensis. Memórias do Instituto Oswaldo Cruz 100: 525-534. SINGH BN, LUCAS JJ, BEACH DH, SHIN ST & GILBERT RO. 1999. Adhesion of Tritrichomonas foetus to bovine vaginal epithelial cells. Infection and Immunity 67: 3847-3854. SINGH BN, BONDURANT RH, CAMPERO CM & CORBEIL LB. 2001. Immunological and biochemical analysis of glycosylated surface antigens and lipophosphoglycan of Tritrichomonas foetus. Journal of Parasitology 87: 770-777. SKIRROW SZ & BONDURANT RH. 1988. Treatment of bovine trichomoniasis with ipronidazole. Australian Veterinary Journal 65: 156. SPARROW CP, LEFFLER H & BARONDES SH. 1987. Multiple soluble β-galactoside-binding lectins from human lung. The Journal of Biological Chemistry 262: 7383-7390. STRAUSS, AH., LEVERY, SB., JASIULIONIS, MG., SALYAN, MEK., STEELE, SJ., TRAVASSOS, LR., HAKOMORI, S. & TAKAHASHI, HK. 1993. Stage-specific glycosphingolipids from amastigote forms of Leishmania amazonensis. The Journal of Biological Chemistry 268: 13723-13730. TALAMÁS-ROHANA, P., WRIGHT, S.D., LENNARTZ, M.R. & RUSSEL, D.G. 1990. Lipophosphoglycan from Leishmania mexicana promastigotes binds to members of the CR3, p150,95 and LFA-1 family of leukocyte integrins. The Journal of Immunology 144: 4817-4824. TALÁMAS-ROHANA P & RIOS A. 2000. Actin Stress Fibres in Entamoeba histolytica induced by fibronectin. Archives of Medical Research 31: 131-133.

89
TALBOT JA, NIELSEN K & CORBEIL LB. 1991. Cleavage of proteins of reproductive secretions by extracellular proteinases of Tritrichomonas foetus. Canadian Journal of Microbiology 37: 384-390. TASHIRO KI, SEPHEL GC, WEEKS B, SASAKI M, MARTIN GR, KLEINMAN HK & YAMADA Y. 1989. A synthetic peptide containing the IKVAV sequences from the α chain of laminin mediates cell attachment, migration and neurite outgrowth. The Journal of Biological Chemistry 264: 16174-16182. TERRANOVA VP, ROHRBACH DH & MARTIN GR. 1980. Role of laminin in the attachment of PAM 212 (Epithelial) cells to basement membrane collagen. Cell 22: 719-726. TIMPL R, WIEDEMANN H, VAN DELDEN V, FURTHMAYR H & KUHN K. 1981. A network model for the organization of type IV collagen molecules in basement membranes. European Journal of Biochemical 120: 203-211. TIMPL R & BROWN JC. 1994. The Laminins. Matrix Biology 14: 275-278. TOWBIN H, STAEHELIN T & GORDON J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proceedings of the national Academy of Sciences USA 76: 350-354. TURCO SJ & DESCOTEAUX A. 1992. The lipophosphoglycan of Leishmania parasites. Annual Review of Microbiology 46: 65-94. VAN DER BRÛLE FA, BUICU C, BALDET M, SOBEL ME, COOPER DNW, MARSCHAL P & CASTRONOVO V. 1995. Galectin-1 modulates human melanoma cell adhesion to laminin. Biochemical and biophysical Research Communications 209: 760-767. VANNIER-SANTOS, M.A., SARAIVA, E.M.B., MARTINY, A., NEVES, A. & DE SOUZA, W. 1992. Fibronectin shedding by Leishmania may influence the parasite-macrophage interaction. European Journal Cell Biology 59: 389-397. VANNIER-SANTOS, M.A., MARTINY, A & DE SOUZA, W. 2002. Cell biology of Leishmania spp.: invading and evading. Current Pharmaceutical Design 8: 297-318. VOGEL V, THOMAS WE, CRAIG DW, KRAMMER A & BANEYX G. 2001. Structural insights into the mechanical regulation of molecular recognition sites. Trends in Biotechnology 19: 416-23. WANG L, ASEM EK & MCLAUGHLIN L. 1994. Calcium Enhances Acanthamoeba polyphaga binding to extracellular matrix proteins. Investigative Ophthalmology & Visual Science 35: 2421-2426. WOO HJ, SHAW LM, MESSIER JM & MERCURIO AM. 1990. The major non-integrin laminin binding protein of macrophages is identical to carbohydrate binding protein 35 (Mac-2). The Journal of Biological Chemistry 265: 7097-7099. WRIGHT, S. & SILVERSTEIN, S.C. 1983. Receptors for C3b and C3bi promote phagocytosis but not the release of toxic oxygen from human phagocytes. The Journal of Experimental Medicine 158: 2017-2023. WYLER DJ, SYPEK JP & MCDONALD JA. 1985. In vitro parasite-monocyte interactions in human leishmaniasis: possible role of fibronectin in parasite attachment. Infection and Immunity 49: 305-311.

90
YANG Z, MOSSER DM, & ZHANG X. 2007. Activation of the MAPK, ERK, following Leishmania amazonensis Infection of Macrophages The Journal of Immunology 178: 1077–1085. YARLETT N. 1988. Polyamine biosynthesis and inhibition in Trichomonas vaginalis. Parasitology Today 4: 357-360. YURCHENCO PD & SHAN-CHENG Y. 1993. Self –assembly and Calcium-binding sites in Laminin: A Three-arm Interaction Model. The Journal of Biological Chemistry, 268: 17286-17299. YURCHENCO PD, AMENTA PS & PATTON BL, 2004. Basement membrane assembly, stability and activities observed through a developmental lens. Matrix Biology 22: 521-538. ZANDBERGEN GV, KLINGER M, MUELLER A, DANNENBERG S, GEBERT A, SOLBACH W & LASKAY T. 2004. Cutting edge: Neutrophil granulocyte serves as a vector for Leishmania entry into macrophages. The Journal of Immunology 173: 6521-6525.

91
ANEXO

92
The binding of Tritrichomonas foetus to immobilized laminin-1 and its role the cytotoxicity exerted by the parasite. Débora Barreiros Petrópolis, Juliany Cola Fernandes Rodrigues, Bruno da Rocha Azevedo
and Fernando Costa e Silva Filho* The contents category for the paper: Pathogens and Pathogenicity / Biochemistry and Molecular Biology The running title: Binding of T. foetus to laminin-1 UFRJ – Instituto de Biofísica Carlos Chagas Filho, Laboratório de Biologia da Superfície Celular, Rio de janeiro, Brazil *Corresponding author. Address: UFRJ – Instituto de Biofísica Carlos Chagas Filho. Prédio do CCS, bloco G, Cidade Universitária, Ilha do Fundão, 21949-900, Rio de Janeiro, RJ, Brazil. E-mail: [email protected]

2

3
Abstract
The extracellular parasitic protozoan Tritrichomonas foetus is the causative agent of bovine
trichomonosis, an urogenital disease of veterinary relevance. Here, we tabe in account tha
the host milieu may determine parasite pathogenicity. Since the extracellular matrix
recognition by pathogens is a phenomenon directly related to infections processes, we did
explore further the interaction between T. foetus and the major basal membrane
glycoprotein Laminin-1 (LMN-1). The chemical nature of the parasite molecules involved
in the attachment of T. foetus to LMN-1 and the influence of LMN-1 in the toxicity exerted
by the parasite to HeLa cells were here studied. Attachment of T. foetus to LMN-1 resulted
in remarkable morphological alterations of the parasite, became it amoeboid. T. foetus
recognized LMN-1 through specific LMN-1 sequences (AG73, C16, A208 or IKVAV and
A13) and the proteic nature of the parasite molecules involved in such recognition were
confirmed by dot blot analyses. This molecular recognition is divalent cation-dependent
and five Laminin-binding molecules (220, 200, 130, 125 and 80 kDa) were identified in T.
foetus. Previous bind of T. foetus to LMN-1 rendered the parasite more toxicic to HeLa cell
monolayers.

4
Introduction
Tritrichomonas foetus is a venereal pathogen of naturally bred cattle in which the parasite
may cause infertility and abortion (Bondurant, 2005). The pathology, commonly referred as
bovine trichomoniasis, presents different clinical manifestations between bulls and cows
(BonDurant, 1997). In bulls the parasite can be detected at the preputial cavity until urethra,
and also in deeper parts of the urogenital tract (Parsonson et al., 1974). Once infected, bulls
may harbor T. foetus throughout their lives without exhibiting clinical symptoms of the
disease. By contrast, among cows the symptoms of the disease range from unapparent
infections to severe clinical manifestations including vaginitis, cervicitis, endometritis and
pyometra, resulting in transient infertility or fetal loss (Parsonson et al., 1976). As
previously suggested (López et al., 2000), such clinical picture indicates the possibility of
T. foetus to perform tissue invasiveness.
Trichomonads possess a diversity of surface glycoconjugates including adhesins (Corbeil
et al., 1989; Singh et al., 1999; Alderete & Garza, 1988) and extracellular matrix (ECM)-
binding molecules (Silva-Filho et al., 1986), which play important roles in the interaction
of the parasite with the host.
In mammals, the extracellular matrix (ECM) is composed by a complex assortment of
glycoproteins and proteoglycans, which in turn, not only serves as scaffolds to provide a
structural framework for tissues but also regulates cell behavior (Bissel & Barcellos-Holff,
1987).
Laminin (LMN) is a large (≈ 850kDa) ECM glycoprotein involved in relevant biological
phenomena (Ekblom et al., 2003). LMN-1 provides signaling cues that mediates important
cell functions, including cell adhesion (Arrighi & Hurd, 2002; Gosh et al., 1999; Gordon et
al., 1988; Silva-Filho et al., 2002) and invasiveness by microorganisms (Bandyopadhyay et
al., 2001; Li et al., 1995).
Biochemical studies focusing the binding of LMN to eukaryotic cells, and the chemical
nature of cell surface receptors have shown that the binding of the isoform 1 of LMN
(LMN-1) to some eukaryotic cells requires the participation of certain cationic species, as
the main class of ECM receptors called integrins (Hynes, 1982; Mercurio, 1995) that have

5
their functionalities revealed only in the presence of minimum amounts of Mn++, Ca++,
Mg++ or Ca++/Mg++ (Wewer & Engvall, 1994). In addition, it has been found
oligosaccharide-binding sites on LMN-1 (Fujiwara et al., 1988) and lectin-like laminin-
binding molecules at pathogen surfaces have been described (Ferreira et al, 2006, Saarela et
al., 1996).
Since ECM binding performed by prokaryotes and eukaryotes is a relevant event for
pathogenesis and lastly tissue invasion (Li et al., 1995; Hernandez-Ramirez et al., 2000;
Meza, 2000), the aim of this study is to characterize the binding performed by T. foetus to
LMN-1, including the identification of the parasite molecules which mediate the
phenomenon, as well as the role played by such binding in the interaction between the
parasite and mammalian cultured cells.

6
Materials and Methods
Parasites.
The K strain of T. foetus (Silva-Filho et al., 1986) was used throughout. The parasites were
cultured in modified TYM medium (Lockwood et al 1988) suplemented with 10% fetal calf
serum. Both parasite density and viability were counted in a hemotimetric chamber by
using the erythrosine-B dye exclusion test (Phillips, 1973). Only parasite suspensions (≥
106.ml-1) exhibiting viability indexes higher than 90% were used.
Laminin-1 immobilization on glass surfaces.
Glass slides which were pre-cleaned with methanol were coated or not with each one of the
solution of 5, 10, 20 or 30 µg.ml-1 LMN-1 (LMN from Engelbreth-Holm Swarm Tumour;
Sigma Chem Co., USA) (Silva-Filho et al., 1988) made in Ca++, Mg++ and Mn++ -
containing 50mM HEPES free acid, pH 6.6, for 2h at 37 oC. Just after, the unbounded
LMN-1 was aspirated and discarded.
[3H]-thymidine T. foetus labeling.
Parasites grown at 37oC for 24hours were collected by centrifugation, washed once in TYM
medium without serum, and ressuspended in complete TYM medium containing 30μCi
[3H]-thymidine (Alderete et al., 1995, Rocha-Azevedo et al., 2005). After 24 hours of
incubation the resulting radiolabelled parasites were collected by centrifugation (450g), and
washed three times with 0.01M phosphate buffered 0.145M NaCl (PBS), pH 7.2, in order
to remove unassociated radioactive tracer.
Binding to immobilized LMN-1.
[3H]-thymidine labelled parasites, which were found ressuspended in sterile HEPES 10mM,
sucrose 125mM containing or not 0.01M of each one of CaCl2, MgCl2, MnCl2, following or
not an additional incubation with 5mM EDTA, were allowed to interact with each one of
uncoated and LMN-1-coated slides for 1 hour at 37 oC. In some assays the sucrose of the
buffer as replaced by 125mM of β-Lactose or Galactose. Following interaction with LMN-
1, the unbounded parasites were aspirated and discarded whereas the parasites associated to

7
the slides were solubilized with lyses buffer (0,2% SDS on 0,2% NaOH). The radioactivity
associated to the slides was measured by liquid scintillation counting. The resulting cpm
values were converted to percent values according to Rocha Azevedo et al., 2005.
Scanning electron microscopy.
Parasites were allowed to interact to glass coverslips coated or not with 20µg.ml-1 of LMN-
1 for 60 minutes at 37oC, and then carefully washed three times with PBS following
fixation with 2.5% glutaraldehyde made in 0.1M sodium cacodylate buffer, pH 7.2. Just
after, the samples were sequentially washed with PBS, post-fixed with 1% osmium
tetroxide made in 0.1M sodium cacodylate buffer (pH 7.2), dehydrated in a crescent serie
of ethanol, critical-point-dried by using CO2, and sputter-coated with gold. The samples
were examined in a JEOL JSM-5310 scanning electron microscope. The morphological
types found among the parasites were scored by counting the following: tear-shaped or
slight rounded (no amoeboid) and spreaded (amoeboid) T. foetus, in 10 randomly chosen
fields at 1000X magnification.
Trypsin and periodate treatments.
Before interaction with LMN-1, parasites were treated or not with each one of 3 mg.ml-1
trypsin for 30 min at 37°C, or 1mM sodium periodate for 30 min at 4°C, as previously
described (Crouch & Alderete, 1999). They were then washed with PBS, and their viability
indexes were calculated before assayes.
Dot blot.
Parasites were disrupted in lysis buffer (Galán et al., 1992), and incubated overnight at 4oC
with 10% trichloracetic acid (Grimbleby and Ntailianas, 1961). The resulting proteins were
resuspended in dissolving buffer (16.5mM Tris, 0.5% SDS, 0.5% β-mercaptoethanol, pH
6.8), and quantified according to Bradford (1976). They were then treated or not with 10,
50 and 100 mg.ml-1 trypsin for 15 min at 37°C or with each one of 5, 10, 20 and 50mM
sodium periodate for 30min at 4°C, and blotted onto nitrocellulose membranes which were
incubated overnight with TBST blocking buffer (Tris buffer saline with 0,1% tween 20, 5%
Molico´s milk and 1% BSA). Just after, the membranes were sequentially loaded with 10

8
μg.ml-1 LMN-1 for 2h in Tris buffered NaCl (TBS) containing 1% BSA and 0.01M of each
one of Ca++, Mg++ or Mn++ salts, and sequentially incubated for 1 h with a polyclonal anti-
laminin-1 antibody (1:100) and a secondary one conjugated to peroxidase (1:8000) in
TBST blocking buffer.
Competition with LMN-1 related peptides.
Previously to the interaction with immobilized LMN-1, parasites were pre-incubated or not
for 30 min at 4°C with 30 µg.ml-1 of each one of the following LMN-1 related peptides (a
kind gift offered by Dr M Nomizu, Hokkaido University, Japan): AG73, C16, A208 and
A13 (Table I). In other set of experiments each one of the peptides was suspended in the
interaction medium, and the non-attached parasites were aspirated and discarded, and those
which remained associated to the LMN-1-coated slides were counted by light microscopy.
Indirect imunoblotting.
Parasites were disrupted in lysis buffer (Galán et al., 1992), and incubated overnight at 4oC
with 10% trichloracetic acid (Grimbleby and Ntailianas, 1961). Subsequently, the resulting
proteins were resuspended in dissolving buffer (16.5mM Tris, 0.5% SDS, 0.5% β-
mercaptoethanol, pH 6.8), and quantified according to Bradford (1976). Aliquots of 3mg
protein.ml-1 were diluted in sample buffer (Laemmli, 1970) to a final concentration of
1mg/ml, and assayed or stored at – 20oC. SDS-PAGE analysis was performed in 7.5%
acrylamide mini-gels by using 30µg protein per slot. Protein bands separated by SDS-
PAGE were electrotransferred to nitrocellulose membranes (Towbin et al., 1979). After
protein blotting, the membranes were stored at 4°C or immediately incubated with TBST
blocking buffer followed by sequential incubations with LMN-1, a LMN-1 goat antibody,
and an anti-goat antibody conjugated to peroxidase, as above described.
Epithelial cell culture.
HeLa cells were cultured in RPMI 1640 medium (Gibco Laboratories, USA) supplemented
with 25mM Hepes, and 10% FCS. Cells were seeded in plastic bottles, and incubated under
5% CO2 atmosphere at 37°C until confluence. Cell monolayers were disrupted with 0.2%

9
trypsin and 0.58mM EDTA in PBS. The cells were then washed twice in complete culture
medium and then poured into 96 multiwell plates (105cells/ well).
Cytotoxicity.
HeLa cell monolayers were labeled with [3H]-thymidine (4μCi / 105 cells / well) overnight,
according to Singh et al., 1999. Just after, the culture medium was aspirated, and the wells
were washed twice with warm PBS prior to the addition of T. foetus. Parasites that had
been previously washed were ressuspended in RPMI medium without serum, supplemented
or not with 20 µg.ml-1 LMN-1, and then added to the monolayers (10 parasites: 1 cell).
After 5h of cell-parasite interaction carried out at 37oC the interaction medium was
collected and the released [3H]-thymidine was evaluated by liquid scintillation counting.
The cytotoxicity indexes were expressed as percent of the control (monolayer lised by 0.2%
SDS on 0.2% NaOH solution). The serum free mediun in which HeLa cells were
maintained was collected and also used as control.

10
Results
The T. foetus binding to immobilized LMN-1.
LMN-1 was immobilized onto glass slides at each one of 5, 10, 20 and 30μg.ml1, and the T.
foetus trophozoites were allowed to interact with each one of uncoated and LMN-1-coated
slides. As can be seen in the Figure 1 most of the parasites could be found attached to the
20μg.ml1 protein-coating slides. Therefore, we did use 20μg.ml-1-coated slides throughout.
Morphological aspects of the LMN-1-bound T.foetus
T. foetus was allowed to interact with each one of uncoated and LMN-1-coated slides.
Just after few minutes of contact with immobilized LMN-1 most of the parasites began to
exhibit morphological alterations. Many amoeboid forms presenting rough surfaces and
unusual cytoplasmic projections appeared among those parasites attached to LMN-1-coated
slides (Fig. 2A, B, arrows), while the usual tear-like shape was predominant in controls
(Fig. 2C, D).
A quantitative analysis was performed by recording each one of amoeboid and non-
amoeboid forms of T. foetus before and after attachment to immobilized LMN-1. The data
presented in the Table II show that the contact with LMN-1 rendered parasites more
amoeboid (25.3 per field) than the ones found attached to LMN-1 uncoated slides (3 per
field).
The chemical nature of the parasite molecules involved in the binding to immobilized
LMN-1
Before interaction with LMN-1, parasites were treated or not with trypsin or sodium
periodate. Data presented in the figure 3 show that after 60 min of contact with LMN-1 the
trypsin-treated parasites were 52% less associated to protein-coated slides than the control.
In the other hand, previous incubation of the parasites with sodium periodate did not
significantly influence their association to the immobilized protein.
Indirect dot blot analyses identified the presence of laminin-binding molecules in T.
foetus (Fig. 4). Nevertheless, when whole cell extracts of the parasite were pre-treated with
each one of 10, 50 and 100mg.ml-1 trypsin the spots corresponding to laminin-binding

11
molecules were progressively disappearing (Fig. 4). By contrast, whole extracts of T. foetus
pre-treated with sodium periodate did not exhibited significant alterations in such labeling
(Fig. 4).
The role played by cations and sugars in the parasite binding to LMN-1
Parasites allowed to interact with LMN-1-coated slides in the presence of Ca+2, Mg+2 and
Mn+2 resulted in significant increase in parasite binding to LMN-1 (Fig. 5a). The specificity
of this cation-dependent phenomenon could be confirmed since the addition of the cation
chelator EDTA to the interaction medium resulted in a binding decrease.
Experiments were designed in order to investigate the interference of LMN-1 sugar
moieties in the attachment performed by T. foetus to immobilized LMN-1. Parasites were
allowed to interact with LMN-1 in the presence of 125mM β-lactose (Fig. 5b) or galactose
(data not show). After 1 hour of interaction we did not found a significant alteration in the
number of parasites attached to LMN-1 coated slides (Fig. 5b).
Identification of LMN-1-residing peptides recognized by T. foetus
Previously to the interaction with immobilized LMN-1, parasites were incubated or not
with each one of AG73, C16, A208, and A13 LMN-1 related peptides. Alternatively, the
same amounts of each one of these four peptides were suspended in the interaction
medium. When the peptides were suspended in the interaction medium before binding of
the parasites to immobilized LMN-1, the following parasite binding rates were obtained:
49.0%, 11.0%, 29%, and 20.3%, for AG73, C16, A208, and A13, respectively (Fig. 6).
Otherwise, when the parasites were pre-incubated at 4oC for 30 min with each one of the
peptides and then poured in contact with LMN-1-coated slides, the following parasite
binding rates were obtained: 3.7%, 12.3%, 7.3%, and 9.0%, for AG73, C16, A208, and
A13, respectively (Fig. 6).
Identification of LMN-1 binding proteins
Whole extracts of T. foetus were submitted to electrophoresis, and then blotted onto
nitrocellulose. The last which were sequentially incubated with LMN-1, a polyclonal
antibody anti-LMN-1, and a peroxidase-conjugated secondary antibody resulted in the

12
appearance of five different bands of 220, 200, 130, 125 and 80 kDa (Fig. 7). When the
incubation step with LMN-1 was omitted, the incubation of the membranes with both
primary and secondary antibodies did not result in any labeling.
The cell-parasite interaction: the role played by LMN-1
Since certain extracellular matrix glycoproteins seem to improve either the cytoadhesion
as the cytotoxicity exerted by trichomonads to mammalian cultured cells (Silva-Filho et al,
1988; Crouch & Alderete, 1999) we did decide to explore the possible role played by
LMN-1 in the cytotoxicity exerted by T. foetus to HeLa cells. Suspended parasites were
incubated or not with LMN-1 and allowed with to monolayers formed by HeLa cells
previously labeled with [3H] thymidine. We did observe an improvement in the citotoxicity
performed by T. foetus when LMN-1 were present in the interaction milieu. The
monolayers formed by HeLa cells were destroyed at high extent after incubation with T.
foetus in presence of LMN-1 (Fig. 8).

13
Discussion
Tritrichomonas foetus is an extracellular parasite able to live associated to the epithelium
lining urogenital cavities and under certain circumstances to perform tissue invasiness. In
order to be infectious, T. foetus, as well as most of the extracellular parasites, have to fit
tightly on the host tissue to overcome removal by physical forces. Therefore, it is important
for these parasites to recognize the host extracellular environment, being essential for the
infection establishment and persistence. The major component of basal membrane LMN-1
probably play a remarkable role during T. foetus binding and invasiveness of the host
tissue. As previously described (Silva-Filho et al., 1988), we also observed that after few
minutes of interaction between T. foetus and immobilized LMN-1, most of parasites were
physically associated to the glycoprotein (data not show), and the number of parasites
attached to LMN-1 was directly related to the time of such interaction process. We
observed a directly correlation between the number of parasites attached to LMN-1 and the
amount of LMN-1 immobilized onto slides (Fig 1). The highest T. foetus adhesion index
was obtained at 20μg.mL-1 LMN-1. The directly relation between T. foetus adhesion and
time or protein concentration lead us to infer that the attachment of T. foetus to LMN-1
binding on laminin is a specific process where a ligand-receptor phenomenon is involved.
A strong morphological alteration in T. foetus was observed when it was found
attached to immobilized (Fig. 2). The major occurrence of amoeboid forms among parasites
attached to the LMN-1 biofilms suggests not only a parasite attachment but beside it a
adhesion process which lead to a remodeling of both parasite surface and matrix formed by
LMN-1. Thus we considered that this drastic morphological transformation triggered the
binding of T. foetus to LMN-1 might involve the participation of LMN-1 receptors of the
parasite. Trichomonas vaginalis also alters its morphology as soon as it adhere to each one
of host cells and some extracellular matrix glycoproteins (Sckwebke & Burgess, 2004;
Crouch et al., 1999; Petrin et al., 1998). Despite the potential importance of membrane
trafficking and signaling to these clinical aspects of Trichomonas, little is still known about
the molecular mechanisms underlying such morphological alteration (Arroyo et al., 1993;
Lal et al., 2006).

14
A significant number of laminin-binding molecules have been described among
microorganisms and mammalian cells, including LBPs, laminin-binding glycoproteins,
integrins and receptins (Mercurio et al., 1995 and Ferreira et al, 2006; Kronvall et al.,
1999). Here, we identify the proteic nature of LMN-binding molecules of T. foetus, and the
dependence of divalent cations to such binding. The last demonstrated the occurrence of
divalent cations dependent LMN-1 binding proteins in T. foetus we suggested at a first
glace to be integrins. It is already known the presence of integrins and integrin-like proteins
in some parasitic protozoa (Hernández-Ramírez et al., 2000). The occurrence of integrin in
T. foetus is currently under investigation in our laboratory.
It has been found different sites in the LMN-1 polypeptide chain as possible targets
for binding of both prokaryotic and eukaryotic cells (Nomizu et al., 1995; 1997; 1998).
Inasmuch, oligosaccharides present at high extent in some parts of LMN-1 structure have
been also considered as important LMN-1 recognition sites which may have lectin-like
molecules as counterparts in microorganisms (Fujiwora et al., 1988; Ferreira et al., 2006).
Some lectin-like molecules recognize LMN through β-galactose and lactose
residues (Elola et al., 2005; Wood et al., 1990; Gu et al., 1994). Tabing this in mind we did
decide to investigate the possibility of T. foetus to recognize LMN-1 through
oligosaccharide chains present at the molecule. It is important to here claim out that T.
foetus possess surface lectins which recognize lactose and galactose-rich olygoconjugates.
The presence of β-lactose or galactose (data not show) during the interaction between T.
foetus and immobilized LMN-1 did not alter the binding of the parasite to LMN-1. It
demonstrated that the parasite did not recognize LMN-1 trough any one of its galactose or
lactose oligosaccharide residues. On the other hand, we identify the recognition of some
LMN-residing peptides by T. foetus. We tested five LMN-1 related peptides in the here
described experiment. All the LMN peptides tested include well-known binding sequences
recognized by mammalian cells and microorganisms (Nomizu, et al., 1995; 1997; 1998;
Ferreira et al., 2006). It is important to here point out that each one of the peptides includes
different α1 and γ1 LMN-1 aminoacid sequences.
We demonstrated that T. foetus is able to react with LMN-1 though each one of the
five proteins. Laminin binding proteins (LBPs) of 220, 200, 130, 125 and 80 KDa could be
detected in the extract of the parasite. The recognition of five different LMN-1 related

15
peptides by T. foetus was explored by incubation of the parasite with each one of the
peptides. Previously to the attachment to the immobilized LMN-1 the parasites were treated
or not with the cited peptides. Alternatively, we added the peptides to the interaction
medium. In both cases we did find a significant inhibition in the attachment of T. foetus to
immobilized LMN-1. It means, therefore, that the surprising results obtained in the
immunobloting assay were not art factual. Nevertheless, we still do not known if chemical
entity present at the T. foetus cell surface we are here named LBP correspond to one LBP
multiligant or five different LBPs. As such binding to LMN-1 was a Ca++ / Mg++ dependent
phenomenon we have a tendency to consider the presence of integrin at the cell surface of
T. foetus. Otherwise the number of LBPs in T. foetus might be related to number of
different cell recognition sites present in the LMN-1 molecule. YIGSR, IKVAV have been
pointed out as LMN-1 residing sequences, as used as the LNG site (Rudenko et al., 2001),
as able to be recognized by prokaryotes and eukaryotes. In addition, the occurrence of a
promiscuous surface glycoprotein of T. foetus to be involved in the recognition of LMN-1
can not be ruled out.
We demonstrated that T. foetus improves its toxicity to HeLa cells when the parasite
is facing LMN-1. The increase in this cytotoxic effect might be related with an
enhancement in cytoadhesion, since LMN-1-treated T. foetus adhered more to MDCK cells
than untreated ones (Silva-Filho et al., 1988). We were just looking for cytotoxicity since
we do not consider the cytotoxicity exerted by T. foetus as a cytoadhesion-dependent
phenomenon (Silva-Filho et al., 1986). The unspecific attachment and the specific binding
of T. foetus to LMN-1 seems to us to trigger a signaling cue that might culminated in the
upregulation of cytotoxic factors as the proteases that have been found to be released during
interaction of trichomonands with mammalian cells (Alvarez-Sánchez et al., 2000).
Data available on the ECM molecules-parasitic protozoa interactions underlie the
occurrence of a permanent exchange of signals from ECM to parasites and from parasites to
ECM (Petropolis et al., 2007). The features of such “conversation” is believed to leading
parasites to take decisions on what and how they have to do into hosts. Further work is
supported by the hypothesis that the understanding of the interaction between parasites and
host extracellular matrix may provide new strategies for treatment of parasitic diseases as
trichomoniasis.

16
Acknowledgements
The authors are grateful to Dr M Nomizu (Hokkaido University, Japan) for the kind gift of
the laminin peptides. This work has been supported by the following Brazilian agencies:
CNPq-Universal, MCT-PRONEX and FUJB-UFRJ. The experiments related in this
manuscript are in agreement with the current laws of Brazil.

17
References BonDurant, R.H. (1997). Pathogenesis, diagnosis, and management of trichomoniasis in cattle. Vet Clin North Am Food Anim Pract 13(2), 345-61. Parsonson I.M., Clark B.L. & Dufty J.H. (1974). The pathogenesis of Tritrichomonas foetus infection in the bull. Aust Vet J 50, 421-423. Parsonson I.M., Clark B.L. & Dufty J.H. (1976). Early pathogenesis and pathology of Tritrichomonas foetus infection in virgin heifers. J Comp Pathol 86(1), 59-66. Singh B.N., Lucas J.J., Beach D.H., Shin S.T. & Gilbert R.O. (1999). Adhesion of Tritrichomonas foetus to bovine vaginal epithelial cells. Infect Immun 67, 3847-54 Corbeil L.B., Hodgson J.L., Jones D.W., Corbeil R.R., Widders P.R. & Stephens L.R. (1989) Adherence of Tritrichomonas foetus to bovine vaginal epithelial cells. Infect. immu. 57, 2158-2165. Bissel M.J. & Barcellos-Hoff M.H. (1987). The influence of extracellular matrix on gene expression: is structure the message? J Cell Sci Suppl 8, 327-343. Ekblom P., Lonai P. & Talts J.F. (2003). Expression and biological role of Laminin-1. Matrix Biol 22 (1), 35-47 Sasaki T., Fassler R. & Hohenester E. (2004). Laminin: the crux of basement membrane assembly. J Cell Biol 164, 959-963
Ghosh A., Bandyopadhyay K., Kole L. & Das P.K.(1999). Isolation of a laminin binding protein from the protozoan parasite Leishmania donovani that may mediate cell adhesion. Biochem J 337, 551-558.
Silva-Filho F.C., Kasai S.,Nomizu M.,Lopez L.B.,Melo-Braga M.B., Rocha-Azevedo B.,Petropolis D.B. & Horbach I.S. (2002). How laminin-1 can be recognized by the protozoan parasite Tritrichomonas foetus: possible role played by the extracellular matrix glycoprotein in both cytoadhesion and cytotoxicity exerted by the parasite. Parasitol Int 51, 305-307.
Silva-Filho F.C., De Souza W. & Lopes J.D. (1988). Presence of laminin-biding proteins in trichomonads and their role in adhesion. Proc Natl Acad Sci USA 85, 8042-46 Casta e Silva Filho F, de Souza W, Lopes JD. (1988) Presence of laminin-binding proteins in trichomonads and their role in adhesion. Proc Natl Acad Sci USA. 85, 8042-6. Gordon VR, Asen E.K., Vodkin M.H. & McLaughlin G.L. (1993). Acanthamoeba binds to extracellular matrix proteins in vitro. Invest Ophthalmol Vis Sci 34, 658-662

18
Arrighi R.B. & Hurd H. (2002). the role of plasmodium berghei ookinete proteins in binding to basal lamina components and transformation into oocyts. Inter J Parasitol 32, 91-98 Li E., Yang W.G., Zhang T. & Stanley Jr S.L. (1995). Interaction of laminin with Entamoeba histolytica cysteine proteinases and its effects on amebic pathogenesis. Infect immun 63, 4150-53 Bandyopadhyay K., Karmakar S., Ghosh A. & Das P.K. (2001). Role of 67kDa cell surface laminin binding protein of Leishmania donovani in pathogenesis. J Biochem 130, 141-148. Mercurio A.M. (1995). Laminin receptors: achieving specificity through cooperation. Trends Cell Biol 5, 419- 423 Hynes R.O., Destree A.T. & Wagner D.D. (1982). Relationships between microfilaments, cell-substratum adhesion, and fibronectin. Cold Spring Harb Symp Quant Biol 46, 659-70 Wewer, UM & Engvall, E. (1994). Laminins. Meth Enzymol 245, 85-104 Yurchenco P.D. & Shan-Cheng Y. (1993). Self –assembly and Calcium-binding sites in Laminin: A Three-arm Interaction Model. J Biol Chem 268, 17286-99 Freire E. & Coelho-Sampaio T. (2000). Self-assembly of Laminin Induced by Acidic pH. J Biol Chem 275, 817-822 Melo-Braga M.B., da Rocha-Azevedo B. & Silva-Filho F.C. (2003). Tritrichomonas foetus the role played by iron during parasite interaction with epithelial cells. Exp Parasitol 105, 111-20. Diamond L.S. (1957). The estabilishment of various trichomonads of animals and men in axenic culture. J Parasitol 43, 488-490 Galán J.E., Pace J., Hayman M.J. (1992). Involvement of the epidermal growth factor receptor in the mammalian cells by salmonella typhimurium. Nature 257, 588-589 Rocha-Azevedo B., Melo-Braga M.B. & Silva-Filho F.C. (2005). Intra-strain clonal phenotypic variation of Tritrichomonas foetus is related to the cytotoxicity exerted by the parasite to cultured cells. Parasitol Res 95, 106–112 Alderete J.F., Lehker M.W. & Arroyo R. (1995). The mechanisms and molecules involved in cytoadherence and pathogenesis of Trichomonas vaginalis. Parasitol Today 11, 70-74 Nomizu M., Kim W.H., Yamamura K., Utani A., Otaka S.Y.A., Roller P.P., Kleinman H.K. & Yamada Y. (1995).Identification of cell binding sites in the laminin α1 chain carboxyl-terminal globular domain by systematic screening of synthetic peptides. J Biol Chem. 270: 20583-20590

19
Nomizu M., Kuratomi Y, Ponce M.L. Song S.Y., Miyoshi K., Otaka A., Powell S.K., Hoffman M.P., Kleinman H.K. & Yamada Y. (1997). Identification of cell binding sequences in mouse laminin gama 1 chain by systematic peptides screening. J Biol Chem 272, 32198-205 Nomizu M., Kuratomi Y, Malinda K.M., Song S.Y., Miyoshi K., Otaka A., Powell S.K., Hoffman M.P., Kleinman H.K. & Yamada Y. (1998). Cell binding sequences in mouse laminin α1 chain. J biol chem. 273, 32491-99 Grimbleby G F.H. & Ntailianas H.A. (1961). Binding of trichloroacetic acid by protein. Nature 11, 835-836 Bradford M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72, 248-54. Laemmli U.K. (1970). Cleavege of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685 Towbin H., Staehelin T. & Gordon J. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 76, 4350-4. Elola T.M., Chiesa E.M., Alberti A.F., Mordoh J. & Fink E.M. (2005). Galectin-1 receptors in different cell types. J Biomed Sci 12, 13-29. Gu M., Wang W., Song W.K., Cooper D.N.M. & Kaufman S.J. (1994). Selective modulation of the interaction of α7β1 integrin with fibronectin and laminin by L-14 lectin during skeletal muscle differentiation. J Cell Sci 107, 175-181. Woo H., Shaw L.M., Messier JM & Mercurio A.M. (1990). The major non-integrin laminin binding protein of macrophages is identical to carbohydrate binding protein 35 (Mac-2). J Biol Chem 265, 7097-7099. Ferreira E.O., LoboA.L., Petrópolis D.B., Avelar K.E.S., Ferreira M.C., Silva-Filho F.C., Domingues R.M.C.P. (2006). A Bacteroides fragilis surface glycoprotein mediates the interaction between the bacterium and the extracellular matrix component laminin-1. Res Microbiol 157, 960–966. Kronvall G. & Jonsson K. (1999). Receptins: a novel term for an expanding spectrum of natural and engineered microbial proteins with binding properties for mammalian proteins. J Mol Recogn 12, 38-44.

20
Sing B.N., Lucas J.J., Beach D.H., Shin S.T. & Gilbert R.O. (1999). Adhesion of Tritrichomonas foetus to bovine vaginal epithelial cells. Infect Immun 67, 3847-54 Saarela S., Wikstro B.W., Rhen M. & Korhonen T.K. (1996). The GafD Protein of the G (F17) Fimbrial Complex Confers Adhesiveness of Escherichia coli to Laminin. Infect immun 2857–60 Fujiwara S., Shinkai H., Deutzmann, R., Paulsson M. & Timpl R. (1988) Structure and distribution of N-linked oligosaccharide chains on various domains of mouse tumour laminin. Biochem. J 252, 453-461. Phillips Hi. (1973) In Dye Exclusion Tests for Cell Viability, pp 406. Edited by PF Kruse Jr. MK Patterson Jr. Academic Press, N Y Crouch M.L. & Alderete J.F.(1999). Trichomonas vaginalis interaction if fibronectin and LMN. Microbiology 145, 2834-43. Petrin D., Delgaty K., Bhatt R. & Garber G. (1998) Clinical and microbiological aspects of Trichomonas vaginalis. Clin Microbiol Rev 11, 300-17. Schwebke J.R. & Burgess D. (2004). Trichomoniasis. Clin Microbiol Rev 17, 794-803 Lal K., Noel C.J., Field M.C., Goulding D. & Hirt R.P. (2006). Dramatic reorganisation of Trichomonas endomembranes during amoebal transformation: a possible role for G-protei. Mol Biochem Parasitol 148, 99-102 Hernandez-Ramirez V.I., Anaya-Ruiz M., Rios A. & Talamas-Rohana P. (2000). Entamoeba histolytica: tyrosine kinase activity induced by fibronectin through the beta1-integrin-like molecule. Exp Parasitol 95, 85-95 Alvarez-Sanchez M.E., Avila-Gonzalez L., Becerril-Garcia C., Fattel-Facenda L.V., Ortega-Lopez J. & Arroyo R. (2000). A novel cysteine proteinase (CP65) of Trichomonas vaginalis involved in cytotoxicity. Microb Pathog 28, 193-202. Alderete J.F. & Garza G.E. (1988). Identification and properties of Trichomonas vaginalis proteins involved in cytadherence. Infect Immun. 1988 Jan;56(1):28-33. Hernandez-Ramirez V.I., Anaya-Ruiz M., Rios A. & Talamas-Rohana P. (2000). Tyrosine Kinase Activity Induces by Fibronectin through the beta 1- integrin-like Molecule. Exp Parasitol 95, 85-95. Meza I. (2000). Extracellular matrix-induced signaling in Entamoeba histolytica: its role in invasiveness. Parasitol Today 16, 23-8.

21
Lopez L.B., Braga M.B., Lopez J.O., Arroyo R. & Costa e Silva Filho F. (2000). Strategies by which some pathogenic trichomonads integrate diverse signals in the decision-making process. An Acad Bras Cienc 72, 173-86. Rudenko G, Hohenester E & Muller YA (2001). LG/LNS domais: multiple functions-one business end? Trends Biochem Sci 26, 363-68. Crouch M.L. & Alderete J.F. (1999).Trichomonas vaginalis interactions with fibronectin and laminin. Microbiology 145, 2835-43. Arroyo R., Gonzalez-Robles A., Martinez-Palomo A. & Alderete J.F. (1993). Signalling of Trichomonas vaginalis for amoeboid transformation and adhesion synthesis follows cytoadherence. Mol Microbiol 7, 299-309.

22
Tables and Figures
Table 1: LMN-1 related peptides (Nomizu et al., 1995; 1997; 1998)
Figure 1: Binding assay of [3H]-thymidine labelled T. foetus on different concentration (0, 5, 10, 20 and 30 g/ml) of Laminin-1. The number of parasites adhered was the determined by the radioactivity associated to the slides measured in a liquid scintillation counter and expressed as cpma. The bars represents the standard error.
Synthetic peptides Sequence Active sequence Localization
AG78 RKRLQVQLSIRT LQVQLSIR α chain globular domain
A13 RQVFQVAYIIIKA not determined α chain
A208 AASIKVAVSADR IKVAV α chain
C16 KAFDITYVRLKF YVRL γ chain
020406080
100120140160180
0 10 20 30 40 50
concentração de LMN (μg/ml)
cpm
a

23
Table 2: Quantification of T. foetus shape in the presence and absence of Laminin. Values represent the average of the number of parasites/ field counted by using a 1000X magnification.
No amoeboid Amoeboid
Control 22,8 ± 7,8 3 ± 1,6
Laminin 8,1 ± 3,5 25,3 ± 5,6
Control T.foetus adhered on laminin biofilm
A
B D
C
Figure 2: Scanning electron microscopy of parasites adhering to glass coverslips,
coated [C and D] or not [A and B] with laminin. Arrows show typical amoeboid
citoplasmatic projections.
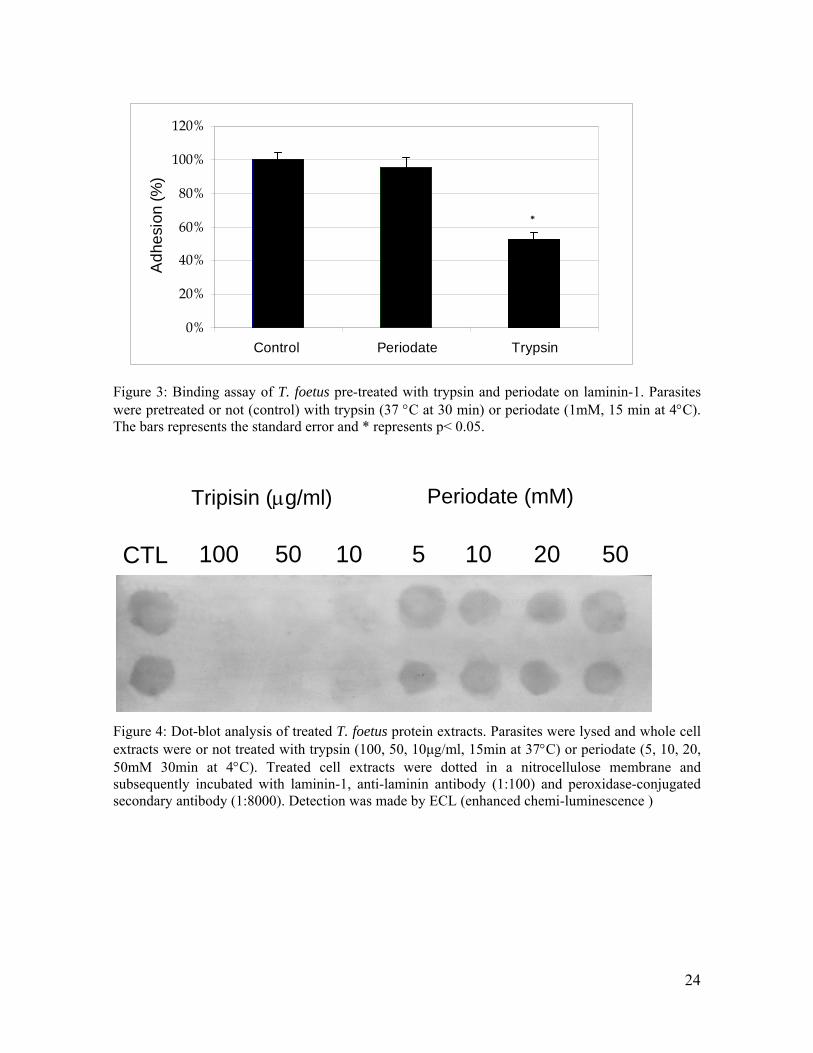
24
Figure 3: Binding assay of T. foetus pre-treated with trypsin and periodate on laminin-1. Parasites were pretreated or not (control) with trypsin (37 °C at 30 min) or periodate (1mM, 15 min at 4°C). The bars represents the standard error and * represents p< 0.05.
Figure 4: Dot-blot analysis of treated T. foetus protein extracts. Parasites were lysed and whole cell extracts were or not treated with trypsin (100, 50, 10μg/ml, 15min at 37°C) or periodate (5, 10, 20, 50mM 30min at 4°C). Treated cell extracts were dotted in a nitrocellulose membrane and subsequently incubated with laminin-1, anti-laminin antibody (1:100) and peroxidase-conjugated secondary antibody (1:8000). Detection was made by ECL (enhanced chemi-luminescence )
0%
20%
40%
60%
80%
100%
120%
Control Periodate Trypsin
Adh
esio
n (%
)
*
CTL
Periodate (mM)
100 50 10
Tripisin (μg/ml)
5 10 20 50

25
Figure 5: Solid phase adhesion assay of Tritrichomonas foetus with laminin. Parasites were poured on laminin-1 coated surface [A] in the presence or not (control) of divalent cations (Ca+2, Mg +2, Mn +2) with or without EDTA (divalent cation quelant); [B] in the presence of sacarose (an carbohydrate without known binding properties) and galactose (sugar commonly found in many extracellular glycoproteins). The bars represents the standart error and * represents p< 0.05.
0,00
1,002,00
3,00
4,00
5,006,00
7,00
8,009,00
10,00
control EDTA Ca+2, Mg+2, Mn+2
Tota
l of a
dher
ed c
ells
(x10
4 )*
* A
00,5
11,5
22,5
33,5
44,5
5
Sacarose galactose
Tota
l of a
dher
ed c
ells
(x10
4 )
B
β lactose

26
Figure 6: Solid-phase adhesion assay of Tritrichomonas foetus with Laminin amino acid sequences AG73 (RKRLQVQLSIRT), C16 (KAFDITYVRLK), A208 (AASIKVAVSADR) and A13 (RQVFQVAYIIIKA). In pre-incubation condition, parasites were pre-incubated with each one of the peptides (30μg/ml) for 30 min at 4°C before contact with Laminin-1 coated cover slips. At the competition assay parasites were poured into Laminin-1 coated surface and overlaid with the peptides (30μg/ml). The amount of attached parasites is given as a percent of the number of parasites that were attached to the wells in the absence of peptides (control). Bars represent standard error.
3,67% 7,33% 9%
100%
12,33%
49%
10,67%
29%
20,33%
0%
20%
40%
60%
80%
100%
120% pa
rasi
tes/
field
control
AG73
C16
A208
A13
Pré-incubation
Competition
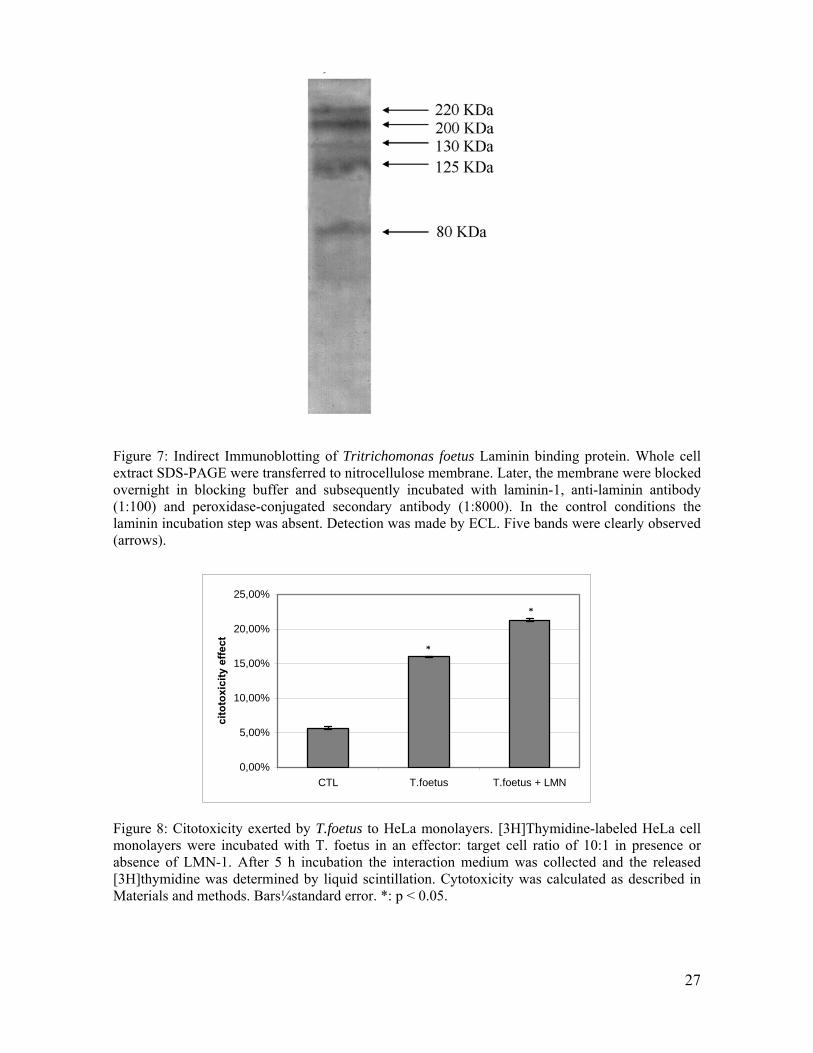
27
Figure 7: Indirect Immunoblotting of Tritrichomonas foetus Laminin binding protein. Whole cell extract SDS-PAGE were transferred to nitrocellulose membrane. Later, the membrane were blocked overnight in blocking buffer and subsequently incubated with laminin-1, anti-laminin antibody (1:100) and peroxidase-conjugated secondary antibody (1:8000). In the control conditions the laminin incubation step was absent. Detection was made by ECL. Five bands were clearly observed (arrows).
Figure 8: Citotoxicity exerted by T.foetus to HeLa monolayers. [3H]Thymidine-labeled HeLa cell monolayers were incubated with T. foetus in an effector: target cell ratio of 10:1 in presence or absence of LMN-1. After 5 h incubation the interaction medium was collected and the released [3H]thymidine was determined by liquid scintillation. Cytotoxicity was calculated as described in Materials and methods. Bars¼standard error. *: p < 0.05.
0,00%
5,00%
10,00%
15,00%
20,00%
25,00%
CTL T.foetus T.foetus + LMN
cito
toxi
city
effe
ct *
*

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo