anaerobic mineralization of pentachlorophenol (pcp) by combining pcp-dechlorinating and...
TRANSCRIPT

ARTICLE
Anaerobic Mineralization of Pentachlorophenol(PCP) by Combining PCP-Dechlorinating andPhenol-Degrading Cultures
Suyin Yang,1 Atsushi Shibata,2 Naoko Yoshida,2 Arata Katayama1,2
1Graduate School of Engineering, Nagoya University, Furo-cho, Chikusa, Nagoya 464-8603,
Japan; telephone: 81-52-789-5856; fax: 81-52-789-5857; e-mail: [email protected] of Integrated Research Projects, EcoTopia Science Institute, Nagoya University,
Furo-cho, Chikusa, Nagoya 464-8603, Japan
Received 17 March 2008; revision received 7 June 2008; accepted 10 June 2008
Published online 18 June 2008 in Wiley InterScience (www.interscience.wiley.com). D
OI 10.1002/bit.22032ABSTRACT: The dechlorination and mineralization of pen-tachlorophenol (PCP) was investigated by simultaneously orsequentially combining two different anaerobic microbialpopulations, a PCP-dechlorinating culture capable of thereductive dechlorination of PCP to phenol and phenol-degrading cultures able to mineralize phenol under sulfate- oriron-reducing conditions. In the simultaneously combinedmixture, PCP (about 35 mM) was mostly dechlorinated tophenol after incubation for 17 days under sulfate-reducingconditions or for 22 days under iron-reducing conditions.Thereafter, the complete removal of phenol occurred within40 days under both conditions. In the sequentially combinedmixture, most of the phenol, the end product of PCPdechlorination, was degraded within 12 days of inoculationwith the phenol degrader, without a lag phase, under bothsulfate- and iron-reducing conditions. In a radioactivityexperiment, [14C–U]–PCP was mineralized to 14CO2 and14CH4 by the combined anaerobic microbial activities. Ana-lysis of electron donor and acceptor utilization and of theproduction and consumption of H2, CO2, and CH4 sug-gested that the dechlorinating and degrading microorgan-isms compete with other microorganisms to perform PCPdechlorination and part of the phenol degradation in com-plex anoxic environments in the presence of electron donorsand acceptors. The presence of a small amount of autoclavedsoil slurry in the medium was possibly another advantageousfactor in the successful dechlorination and mineralization ofPCP by the combined mixtures. This anaerobic–anaerobiccombination technology holds great promise as a cost-effective strategy for complete PCP bioremediation in situ.
Biotechnol. Bioeng. 2009;102: 81–90.
� 2008 Wiley Periodicals, Inc.
Correspondence to: A. Katayama
Contract grant sponsor: Ministry of Education, Science, Sports, and Culture of Japan
Contract grant number: B2:17310045
Contract grant sponsor: Ministry of the Environment (Environmental Technology
Development Fund)
Contract grant sponsor: New Energy and Industrial Technology Development
Organization
� 2008 Wiley Periodicals, Inc.
KEYWORDS: mineralization; chlorinated phenol; sulfatereduction; iron reduction; bioremediation; combination
Introduction
Chlorinated phenols, particularly pentachlorophenol(PCP), have been extensively used as pesticides and woodpreservatives (Crosby, 1981). Because of their toxicity,suspected carcinogenicity, and recalcitrance to biodegrada-tion, the environmental fate of chlorinated phenols hasbecome a primary concern over past decades (Fisher, 1991;McAllister et al., 1996). Although some pure aerobiccultures that can mineralize PCP have been isolated andidentified (Saber and Crawford, 1985; Watanabe, 1973),highly chlorinated phenols are considered to be moreamenable to reductive dechlorination, with the sequentialreplacement of the chlorine atom by a hydrogen atom, to aless-chlorinated phenol or phenol under anaerobic condi-tions. Several anaerobic bacteria that dechlorinate PCP toless-chlorinated phenols have been isolated: Desulfitobacter-ium frappieri PCP-1 of the phylum Firmicutes, whichdechlorinates PCP to 3-chlorophenol (Bouchard et al.,1996), Desulfitobacterium hafniense, which dechlorinatesPCP to 3,4,5-trichlorophenol (Christiansen and Ahring,1996; Madsen and Licht, 1992), and Desulfomonile tirdjeiPCB-1 of the phylum Proteobacteria, which dechlorinatesPCP to 2,4,6-trichlorophenol (2,4,6-TCP; Mohn andKennedy, 1992).
The incomplete reductive dechlorination process underanaerobic conditions decreases, to some extent, the toxicityof highly chlorinated phenols. However, the accumulat-ion of less-chlorinated or dechlorinated compounds as
Biotechnology and Bioengineering, Vol. 102, No. 1, January 1, 2009 81

intermediates still presents a significant threat to theenvironment and human health. Therefore, a subsequenttreatment step is required for the complete mineralization ofthese metabolites. Because these less-chlorinated intermedi-ates are more readily degraded under aerobic conditions,many recent studies have focused on a combined anaerobic–aerobic bioprocess methodology for the complete remedia-tion of chlorinated phenol contamination. Ehlers andRose (2006a) investigated the complete biodegradation of2,4,6-TCP in linked anaerobic and aerobic reactors. Thereductive dechlorination of 2,4,6-TCP to 2,4-dichlorophe-nol, 4-chlorophenol, and phenol was observed undersulfate-reducing conditions in an anaerobic reactor inocu-lated with activated sludge. These metabolites produced inthe dechlorination stage were degraded in the subsequentaerobic reactor by immobilized white-rot fungi. Similarly,the mineralization of 2,4,6-TCP in a two-stage anaerobic–aerobic bioprocess has been reported by Armenante et al.(1999). The authors developed a mathematical model andsucceeded in simulating anaerobic–aerobic processes usingkinetic parameters separately obtained for each process.Wilson et al. (1998) also demonstrated the feasibility of thecomplete removal of PCP in sequential granular activated-carbon fluidized-bed anaerobic and aerobic reactors. Suchintegrated anaerobic–aerobic treatments have also beenreported for the complete removal of other halogenatedorganic and toxic compounds (such as chlorinated ethenes,polychlorinated biphenyls, and azo dyes) (Beeman andBleckmann, 2002; Master et al., 2002; van der Zee andVillaverde, 2005). Although the application of anaerobicconditions followed by aerobic conditions has proved to be afeasible technique for the bioremediation of chlorinated-phenol-contaminated soils and groundwater, the applica-tion of these anaerobic–aerobic combination treatmentsmay be limited because of the relatively high energyconsumption and cost of the aerobic treatment.
For this reason, it is desirable to completely mineralizePCP and chlorophenols under anaerobic conditions. In thepast two decades, the anaerobic oxidation of phenol andmonochlorinated phenols (possible PCP anaerobic reduc-tive dechlorination products) has been described underdenitrifying, sulfate-reducing, iron-reducing, or methano-genic conditions (Bae et al., 2002; Evans and Fuchs, 1988;Haggblom and Young, 1993; Kazumi et al., 1995). A numberof pure phenol-degrading cultures that use nitrate orsulfate as electron acceptors have been isolated (Bak andWiddel, 1986; Boopathy, 1995; Tschech and Fuchs, 1987).A dissimilatory iron-reducing isolate (GS-15) has beenreported that can use phenol (Lovley and Lonergan, 1990).These findings suggest that the anaerobic mineralization ofhighly chlorinated phenols can be achieved by the combinedactivities of a dechlorinating population and an anaerobi-cally oxidizing population of bacteria. However, the twopopulations have different physiological characteristics intheir use of electron donors and acceptors, and no suchcombination technology has been seriously considered untilnow.
82 Biotechnology and Bioengineering, Vol. 102, No. 1, January 1, 2009
In this study, we describe two methods of combining twodifferent anaerobic microbial populations to mineralize PCPin a laboratory-scale batch experiment: a PCP dechlorinatorcapable of dechlorinating PCP to phenol and phenoldegraders able to mineralize phenol under sulfate- or iron-reducing conditions. The utilization of electron donors andacceptors, the production of respiration gases (H2, CO2,CH4), and the metabolites of PCP were analyzed to producea more detailed picture of the relevant biotic reactions inthese combined systems.
Materials and Methods
PCP-Dechlorinating Enrichment Culture
The anaerobic PCP-dechlorinating culture was obtained anddeveloped from an uncontaminated paddy soil collectedfrom Yatomi-cho, Japan. PCP was dechlorinated in sequenceto 2,3,4,5-tetrachlorophenol, then to 3,4,5-trichlorophenol,3,5-dichlorophenol, 3-chlorophenol, and phenol in thisculture, supplemented with 20 mM lactate and a smallamount of sterile soil slurry as a required nutritionalsupplement (Yoshida et al., 2007). A Clostridium species ofthe phylum Firmicutes was considered to dominate themicrobial community. The detailed procedure for itsenrichment and microbial characterization has beenpublished previously (Yoshida et al., 2007). Before thisstudy was begun, the dechlorinating culture was transferred70 times in the enrichment procedure. The culturedechlorinated PCP, at concentrations up to 100 mM, tophenol in 2 weeks.
Phenol-Degrading Soil-Free Enrichment Cultures
Anaerobic phenol-degrading soil-free cultures wereenriched from an uncontaminated paddy soil collectedfrom the Kamajima area of Aichi prefecture, Japan, by serialsubculturing. Briefly, 5 mL of soil slurry, which wasperiodically refed with phenol for 3 years, was transferred toa 60 mL serum bottle filled with 40 mL of sterile anaerobicmineral salt medium. Anoxic phenol solution was addedas the sole carbon and energy source. Sodium sulfate oramorphous Fe(III) oxide (FeOOH) stock solution wasadded anaerobically as the electron acceptor, to a finalconcentration of 10 mM. When the phenol was significantlydepleted, 20% of the culture was transferred and sub-cultured under the same conditions as used for enrichment.The detailed enrichment methods and the microbialcommunity characterization of these two cultures havebeen described previously (Yang et al., 2007). Phenol(maximum concentration 100 mM) was completely miner-alized in 3 weeks under sulfate-reducing conditions andthe culture was predominated by d-Proteobacteria. Phenol(at a maximum concentration of 700 mM) was degraded in1 month under iron-reducing conditions and Geobacter and

an unidentified species with 96% similarity to an unculturedclone in petroleum-contaminated sediment were detected inthe culture. The cultures were transferred 15 times beforethey were used in this study.
Degradation of PCP by Combining Two AnaerobicEnrichment Cultures
Simultaneous combination and sequential combinationmethods were used in two different approaches to combinethe PCP-dechlorinating and phenol-degrading enrichmentcultures for the mineralization of PCP in batch experiments.The simultaneous combination was performed according tothe following procedure: 1.5 mL of the PCP-dechlorinatingenrichment culture and 4 mL of the phenol-degradingsulfate-reducing culture were inoculated into a 60 mL serumbottle with 25 mL of autoclaved oxygen-free sterile soilslurry medium, containing about 3 g (dry weight) of soil,20 mM sodium lactate, 10 mM sodium sulfate, and about40 mM PCP (Condition 1). The sequential combination wasperformed as follows: 1.5 mL of PCP-dechlorinating culturewas transferred to a serum bottle with slurry medium,lactate, and PCP, as described above. After the PCP wasdechlorinated to phenol, 4 mL of phenol-degrading sulfate-reducing culture and 0.3 mmol of sulfate were added to thesame sample bottle (Condition 2). A sample without thecombination was also established as the active control, inwhich only 1.5 mL of the PCP-dechlorinating enrichmentculture was inoculated into the autoclaved soil slurrymedium supplemented with lactate and PCP (Condition 3).Condition 3 was the same as the PCP dechlorination stepfor the sequential combination of cultures in Condition 2.Similarly, phenol-degrading iron-reducing culture was alsosimultaneously or sequentially combined in the samemanner with the PCP-dechlorinating culture, and 20 mMFeOOH (0.6 mmol per bottle) was added instead of sulfate(Conditions 4 and 5, respectively).
All inoculations of the cultures and additions of nutrientswere made with sterile plastic syringes and 22G needlesthrough the rubber stoppers. The sample bottles wereincubated statically in triplicate or duplicate in the dark at308C. Samples (1 mL) were taken periodically during theincubation after the bottles had been shaken by hand, toanalyze PCP degradation, ions, and organic fatty acids,according to the methods described below.
Anaerobic Mineralization of 14C–PCP byCombined Cultures
The mineralization of PCP in the simultaneously combinedanaerobic culture was confirmed using [14C–ring (U)]–PCP(99%, 80 mCi mmol�1; Sigma Chemical Co., St. Louis,MO). The experimental conditions were the same asConditions 1, 2, and 4 described above, except that [14C–U]–PCP–toluene (about 5 kBq, 1� 10�3 mmol) and un-labeled PCP–sodium stock solution (1.2 mmol, 319.2 mg)
were added as 100 mL of acetone solution. The radioactivityadded was 2.9� 105 dpm per bottle and the total initialconcentration of PCP was approximately 40 mM. An abioticcontrol was established in parallel by autoclaving thesamples at 1218C for 1 h and then adding paraformaldehyde.All the samples (six replicates for each condition) wereincubated as described above.
After incubation for 2 months, the headspace gases of thethree replicate samples were scrubbed by flushing with N2
gas for 15 min after acidification with 6 N HCl, and werepassed through 200 mL flasks filled with sponge to trap theradioactive hydrophobic vapors. They were then collectedin two connected 30 mL glass tubes containing 10 mL of asolution of ethanolamine/methanol (1:1) to trap 14CO2. Tomeasure the 14C-radiolabeled methane, a combustion quartztube packed with cupric oxide, to convert the 14CH4 to14CO2 at 8508C, and an extra pure oxygen line (with amaximum flow of 10 mL min�1) were connected betweenthe flask and the glass tubes. Another three samples werepurged with pure oxygen instead of nitrogen at a rate ofapproximately 20 mL min�1 for 4 h in this combustion/trapping line to calculate the total radioactivity of the 14CH4
and 14CO2. The methane combustion efficiency and carbon-dioxide-trapping efficiency of the apparatus, determinedusing 13CH4 and 13CO2, were 30% and 50%, respectively.The acidified samples remaining after flushing with gas werecentrifuged at 8,000g for 15 min and the radioactivitypresent in the aqueous phase was counted. A Tris-bufferedsolution was then added to the soil sample to adjust the pHto 9 to dissolve the PCP adsorbed to the soil. The sampleswere centrifuged and the radioactivity was measured in thealkaline supernatant. Finally, 0.3 g of the remaining soil wasweighed and burned in an automatic sample combustionsystem for 3 min to collect and analyze the remaining 14C inthe soil as 14CO2. All the 14C radioactivities were determinedwith a liquid scintillation counter (LSC 5100, Aloka, Tokyo,Japan) in 5 mL of liquid scintillation cocktail (Clear-sol II,Nacalai Tesque Inc., Kyoto, Japan).
Chemical Analysis Techniques
PCP, phenol, and the other metabolites produced duringthe reductive dechlorination of PCP were extracted andanalyzed using a gas chromatograph–mass spectrometer(GC–MS) equipped with a DB-5MS column (Shimadzu,Kyoto, Japan), as described by Yoshida et al. (2007). A liquidsample (1 mL) was analyzed after dilution and filtration(0.45 mm pore diameter; MillexTM-LH, Millipore Corpora-tion, MA) for sulfate and chloride by ion chromatography(761 Compact ion chromatograph with an SI-90 4E columnand a conductivity detector), using 1.8 mM Na2CO3 and1.7 mM NaHCO3 as the eluent phase. Fe(II) was measuredspectrophotometrically using a modified 1,10-phenanthro-line method, as described previously (Yang et al., 2007).Organic acids (lactate, acetate, propionate) were analyzedwith a high-performance liquid chromatograph (CTO-10A;
Yang et al.: Mineralization of PCP by Two Microbial Populations 83
Biotechnology and Bioengineering
Shimadzu) equipped with an L-column ODS (4.6 mm innerdiameter, 250 mm in length; CRRI, Saitama, Japan) and UVdetection (210 nm). The mobile phase contained 0.1%phosphoric acid and 2% acetonitrile. Hydrogen, methane,and carbon dioxide in the headspaces were collectedwith 100 mL gastight syringes and were analyzed on a gaschromatograph (GC-14B, Shimadzu) equipped with athermal conductivity detector. The concentration of thegas dissolved in the aqueous solution was calculatedaccording to the equilibrium equation described previouslyby Mazur et al. (2003).
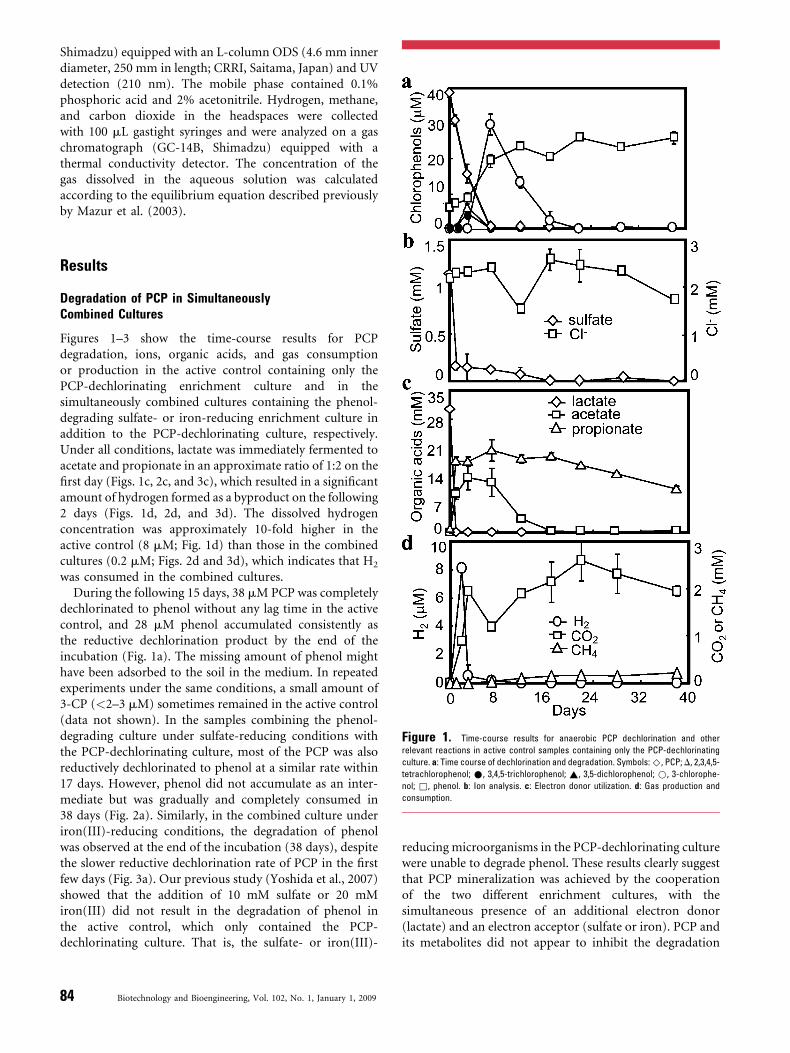
Figure 1. Time-course results for anaerobic PCP dechlorination and other
relevant reactions in active control samples containing only the PCP-dechlorinating
culture. a: Time course of dechlorination and degradation. Symbols: ^, PCP; D, 2,3,4,5-
tetrachlorophenol; *, 3,4,5-trichlorophenol; ~, 3,5-dichlorophenol; *, 3-chlorophe-
nol; &, phenol. b: Ion analysis. c: Electron donor utilization. d: Gas production and
consumption.
Results
Degradation of PCP in SimultaneouslyCombined Cultures
Figures 1–3 show the time-course results for PCPdegradation, ions, organic acids, and gas consumptionor production in the active control containing only thePCP-dechlorinating enrichment culture and in thesimultaneously combined cultures containing the phenol-degrading sulfate- or iron-reducing enrichment culture inaddition to the PCP-dechlorinating culture, respectively.Under all conditions, lactate was immediately fermented toacetate and propionate in an approximate ratio of 1:2 on thefirst day (Figs. 1c, 2c, and 3c), which resulted in a significantamount of hydrogen formed as a byproduct on the following2 days (Figs. 1d, 2d, and 3d). The dissolved hydrogenconcentration was approximately 10-fold higher in theactive control (8 mM; Fig. 1d) than those in the combinedcultures (0.2 mM; Figs. 2d and 3d), which indicates that H2
was consumed in the combined cultures.During the following 15 days, 38 mM PCP was completely
dechlorinated to phenol without any lag time in the activecontrol, and 28 mM phenol accumulated consistently asthe reductive dechlorination product by the end of theincubation (Fig. 1a). The missing amount of phenol mighthave been adsorbed to the soil in the medium. In repeatedexperiments under the same conditions, a small amount of3-CP (<2–3 mM) sometimes remained in the active control(data not shown). In the samples combining the phenol-degrading culture under sulfate-reducing conditions withthe PCP-dechlorinating culture, most of the PCP was alsoreductively dechlorinated to phenol at a similar rate within17 days. However, phenol did not accumulate as an inter-mediate but was gradually and completely consumed in38 days (Fig. 2a). Similarly, in the combined culture underiron(III)-reducing conditions, the degradation of phenolwas observed at the end of the incubation (38 days), despitethe slower reductive dechlorination rate of PCP in the firstfew days (Fig. 3a). Our previous study (Yoshida et al., 2007)showed that the addition of 10 mM sulfate or 20 mMiron(III) did not result in the degradation of phenol inthe active control, which only contained the PCP-dechlorinating culture. That is, the sulfate- or iron(III)-
84 Biotechnology and Bioengineering, Vol. 102, No. 1, January 1, 2009
reducing microorganisms in the PCP-dechlorinating culturewere unable to degrade phenol. These results clearly suggestthat PCP mineralization was achieved by the cooperationof the two different enrichment cultures, with thesimultaneous presence of an additional electron donor(lactate) and an electron acceptor (sulfate or iron). PCP andits metabolites did not appear to inhibit the degradation
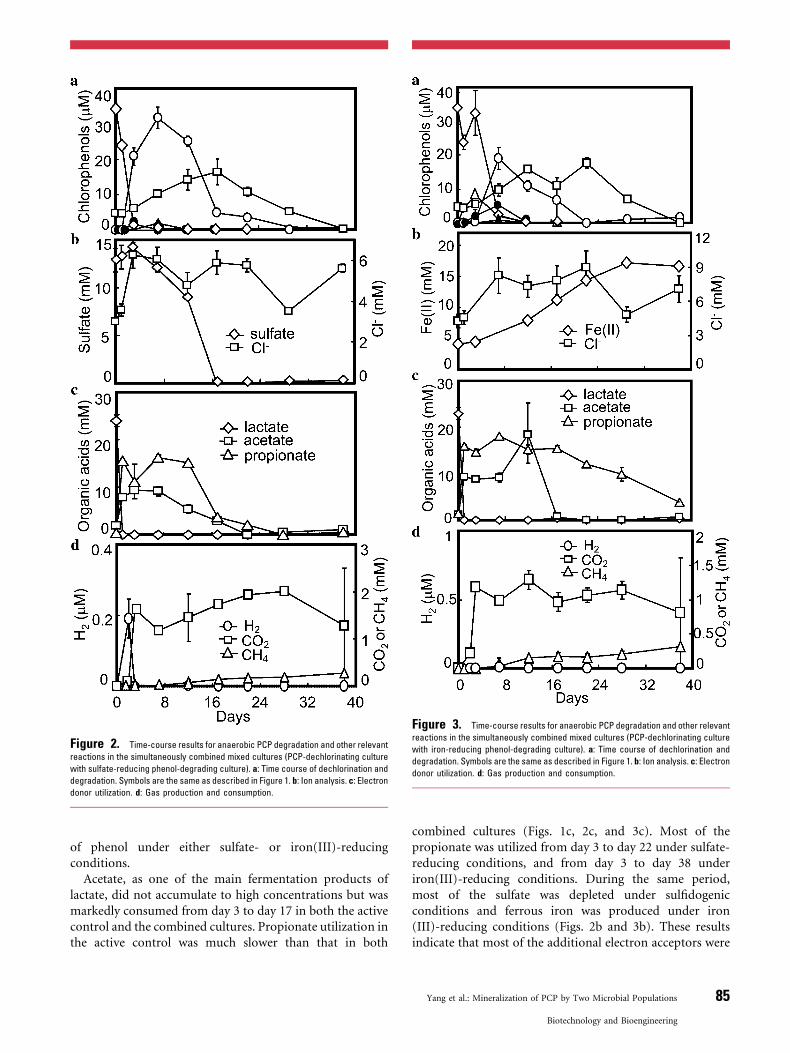
Figure 2. Time-course results for anaerobic PCP degradation and other relevant
reactions in the simultaneously combined mixed cultures (PCP-dechlorinating culture
with sulfate-reducing phenol-degrading culture). a: Time course of dechlorination and
degradation. Symbols are the same as described in Figure 1. b: Ion analysis. c: Electron
donor utilization. d: Gas production and consumption.
Figure 3. Time-course results for anaerobic PCP degradation and other relevant
reactions in the simultaneously combined mixed cultures (PCP-dechlorinating culture
with iron-reducing phenol-degrading culture). a: Time course of dechlorination and
degradation. Symbols are the same as described in Figure 1. b: Ion analysis. c: Electron
donor utilization. d: Gas production and consumption.
of phenol under either sulfate- or iron(III)-reducingconditions.
Acetate, as one of the main fermentation products oflactate, did not accumulate to high concentrations but wasmarkedly consumed from day 3 to day 17 in both the activecontrol and the combined cultures. Propionate utilization inthe active control was much slower than that in both
combined cultures (Figs. 1c, 2c, and 3c). Most of thepropionate was utilized from day 3 to day 22 under sulfate-reducing conditions, and from day 3 to day 38 underiron(III)-reducing conditions. During the same period,most of the sulfate was depleted under sulfidogenicconditions and ferrous iron was produced under iron(III)-reducing conditions (Figs. 2b and 3b). These resultsindicate that most of the additional electron acceptors were
Yang et al.: Mineralization of PCP by Two Microbial Populations 85
Biotechnology and Bioengineering
directly consumed by the supplied electron donor or itsfermentation products.
After the first few days, the concentration of hydrogen wasmaintained below the limit of detection in both combinedcultures and in the active control (Figs. 1d, 2d, and 3d). Thisis consistent with the results of previous research, whichshowed that a low aqueous hydrogen concentration wasmaintained during the reductive dehalogenation of chlor-ophenols (approximately 0.34–5 nM) (Mazur et al., 2003)and sulfate reduction (1–1.5 nM) or iron reduction (0.2 nM)(Lovley, 1988). Most carbon dioxide was produced duringthe initial 3 days and increased only slightly in the followingincubation period, and tended to decline at the end of theincubation period under all three conditions (Figs. 1d, 2d,and 3d). Methane was detected after 6 days of incubationand increased continuously thereafter.
The concentration of chloride ions tended to increaseduring PCP dechlorination under all three conditions(Figs. 1b, 2b, and 3b), because of the high backgroundchloride concentration in the sterile soil, so the mass balanceof PCP dechlorination and chloride production could not becalculated.
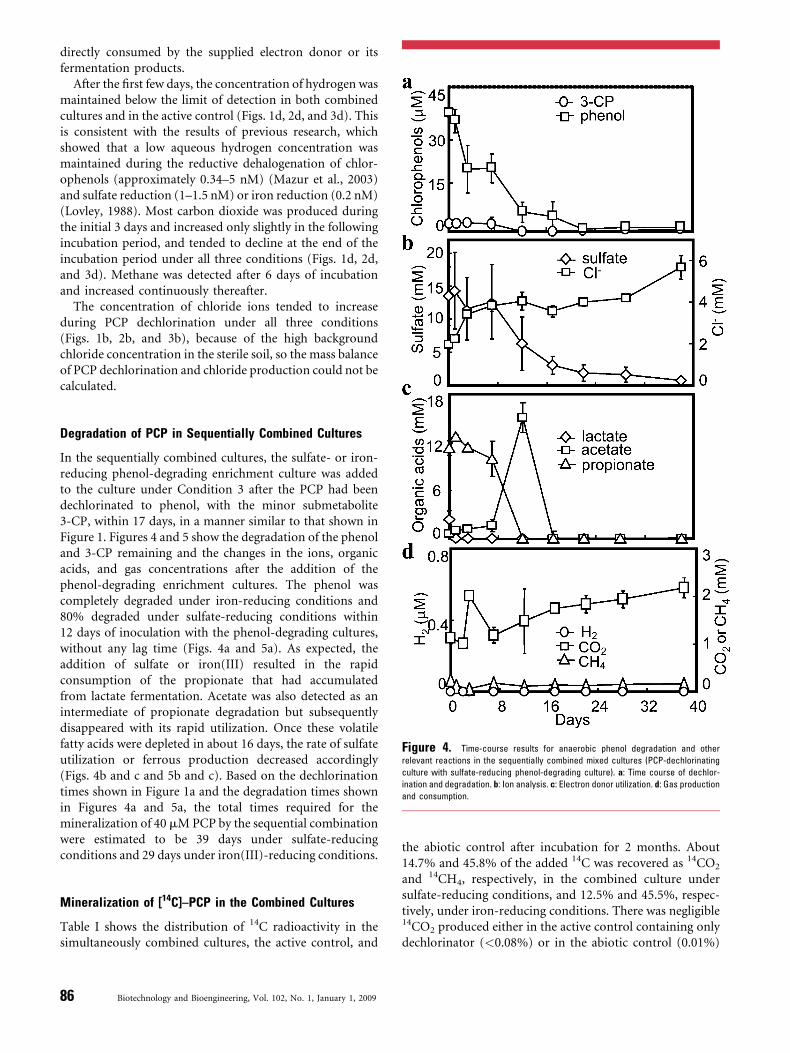
Figure 4. Time-course results for anaerobic phenol degradation and other
relevant reactions in the sequentially combined mixed cultures (PCP-dechlorinating
culture with sulfate-reducing phenol-degrading culture). a: Time course of dechlor-
ination and degradation. b: Ion analysis. c: Electron donor utilization. d: Gas production
and consumption.
Degradation of PCP in Sequentially Combined Cultures
In the sequentially combined cultures, the sulfate- or iron-reducing phenol-degrading enrichment culture was addedto the culture under Condition 3 after the PCP had beendechlorinated to phenol, with the minor submetabolite3-CP, within 17 days, in a manner similar to that shown inFigure 1. Figures 4 and 5 show the degradation of the phenoland 3-CP remaining and the changes in the ions, organicacids, and gas concentrations after the addition of thephenol-degrading enrichment cultures. The phenol wascompletely degraded under iron-reducing conditions and80% degraded under sulfate-reducing conditions within12 days of inoculation with the phenol-degrading cultures,without any lag time (Figs. 4a and 5a). As expected, theaddition of sulfate or iron(III) resulted in the rapidconsumption of the propionate that had accumulatedfrom lactate fermentation. Acetate was also detected as anintermediate of propionate degradation but subsequentlydisappeared with its rapid utilization. Once these volatilefatty acids were depleted in about 16 days, the rate of sulfateutilization or ferrous production decreased accordingly(Figs. 4b and c and 5b and c). Based on the dechlorinationtimes shown in Figure 1a and the degradation times shownin Figures 4a and 5a, the total times required for themineralization of 40 mM PCP by the sequential combinationwere estimated to be 39 days under sulfate-reducingconditions and 29 days under iron(III)-reducing conditions.
Mineralization of [14C]–PCP in the Combined Cultures
Table I shows the distribution of 14C radioactivity in thesimultaneously combined cultures, the active control, and
86 Biotechnology and Bioengineering, Vol. 102, No. 1, January 1, 2009
the abiotic control after incubation for 2 months. About14.7% and 45.8% of the added 14C was recovered as 14CO2
and 14CH4, respectively, in the combined culture undersulfate-reducing conditions, and 12.5% and 45.5%, respec-tively, under iron-reducing conditions. There was negligible14CO2 produced either in the active control containing onlydechlorinator (<0.08%) or in the abiotic control (0.01%)
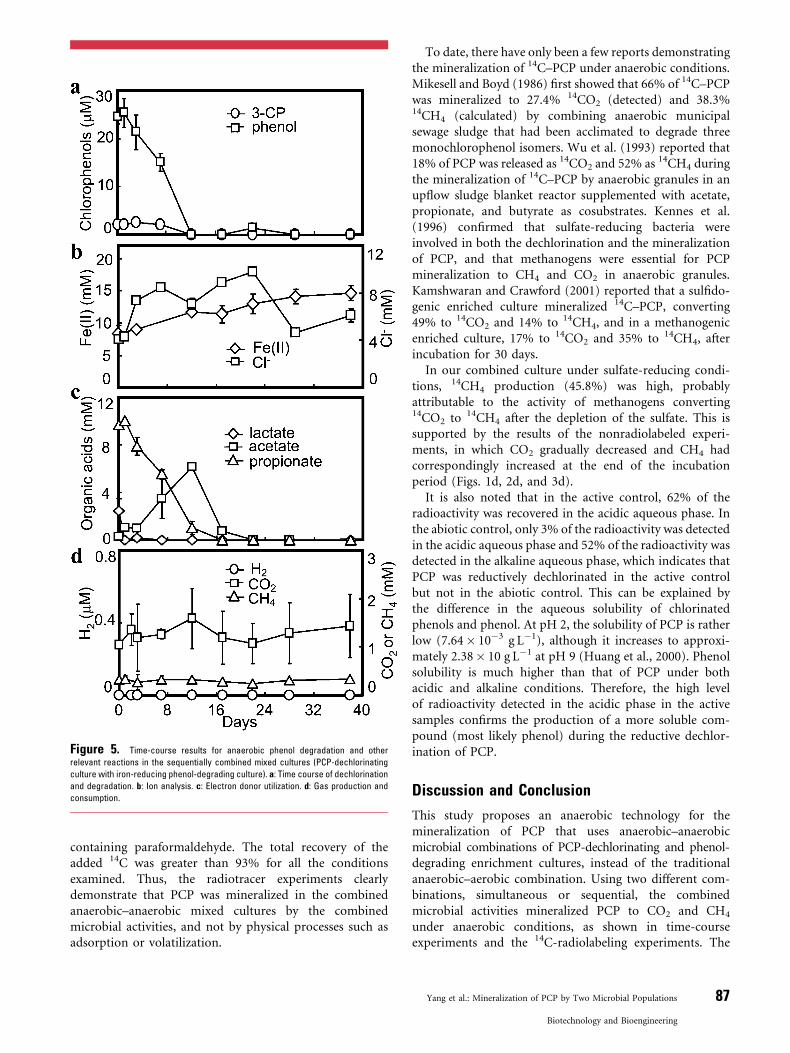
Figure 5. Time-course results for anaerobic phenol degradation and other
relevant reactions in the sequentially combined mixed cultures (PCP-dechlorinating
culture with iron-reducing phenol-degrading culture). a: Time course of dechlorination
and degradation. b: Ion analysis. c: Electron donor utilization. d: Gas production and
consumption.
containing paraformaldehyde. The total recovery of theadded 14C was greater than 93% for all the conditionsexamined. Thus, the radiotracer experiments clearlydemonstrate that PCP was mineralized in the combinedanaerobic–anaerobic mixed cultures by the combinedmicrobial activities, and not by physical processes such asadsorption or volatilization.
To date, there have only been a few reports demonstratingthe mineralization of 14C–PCP under anaerobic conditions.Mikesell and Boyd (1986) first showed that 66% of 14C–PCPwas mineralized to 27.4% 14CO2 (detected) and 38.3%14CH4 (calculated) by combining anaerobic municipalsewage sludge that had been acclimated to degrade threemonochlorophenol isomers. Wu et al. (1993) reported that18% of PCP was released as 14CO2 and 52% as 14CH4 duringthe mineralization of 14C–PCP by anaerobic granules in anupflow sludge blanket reactor supplemented with acetate,propionate, and butyrate as cosubstrates. Kennes et al.(1996) confirmed that sulfate-reducing bacteria wereinvolved in both the dechlorination and the mineralizationof PCP, and that methanogens were essential for PCPmineralization to CH4 and CO2 in anaerobic granules.Kamshwaran and Crawford (2001) reported that a sulfido-genic enriched culture mineralized 14C–PCP, converting49% to 14CO2 and 14% to 14CH4, and in a methanogenicenriched culture, 17% to 14CO2 and 35% to 14CH4, afterincubation for 30 days.
In our combined culture under sulfate-reducing condi-tions, 14CH4 production (45.8%) was high, probablyattributable to the activity of methanogens converting14CO2 to 14CH4 after the depletion of the sulfate. This issupported by the results of the nonradiolabeled experi-ments, in which CO2 gradually decreased and CH4 hadcorrespondingly increased at the end of the incubationperiod (Figs. 1d, 2d, and 3d).
It is also noted that in the active control, 62% of theradioactivity was recovered in the acidic aqueous phase. Inthe abiotic control, only 3% of the radioactivity was detectedin the acidic aqueous phase and 52% of the radioactivity wasdetected in the alkaline aqueous phase, which indicates thatPCP was reductively dechlorinated in the active controlbut not in the abiotic control. This can be explained bythe difference in the aqueous solubility of chlorinatedphenols and phenol. At pH 2, the solubility of PCP is ratherlow (7.64� 10�3 g L�1), although it increases to approxi-mately 2.38� 10 g L�1 at pH 9 (Huang et al., 2000). Phenolsolubility is much higher than that of PCP under bothacidic and alkaline conditions. Therefore, the high levelof radioactivity detected in the acidic phase in the activesamples confirms the production of a more soluble com-pound (most likely phenol) during the reductive dechlor-ination of PCP.
Discussion and Conclusion
This study proposes an anaerobic technology for themineralization of PCP that uses anaerobic–anaerobicmicrobial combinations of PCP-dechlorinating and phenol-degrading enrichment cultures, instead of the traditionalanaerobic–aerobic combination. Using two different com-binations, simultaneous or sequential, the combinedmicrobial activities mineralized PCP to CO2 and CH4
under anaerobic conditions, as shown in time-courseexperiments and the 14C-radiolabeling experiments. The
Yang et al.: Mineralization of PCP by Two Microbial Populations 87
Biotechnology and Bioengineering

Table I. Distribution of 14C in different condition samples after 2 months of incubation.
Sample 14CO2 (%) 14CH4 (%)
Water phase 14C
(%), pH 2
Water phase 14C
(%), pH 9
14C remaining in
soil (%)
Total 14C
(%)
Active sample (sulfate) 14.7� 2.4 45.8� 5.4 16.7� 3.5 4.6� 1.3 16.9� 4.4 98.7� 4.5
Active sample (iron(III)) 12.5� 1.6 45.5� 5.6 11.4� 2.9 4.5� 2.2 22.0� 2.7 95.9� 2.9
Active control 0.08� 0.1 — 62.3� 10.6 13.7� 3.8 20.9� 5.8 96.7� 4.9
Abiotic control 0.01 — 2.7� 0.6 51.6� 13.7 39.3� 6.2 93.1� 12.6
The results are means of three or four incubations� standard deviations; ‘‘—’’ means samples were not analyzed for 14CH4.
simultaneously combined cultures mineralized 10 mg L�1
PCP within 38 days (about 0.25 mg day�1) under bothsulfate-reducing and iron-reducing conditions. In ananaerobic–aerobic combination using a fluidized-bed reactor,Wilson et al. (1998) reported that 200 mg L�1 PCP wasstoichiometrically converted to chlorophenol (CP) orphenol in 16.8–2.3 h, after which the CP and phenol weredegraded quickly and completely in the subsequent aerobicbioreactor. Although the PCP mineralization rate in ourcombined culture was lower than that in the anaerobic–aerobic treatment, such an anaerobic–anaerobic combina-tion may significantly reduce the energy required to supplyoxygen during the aerobic treatment and may avoid thedamaging exposure of sensitive anaerobic organisms tooxygen. Therefore, the combined anaerobic–anaerobictreatment is of great importance and is potentially a feasibleand more cost-effective strategy for the complete removal ofPCP contamination. The simultaneous combination isespecially advantageous in the bioaugmentation treatmentof the source zones of contaminated sites in situ, whereasthe sequential combination, as two connected anaerobicbiobarriers, is advantageous in the remediation of con-taminant plumes in groundwater, because it avoids thesimultaneous addition of both an electron donor and anelectron acceptor, thus reducing their useless consumption.
Our detailed comparison of the production andconsumption of ions, volatile fatty acids, and gases, togetherwith PCP mineralization (shown in Fig. 2), suggests themain biochemical reactions associated with PCP miner-alization. First, the production of approximately 8 mMacetate and 16 mM propionate from around 24 mM lactateconfirms the reaction pathway of lactate fermentation, asreported previously (Thauer et al., 1977): 3 lactate� !acetate�þ2 propionate�þHCO�
3 þHþ and lactate�þ2H2O
! acetate� þ 2H2 þ HCO�3 þ Hþ. Except in the first few
days, hydrogen was not detected during the incubationperiod, which is consistent with the previously drawnconclusion that the hydrogen produced by fermentativemicroorganisms in an anoxic environment is rapidlyconsumed and maintained at low levels by terminalmicroorganisms, such as sulfate-reducers or methanogens(Harris et al., 2007). H2 has been reported to be an effectiveelectron donor for the reductive dechlorination process inenriched cultures and bacterial isolates (El Fantroussi et al.,1998; Fung et al., 2007). In this study, when facilitated by an
88 Biotechnology and Bioengineering, Vol. 102, No. 1, January 1, 2009
electron donor (possibly H2), the dechlorination of theadded 35 mM PCP should result in the formation of 35 mMphenol based on the equation for the reductive dechlorina-tion of PCP: 2PCPþ 5H2 ! 2 phenolþ 10Hþþ 10Cl�.Actually, only 16 mM phenol was detected at maximum,which is lower than that ideally formed 35 mM or the 28 mMphenol formed in the active control. These results indicatethat phenol-degrading culture might commence utilizingphenol during the production of phenol from PCP dechlori-nation, even though the reaction using acetate and propio-nate is thermodynamically more favorable than the reactionin which phenol is the electron donor (Liamlean andAnnachhatre, 2007). Phenol was anaerobically oxidized withthe reduction of sulfate as the electron acceptor: 2 phenolþ7SO2�
4 þ 6H2O ! 12HCO�3 þ 7S2� þ 12Hþ. Under iron-
(III)-reducing conditions, phenol degradation is consideredto be as follows: phenol þ 28Fe3þ þ 17H2O ! 6HCO�
3 þ28Fe2þ þ 34Hþ. Accordingly, the amount or concentrationratio of electron donors and acceptors should have asignificant effect on the PCP mineralization efficiency in thecombined culture. The optimization of the electron donorsand acceptors is currently underway to increase the activityof the combined culture for the complete mineralizationof PCP.
We observed that phenol degradation did not cease evenafter sulfate depletion (Fig. 2). A polymerase chain reaction(PCR)–denaturing gradient gel electrophoresis (DGGE)analysis of 16S rRNA genes (Yang et al., 2007) suggested thatour phenol-degrading enrichment culture under sulfate-reducing conditions harbored a microorganism with 92%similarity to a recently isolated strictly anaerobic syntrophicbacterial strain, UIT, which oxidized phenol in associationwith an H2-utilizing methanogen (Qiu et al., 2008).Interestingly, when 10 mM sulfate was added, strain UIT
grew even faster with the Desulfovibrio-like bacterium(sulfate reducer) than with the methanogen as its H2-scavenging partner. Desulfovibrio species were also present inour culture (data not shown). It is possible that, in thecombined culture, phenol was utilized by a syntrophicmicroorganism in association with the H2-scavengingsulfate reducer. When the sulfate was completely consumed,the methanogenic bacteria could have taken over the roleof H2 scavenger, and CO2 could have been converted to CH4
during phenol degradation, as shown in Figure 2a. Furtherstudy is required to identify the microorganisms involved in

PCP mineralization in the combined culture after thedepletion of sulfate.
The simultaneously combined culture allowed theconcurrent reactions to act cooperatively: the reductivedechlorination of PCP to phenol, the anaerobic oxidation ofphenol, the H2-scavenging reactions (sulfate reduction oriron[III] reduction), and organic acid oxidization. It hasoften been considered that the occurrence of alternativeelectron acceptors (especially sulfate) with reducing micro-organisms inhibits the dechlorinating microorganismsbecause of the competition between the microorganismsfor electron donors (Kohring et al., 1989; Madsen andAamand, 1991). Therefore, the addition of excess electrondonors has been recommended to completely deplete thealternative electron acceptors and to enhance the dechlor-ination process (ESTCP, 2001). However, dechlorinationoccurred under sulfate-reducing conditions in our com-bined culture, as other recent studies have reported (Ehlersand Rose, 2006b; Hoelen and Reinhard, 2004). Moreover,Aulenta et al. (2007) have demonstrated that it may not benecessary to consume sulfate by supplying an electron donorto achieve substantial dechlorination.
It should be noted that our combined cultures containeda small amount of autoclaved soil slurry. Our PCP-dechlorinating enrichment culture lost its dechlorinatingactivity without the autoclaved soil slurry. Furthermore, noPCP dechlorination was observed in cultures containingother materials, such as glass beads (0.5 mm) or organic-matter-free burnt soils instead of the autoclaved soil slurry(data not shown). These results indicate that the organicmatter in the soil slurry may have acted as an essentialgrowth factor in the dechlorinating cultures. Ehlers andRose (2006a) have demonstrated that anaerobic 2,4,6-TCP-dechlorinating bacterial colonies were only detectable in thesoil fractions in inoculated flasks, and not in the liquidfractions. They therefore assumed that the surfaces ofthe soil contributed to the maintenance of cell densities.Moreover, increased soil organic matter and soil watercontent have been reported to enhance PCP degradation(Kuwatsuka and Igarashi, 1975; Seech et al., 1991). A soilslurry might also serve as a structural backbone, allowing theattachment of different microbial species (Sekiguchi et al.,1999), and thus increasing the cooperation efficiency of thedifferent microorganisms. It will be interesting to clarify theroles of the soil slurry in the mineralization of PCP.
In summary, we have demonstrated the mineralization ofPCP by the simultaneous or sequential combination of aPCP-dechlorinating enrichment culture and a phenol-degrading enrichment culture under anaerobic conditions.The main relevant biotic reactions in the combined systemssuggest that the dechlorinating and degrading micro-organisms cooperated in PCP dechlorination and phenoldegradation in an anoxic culture in the presence of electrondonors and acceptors. This successful batch anaerobic–anaerobic combination presents an attractive and cost-effective technology for the bioremediation of PCP andother chlorinated aromatic compounds. This combined
technology is currently being applied in the design of acolumn system that can be used in situ.
The authors wish to thank the members of Aichi-ken Agricultural
Research Center for providing the soil samples. This research was
partly supported by a Grant-in-Aid from the Ministry of Education,
Science, Sports, and Culture of Japan (B2:17310045), by the Envir-
onmental Technology Development Fund of the Ministry of the
Environment, and by the New Energy and Industrial Technology
Development Organization.
References
Armenante MP, Kafkewitz D, Lewandowski AG, Jou CJ. 1999. Anaerobic–
aerobic treatment of halogenated phenolic compounds. Water Res
33:681–692.
Aulenta F, Pera A, Rossetti S, Papini MP, Majone M. 2007. Relevance of side
reactions in anaerobic reductive dechlorination microcosms amended
with different electron donors. Water Res 41:27–38.
Bae HS, Yamagishi T, Suwa Y. 2002. Developing and sustaining 3-chlor-
ophenol-degrading populations in up-flow anaerobic column reactors
under circum-denitrifying conditions. Appl Microbiol Biotechnol 59:
118–124.
Bak F, Widdel F. 1986. Anaerobic degradation of phenol and phenol
derivatives by Desulfobacterium phenolicum sp. nov. Arch Microbiol
46:177–180.
Beeman RE, Bleckmann CA. 2002. Sequential anaerobic–aerobic treatment
of an aquifer contaminated by halogenated organics: Field results.
J Contam Hydrol 57:147–159.
Boopathy R. 1995. Isolation and characterization of a phenol-degrading,
sulfate-reducing bacterium from swine manure. Bioresource Technol
54:29–33.
Bouchard B, Beaudet R, Villemur R, McSween G, Lepine F, Bisaillon JG.
1996. Isolation and characterization of Desulfitobacterium frappieri
sp. nov., an anaerobic bacterium which reductively dechlorinates
pentachlorophenol to 3-chlorophenol. Int J Syst Bacteriol 46:1010–
1015.
Christiansen N, Ahring BK. 1996. Desulfitobacterium hafniense sp. nov., an
anaerobic, reductively dechlorinating bacterium. Int J Syst Bacteriol
46:442–448.
Crosby DG. 1981. Environmental chemistry of pentachlorophenol. Pure
Appl Chem 53:1051–1080.
Ehlers GA, Rose PD. 2006a. An integrated anaerobic/aerobic bioprocess for
the remediation of chlorinated phenol-contaminated soil and ground-
water. Water Environ Res 78:701–709.
Ehlers GA, Rose PD. 2006b. The potential for reductive dehalogenation of
chlorinated phenol in a sulphidogenic environment in in situ enhanced
biodegradation. Water SA 32:243–248.
El Fantroussi S, Naveau H, Agathos SN. 1998. Anaerobic dechlorinating
bacteria. Biotechnol Prog 14:167–188.
ESTCP. 2001. http://www.estcp.org/documents/techdocs/Rabitt_Update.
pdf.
Evans WC, Fuchs G. 1988. Anaerobic degradation of aromatic compounds.
Annu Rev Microbiol 42:289–317.
Fisher B. 1991. Pentachlorophenol: Toxicology and environmental fate.
J Pestic Reform 11:2–5.
Fung JM, Morris RM, Adrian L, Zinder SH. 2007. Expression of reductive
dehalogenase genes in Dehalococcoides ethenogenes strain 195 growing
on tetrachloroethene, trichloroethene, or 2,3-dichlorophenol. Appl
Environ Microbiol 73:4439–4445.
Haggblom MM, Young LY. 1993. Influence of alternative electron acceptors
on the anaerobic biodegradability of chlorinated phenols and benzoic
acids. Appl Environ Microbiol 59:1162–1167.
Harris SH, Smith RL, Suflita JM. 2007. In situ hydrogen consumption
kinetics as an indicator of subsurface microbial activity. FEMS Micro-
biol Ecol 60:220–228.
Yang et al.: Mineralization of PCP by Two Microbial Populations 89
Biotechnology and Bioengineering

Hoelen TP, Reinhard M. 2004. Complete biological dehalogenation of
chlorinated ethylenes in sulfate containing groundwater. Biodegrada-
tion 15:395–403.
Huang GL, Xiao H, Chi J, Shiu WY, Mackay D. 2000. Effects of pH on the
aqueous solubility of selected chlorinated phenols. J Chem Eng Data
45:411–414.
Kamshwaran SR, Crawford D. 2001. Anaerobic biodegradation of penta-
chlorophenol in mixtures containing cadmium by two physiologically
distinct microbial enrichment cultures. J Ind Microbiol Biotechnol 27:
11–17.
Kazumi J, Haggblom MM, Young LY. 1995. Degradation of monochlori-
nated and nonchlorinated aromatic compounds under iron-reducing
conditions. Appl Environ Microbiol 61:4069–4073.
Kennes C, Wu WM, Bhatnagar L, Zeikus JG. 1996. Anaerobic dechlorina-
tion and mineralization of pentachlorophenol and 2,4,6-trichlorophe-
nol by methanogenic pentachlorophenol-degrading granules. Appl
Microbiol Biotechnol 44:801–806.
Kohring GW, Zhang XM, Wiegel J. 1989. Anaerobic dechlorination of 2, 4-
dichlorophenol in freshwater sediments in the presence of sulfate. Appl
Environ Microbiol 55:2735–2737.
Kuwatsuka S, Igarashi M. 1975. Degradation of PCP in soil II. The relation-
ship between the degradation of PCP and the properties of soils and the
identification of the degradation products of PCP. Soil Sci Plant Nutr
21:405–414.
Liamlean W, Annachhatre AP. 2007. Electron donors for biological sulfate
reduction. Biotechnol Adv 25:452–463.
Lovley DR. 1988. Hydrogen concentrations as an indicator of the pre-
dominant terminal electron-accepting reactions in aquatic sediments.
Geochim Cosmochim Acta 52:2993–3003.
Lovley DR, Lonergan DJ. 1990. Anaerobic oxidation of toluene, phenol, and
p-cresol by the dissimilatory iron-reducing organism, GS-15. Appl
Environ Microbiol 56:1858–1864.
Madsen T, Aamand J. 1991. Effects of sulfuroxy anions on degradation of
pentachlorophenol by a methanogenic enrichment culture. Appl
Environ Microbiol 57:2453–2458.
Madsen T, Licht D. 1992. Isolation and characterization of an anaerobic
chlorophenol-transforming bacterium. Appl Environ Microbiol 58:
2874–2878.
Master ER, Lai VW, Kuipers B, Cullen WR, Mohn WW. 2002. Sequential
anaerobic–aerobic treatment of soil contaminated with weathered
Aroclor 1260. Environ Sci Technol 36:100–103.
Mazur CS, Jones WJ, Tebes-Stevens C. 2003. H2 consumption during the
microbial reductive dehalogenation of chlorinated phenols and tetra-
chloroethene. Biodegradation 14:285–295.
McAllister KA, Lee H, Trevors JT. 1996. Microbial degradation of penta-
chlorophenol. Biodegradation 7:1–40.
90 Biotechnology and Bioengineering, Vol. 102, No. 1, January 1, 2009
Mikesell MD, Boyd SA. 1986. Complete reductive dechlorination and
mineralization of pentachlorophenol by anaerobic microorganisms.
Appl Environ Microbiol 52:861–865.
Mohn WW, Kennedy KJ. 1992. Reductive dehalogenation of chlorophenols
by Desulfomonile tiedjei DCB-1. Appl Environ Microbiol 58:1367–1370.
Qiu YL, Hanada S, Ohashi A, Harada H, Kamagata Y, Sekiguchi Y. 2008.
Syntrophorhabdus aromaticivorans gen. nov., sp. nov., the first cul-
tured anaerobe capable of degrading phenol into acetate in obligate
syntrophic associations with a hydrogenotrophic methanogen. Appl
Environ Microbiol 74:2051–2058.
Saber DL, Crawford RL. 1985. Isolation and characterization of Flavobac-
terium strains that degrade pentachlorophenol. Appl Environ Micro-
biol 50:1512–1518.
Seech AG, Trevors JT, Bulman TL. 1991. Biodegradation of pentachlor-
ophenol in soil: The response to physical, chemical, and biological
treatments. Can J Microbiol 37:440–444.
Sekiguchi Y, Kamagata Y, Nakamura K, Ohashi A, Harada H. 1999.
Fluorescence in situ hybridization using 16S rRNA-targeted oligonu-
cleotides reveals localization of methanogens and selected uncultured
bacteria in mesophilic and thermophilic sludge granules. Appl Micro-
biol Biotechnol 65:1280–1288.
Thauer RK, Jungermann K, Decker K. 1977. Energy conservation in
chemotrophic anaerobic bacteria. Bacteriol Rev 41:100–180.
Tschech A, Fuchs G. 1987. Anaerobic degradation of phenol by pure
cultures of newly isolated denitrifying pseudomonads. Arch Microbiol
148:213–217.
van der Zee FP, Villaverde S. 2005. Combined anaerobic–aerobic treatment
of azo dyes—A short review of bioreactor studies. Water Res 39:1425–
1440.
Watanabe I. 1973. Isolation of pentachlorophenol decomposing bacteria
from soil. Soil Sci Plant Nutr 19:109–116.
Wilson GJ, Khodadoust AP, Suidan MT, Brenner RC, Acheson CM. 1998.
Anaerobic/aerobic biodegradation of pentachlorophenol using GAC
fluidized bed reactors: Optimization of the empty bed contact time.
Water Sci Technol 38:9–17.
Wu WM, Bhatnagar L, Zeikus G. 1993. Performance of anaerobic granules
for degradation of pentachlorophenol. Appl Environ Microbiol
59:389–397.
Yang SY, Shibata A, Yoshida N, Katayama A. 2007. Anaerobic degradation
of phenol by enrichment cultures under sulfate- and iron-reducing
conditions. Proceedings of the International Symposium on Ecotopia
Science 2007, ISETS07.
Yoshida N, Yoshida Y, Handa Y, Kim HK, Ichihara S, Katayama A. 2007.
Polyphasic characterization of a PCP-to-phenol dechlorinating micro-
bial community enriched from paddy soil. Sci Total Environ 381:233–
242.