an enzyme immunoassay for dengue antibody using infected cultured mosquito cells as antigen
TRANSCRIPT

Journul of ~/irolo~ic~zl &Whods, 17 (1987) 19 l-198
Elsevier
JVM 00609
191
An enzyme immunoassay for dengue antibody using infected cultured mosquito cells as antigen
Luiz Tadeu M. Figueiredo and Robert E. Shope Yale Arbovirus Research Unit, Department of Epidemiology and Pubiic Health, Yale University
School of Medicine, New Haven, Connecticut, U.S.A.
(Accepted 31 March 1987)
Summary
A simple enzyme immunoassay (EIA) for dengue virus antibody detection used infected tissue culture cells as antigen. C6136 Aedes alhupict~ls cells infected with dengue virus. and uninfected control cells were fixed with 3.3% formalin. This technique eliminated microplate coating and laborious antigen preparation, and it facilitated rapid screening of large numbers of sera. Formalin also inactivated den-
gue virus infectivity. Microplates prepared by this technique could be stored at -20 or -70°C for at least two months. Human sera were adsorbed with C6136 cells prior to testing in the EIA in order to reduce nonspecific binding to C6/36 cells.
Dengue; Enzyme immunoassay; Mosquito cell -
Introduction
The four mosquito-borne dengue virus (family Falviviridae) serotypes cause a spectrum of human disease ranging from classic dengue to dengue hemorrhagic fe- ver with shock syndrome (Halstead, 1980). Dengue viruses infect people through- out the tropics and are a leading cause of hospitalization and death among chil- dren in tropicat countries (WHO, 1985). Rapid and accurate serological diagnosis is fundamental for dengue surveillance programs (Halstead et al., 1983).
Classic serologic techniques include the hemagglutination-inhibition test (Clarke and Casals, 1958) for rapid flavivirus antibody screening and the plaque reduction neutralization test for specific serotype determination (Russell et al., 1967). En-
Correspondence to: L.T.M. Figueiredo. Yale Arbovirus Research tlnit, Department of Epidemiology and Public Health, Yale University School of Medicine. New Haven. CT 06510, U.S.A.

192
zyme immunoassays (EIAs) for dengue antibody detection have been developed during the last 5 yr (Bundo and Igarashi, 1985; Burke, 1983). EIA is inexpensive, quick and simple to perform (Aaskov, 1983). It has many of the properties needed for a good screening test including broad cross-reactivity and high sensitivity. Sev- eral EIAs for arbovirus antibody detection have been described. In the double an- tibody sandwich EIA, viral antigen is captured by affinity-purified antibodies ad- sorbed to the solid phase; human serum is later added and binding of antibodies is detected by an enzyme-linked anti-human conjugate (Niklasson et al., 1984). Other methods include an assay using a solid phase coated with anti-human k chain immunoglobulins which captures human IgM, followed by viral antigen and indi- cator antibodies (Burke, 1983; Niklasson et al., 1984; Monath et al., 1984), and a direct antibody assay using microplates coated with gradient-purified virus (Oseni et al., 1983).
We describe here a simple technique of dengue antibody detection using dengue virus-infected cultured cells as antigen. For this we have adapted EIAs proposed recently for antigen detection of other viruses in cell cultures (Anderson and Rowe, 1982; Smith, 1985; Zhang et al., 1984). The use of infected cultured cells avoids laborious and expensive virus purification and the need for an antigen capture step.
Materials and Methods
Virus strains
Dengue-1 virus strain 1413 was obtained from D.J. Gubler. It was isolated from human serum in Haiti in 1983 and was passed twice in mosquitoes and once in C6/36 Aedes albopictus cells. Dengue-2 virus strain 1232 was obtained from L. Rosen. It was isolated from human serum in Indonesia in 1978 and was passed twice in mos- quitoes and once in C6136 cells.
Antisera
Dengue-1 and dengue-2 hyperimmune mouse immune ascitic fluids were pre- pared using prototype mouse-adapted viruses foilowing the method of Brandt et al. (1967).
Human serf2
The following sera were used: (a) from a person 10 years post-dengue infection plus vaccinations for yellow fever and Japanese encephalitis; (b) from a person two or more years after dengue-2 infection; (c) from a yellow fever convalescent pa- tient; (d) (e) and (f) from persons after inoculation with 17D yellow fever vaccine; (g) (h) and (i) were pooled sera obtained in 1982 from 40 residents of Cuba fol- lowing epidemics of dengue-1 and dengue-2; and (j) from a North American res- ident with no known flavivirus exposure. Sera were obtained by venipuncture and kept at minus 20°C.

193
Cell cultures
CER, LLC-MK2, and Vero cells were grown in minimal essential medium (MEM) containing equal parts of Hanks’ and Earle’s salts (Grand Island Biolog- ical Co. (Gibco), Grand Island, NY). 1% L-glutamine, 10% fetal bovine serum (FBS), 100 U/ml penicillin and 100 kg/ml streptomycin. BHK-21 cells were grown in Glasgow MEM (Gibco) containing 10% FBS, 10% tryptose phosphate broth (TPB), 100 U/ml penicillin, and 100 pgiml streptomycin. These vertebrate cell lines were maintained at 37°C in 5% COZ humidified atmosphere.
C6’36 A&es uibu~icrl~s cells (Igarashi, 1978) were grown in Leibowitz L15 me- dium (Gibco) containing 10% heat-inactivated FBS, 10% TPB, 100 Uiml penicil- lin and 100 pgiml streptomycin, and were maintained at 30°C in a humidified at- mosphere.
Viral injection and aiztigen processing
Suspensions of CER, Vero, LLC-MK2, or BHK-22 cells were added to 96well microplates (Corning No. 25860. Corning, NY) in 150 ~1 at a density of 10” cells per well. C6136 cells were plated at a density of 2x 10’ cells per well. After 24 h the cells were infected either with logi,, dilutions of dengue viruses or with 100 TCII.& of virus. Alternate columns of the microplate wells contained cells which were uninfected. After suitable incubation periods, (usually 4 days) the wells, each containing 200 1.~1, received 100 ~1 of neutral buffered formalin (NBF), pH 7 (37-40% formaldehyde 100 ml, sodium phosphate dibasic-anhydrous 6.5 g, so- dium phosphate monobasic 4.0 g, and distilled water 900 ml (Smith, 1985)) and were held overnight at 4°C. Within 18-24 h of fixation, the cells were washed twice with phosphate buffered saline (PBS). Some microplates were processed imme- diately for EIA. Others were air dried, put in plastic bags and sealed in a nitrogen atmosphere. The microplates containing infected and non-infected fixed cells were stored at 25, 4, -20, or -70°C.
EfA
The EIA was performed as described by Smith (1985) with some modifications. Cells were saturated with 200 ~1 of blocking buffers (0.5% bovine serum albumin (BSA) in PBS; 0.05% Tween 20 in PBS; 5% horse serum in Tween-PBS; or 3% gelatin in PBS). A comparison of the four blocking buffers used in combination with dengue-1 virus-infected and uninfected C6136 cells showed that none of the blockers was significantly better than any other, and 3% gelatin in PBS was adopted for use. After 1 h at 37”C, antiserum diluted in PBS containing 0.5% BSA was added for 1 h. Horseradish peroxidase-conjugated goat anti-mouse IgG or goat anti- human immunoglobulin (diluted 1:2000; Tago, Inc., Burlingame, CA) in 0.5% BSA-PBS was added for 1 h at 37°C. Results were read visually and spectropho- tometrically (414 nm) 20-30 min after the addition of ABTS substrate (Kirkegaard and Perry, Gaithersburg, MD) to wells that had been washed five times with 0.0570 Tween 20 in PBS.

194
Calculations
Optical density (OD) values were considered significant if they exceeded by 3
SD the mean value for control wells in the same assay. Virus titers were calculated by the method of Reed and Muench (1938).
Adsorption of human serum with C6136 cells
One hundred ~1 of human sera were diluted 1:lO in 0.5% BSA-PBS and ad- sorbed at 4°C overnight with previously pelleted, washed C6136 cells.
Tests of virus inactivation
C6/36 cells growing in 25 cm’ flasks were infected with 1000 TCID5,, of dengue- 1 or dengue-2 virus. Formalin was added to the flasks 6 days after virus inoculation (final concentration 3.3%). A similar volume of PBS was added to control flasks. After 18 h at 4”C, the cells were washed, disrupted by one cycle of freezing and thawing and centrifuged at 10000 x g for 10 min. Fifty ~1 of supernatants and log,,, dilutions were used to infect C6/36 cell cultures growing in microplates. The cells were processed 6 days later by EIA.
Results
To determine which cell line was most permissive to dengue virus, CER, BHK- 21, Vero, C6136, and LLC-MK2 cells in microplates were exposed to log,,, dilu- tions of dengue-2 virus. EIAs were performed six days post-infection. Dengue-2 virus titers were significantly higher in C6136 cells than in the other cell lines (C6136, 5.7 log,,/ml; BHK-21, 3.5 log,,/ml; Vero, 2.8 log,,,/ml). CER and LLC-MK2 cells yielded no reaction product. All cell monolayers adhered to the plastic wells. Based
TABLE 1
Development of dengue 1 virus in C6136 cells as determined by EIA with dengue 1 mouse immune
ascitic fluid.
Days post-inoculation Virus titers (log,,,/ml)
1 1.8
2 <2.3
3 3.6
4 4.7
5 5.2
6 5.9 7 6.0
8 6.3
9 5.9 10 5.9
195
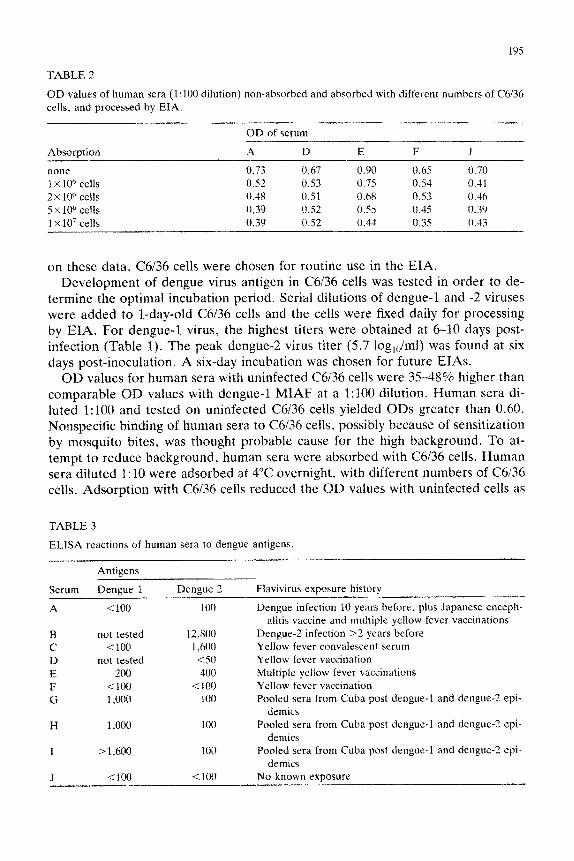
TABLE 2
OD values of human sera (1:lOO dilution) non-absorbed and absorbed with different numbers of C6136
cells, and processed by EIA.
Absorption
none
1 x 10” cells
2x 106 cells
5 x 10” cells
1 X 107 cells
OD of serum
A D E F J
0.73 0.67 0.90 0.65 0.70 0.52 0.53 0.75 0.54 0.41 0.48 0.51 0.68 0.53 0.46
0.39 0.52 0.55 0.45 0.39 0.39 0.52 0.34 0.35 0.43
on these data, 05136 cells were chosen for routine use in the EIA. Development of dengue virus antigen in C6136 cells was tested in order to de-
termine the optimal incubation period. Serial dilutions of dengue-1 and -2 viruses were added to l-day-old C6/36 cells and the cells were fixed daily for processing by EIA. For denguc-1 virus, the highest titers were obtained at 6-30 days post- infection (Table 1). The peak dengue-2 virus titer (5.7 log,,,/ml) was found at six days post-inoculation. A six-day incubation was chosen for future EIAs.
OD values for human sera with uninfected C6/36 cells were 3548% higher than comparable OD values with dengue-1 MIAF at a 1:lOO dilution. Human sera di- luted 1:lOO and tested on uninfected C6136 cells yielded ODs greater than 0.60. Nonspecific binding of human sera to C6/36 cells, possibly because of sensitization by mosquito bites, was thought probable cause for the high background. To at- tempt to reduce background, human sera were absorbed with C6136 cells. Human sera diluted 1:lO were adsorbed at 4°C overnight, with different numbers of C6136 cells. Adsorption with C6136 cells reduced the OD values with uninfected cells as
TABLE 3
ELlSA reactions of human sera to dengue antigens.
Antigens
Serum Deneue 1 Dengue 2 Flavivirus exposure history
A <lOO 100 Dengue infection 10 years before, plus Japanese enceph- alitis vaccine and multiple yellow fever vaccinations
B not tested 12,800 Dengue-2 infection >2 years before
C <lOO 1,600 Yellow fever convalescent serum
D not tested <SO Yellow fever vacfination
E 200 4OU Multiple yellow fever vaccinations
F <loo <l(K) Yellow fever vaccination
G 1,000 100 Pooled sera from Cuba post dengue-1 and dengue-2 epi-
demics
H 1,000 100 Pooled sera from Cuba post denguc-1 and dengue-2 epi-
demics
r >1,600 100 Pooled sera from Cuba post dengue-I and dengue-2 epi-
demics
J <IDO <lOtI No known exposure

shown in Table 2. The effect seemed optimal at 5X 10” cells (46% reduction for serum A).
Ten adsorbed human sera with reduced background were tested with dengue-1 and dengue-2 infected C6136 cells. Sera E (1:200 titer), G (1: 1000 titer), H (1: 1000 titer), and I (1:1600 or greater titer) were positive for dengue-1 antibodies and sera A (1:lOO titer), B (1:12800 titer), C (1:1600 titer), E (1:400 titer), and G, H, and I (1:lOO titer) were positive for dengue-2 antibodies (Table 3).
To determine whether dengue antigen in the form of infected C6i36 cells was stable, plates were stored at different temperatures after formalin fixation. An EIA was performed with cells immediately after fixation and with cells after 2 mth stor- age at varying temperatures. The dengue-1 MIAF titer on dengue virus-infected C6136 cells was 1:6400 on fresh cells. The MIAF titer decreased greater than 32- fold when tested on cells stored for 2 mth at room temperature (titer 1:200) and 4°C (titer <l:lOO). The MIAF titer did not change significantly in assays of cells held for 2 mth at -20°C and -70°C (titers 1:12800).
Infectivity was not detected in cell lysates derived from formalin-treated cells. Lysates of PBS-treated cells yielded 1O3.’ TCIDS,,/ml (dengue-1 virus) and 10s.’ TCID,,,/ml (dengue-2 virus),
Discussion
EIAs are sensitive tests based, in part, on the amplification factor inherent in the nature of enzyme-substrate reactions (Kurstak et al., 1984). The EIA de- scribed here using infected cultured cells for dengue antibody detection has many advantages over conventional antibody capture EIAs and other tests. It eliminates solid phase coating with dengue virus and laborious antigen preparation; it permits screening of large numbers of sera faster and more easily than by HI or PRNT; 3.3% formalin inactivates viral infectivity, thereby reducing the potential hazard of laboratory infection; C6136 cells infected in microplates with dengue virus and fixed with formalin can be stored for at least 2 mth at -20 or -70°C and also can be transported for use in remote dengue virus endemic or epidemic areas.
The finding of higher dengue-1 virus titers in C6136 cells than in other cell lines is consistent with results of other studies of dengue virus infection of this cell line (fgarashi, 1978; Tesh, 1979). C6136 cells have advantages over other cell lines: they grow rapidly, can be nlaintain~d in the tropics without an incubator, and contain a high percentage of antigen-positive cells in spite of no evident cytopathic effect (Tesh, 1979).
Non-specific reactions have been associated with many antibody detection tech- niques. Non-specific inhibitors and natural agglutinins are present in human sera and must be removed before the HI test (Clarke and Casals, 1958). The presence of rheumatoid factor or antinuclear antibody in high concentration can be the cause of EIA false positives (Katchaki et al., 1986). Elevated OD values for wells con- taining uninfected C6136 cells and reacted with human sera were a problem in the dengue virus EIA. The background reduction associated with adsorption of hu-

197
man sera with C6136 cells was satisfactory although the adsorption procedure added another step to the assay.
The EIA detected dengue-1 antibodies present in sera from the three pools of Cuban dengue virus-infected workers. The EIA also detected dengue-2 antibodies in sera from two dengue-2 virus-infected patients, one yellow fever convalescent patient and one person who had received multiple yellow fever vaccinations. Al- though these experiments were not designed to test the specificity of the EIA, it is clear that the test is cross-reactive among flavivirus antibodies. Dengue-1 anti- bodies were not detected in sera from a person without antecedent flavivirus in- fection, or from one of two yellow fever vaccinees. The detection of dengue-2 an- tibody at a high dilution (1:12 800) of human serum B suggests that the EIA is a very sensitive test. The test is applicable as a rapid screening procedure for flavi- virus antibody in epidemiologic studies.
Further studies with this EIA technique are needed to optimize the reduction of non-specific binding of human sera to C6136 cells, to adapt the test for human serum IgM detection, and to get more information about the specificity of antibody in human sera with dengue viruses and other flaviviruses.
Acknowledgements
This work was supported by the Brazilian Council for Scientific and Technology Development (CNPq) award no. 20.0035185, by the World Health Organization, by grant AI 10984 of the U.S. National Institutes of Health, and by grant DAMD17-87-G-7005 from the U.S. Army Medical Research and Development Command. We thank L. Rosen, D. Gubler, J. Olson, and N. Marchette for sup-
plying sera and viruses.
References
Aaskov, J. (1983) In: Proc. Int. Conf. DengueiDengue haemorrhagic fever. (Pang. T. and Pathman-
athan, R., eds.), p. 403. University of Malaya, Kuala Lumpur.
Anderson, J. and Rowe, L.W. (1982) J. Immunol. Methods 53, 183.
Barman, T.E. (1974) Enzyme Handbook, Vol. 2. Springer-Verlag. Berlin and New York.
Brandt, W.E.. Buescher, E.L. and Hetrick, F.M. (1967) Am J. Trop. Med. Hyg. 16. 333.
Bundo, K. and Igarashi, A. (1985) J. Virol. Methods 11, 15.
Burke, D.S. (1983) In: Proc. Int. Conf. DengueiDengue haemorrhagic fever. (Pang, T. and Pathman-
athan, R., eds.), p. 72. University of Malaya, Kuala Lumpur. Clarke, D.H. and Casals, J. (1958) Am J. Trop. Med. Hyg. 7, 561.
Halstead, S.B. (1980) In: The Togaviruses, (Schlesinger, R.W., ed.), p. 107. Academic Press, New
York.
Halstead, S.B., Rojanasuphot, S. and Sangkawibha, N. (1983) Am. J. Trop. Med. Hyg. 32, 154. Igarashi. A. (1978) J. Gen. Viral. 40, 531.
Katchaki, J.N.. Siem, T.H., Brower, R. and Nieste, H.L.J. (1986) Lancet i, 448.
Kurstak, E., Tijssen, P. and Kurstak, C. (1984) Applied Virology, (Kurstak, E., ed.), p. 479. Aca- demic Press. New York.

198
Monath, T.P., Nystrom. R.R.. Bailey, R.E.. C&her. C.H. and Muth. D.J. (1984) J. Clin. Microbial. 20. 7x4.
Niklasson, 8.. Peters, C.J.. Grandien. M. and Wood. 0. (1984) J. Clin. Microbial. 19. 225. Oseni, R.A., Donaldson. M.D.. Dalglish, D.A. and Aaskov. J.G. (1983) Bull. W.H.O. hi. 703. Reed, L.J. and Muench. H. (1938) Am. J. Hyg. 27. 493. Russell, P.K.. Nisalak, A.. Sukhayachana. P. and Vivona, S. (1967) J. Immunol. 99, 2X.5. Smith, A.L. (1985) J. Viral. Methods II, 321. Tesh, R.B. (1979) Am. J. Trap. Med. Hyg. 28, 1053. World Health ~rgan~~~tion (1985) Arthropod-borne and rodent-borne viral diseases. Report of a WHO
Scientific Group. Technical Report Series 710 World Health Organization, Geneva. Zhang. Y.-H.. Yu, W.-F.. Tian. Z.-W., Ge, J.-O., Chen, O.-S. and Wang, Y.-M. (1984) J. Viral.
Methods 9, 45.