alternative mating tactics in the great plains toad
TRANSCRIPT

Anim. Behav., 1989,37, 1035-1043
Alternative mating tactics in the Great Plains toad
JAMES J. KRUPA* Department of Zoology and the Oklahoma Biological Survey, University of Oklahoma, Norman,
Oklahoma 73019, U.S.A.
Abstract. Male Great Plains toads, Bufo cognatus, exhibit two mating tactics. At any time, most males give advertisement calls to attract females, while other males, ‘satellites’, remain silent and station themselves near callers in an attempt to intercept females. Females are usually capable of moving through choruses undetected by males. Those females detected by males can avoid being clasped by quickly darting away; but if clasped, can detach the male by inflating. Females initiated amplexus with callers; subsequently, calling males mated with 92% of the females and satellites mated with the remaining 8%. Toads employing the satellite tactic associated with males that had longer calls and that were larger, although call duration was the stronger predictor. The percentage of males exhibiting satellite behaviour during a sampling period varied from 0 to 57%. Several hypotheses explaining satellite frequency were explored, including variation in male density, male quality, and operational sex ratio. Of the variables considered, only two aspects of male density (namely, the number of unmated males per m of shoreline and number of calling males per m shoreline) affected satellite frequency. High male density may cause at least some calling males to lose vocal conspicuousness, thereby favouring satellite behaviour. However, male density accounted for only 17% of the observed variation in satellite frequency. This, plus the fact that up to 74% of marked males switched between tactics, suggests that factors such as higher energetic demands of calling may affect satellite frequency.
Many examples of intraspecific variation in male mating tactics are known for anuran amphibians (reviewed by Wells 1977; Waltz 1982; Arak 1983a; Perrill & Magier 1988). In this group the predomin- ant mating tactic consists of advertisement calls given by stationary males for attracting females. Non-calling tactics include satellite behaviour (males that remain silently near callers; Wells 1977; Howard 1978; Arak 1983b) and males that actively search the breeding sites for single females (e.g. Davies & Halliday 1979; Fairchild 1984).
Calling tactics result in a higher probability of mating than achieved by satellites in most of the species studied to date (Howard 1978; Fellers 1979; Gatz 1981; Sullivan 1982a; Arak 1983a; Robertson 1986; Arak 1988). Alternative mating tactics are thought to have evolved because they are the best reproductive option for disadvantaged males (Dawkins 1980; Waltz 1982; Waltz & Wolf 1984). For example, if large males are more attractive to females or only certain subareas on a breeding site permit high egg survival, then small males may profit from adopting the non-calling tactic and positioning themselves in an area of high female * Present address: School of Biological Sciences, Univer-
sity of Kentucky, Lexington, KY 40506-0225, U.S.A.
traffic. Satellite males may be young and unable to hold a quality site or individuals that are relatively unattractive. In either situation, satellites are pre- dicted to associate with the areas of males that are most attractive to females in order to improve their chances of intercepting females or of acquiring vacated territories (Waltz 1982).
In contrast to age or morphological constraints, males of some species exhibit behavioural flexibi- lity by switching between tactics on a short-term basis, even though the non-calling tactic results in lower mating success (Fellers 1979; Gatz 1981; Sullivan 1982b; Fairchild 1984; Robertson 1986; Arak 1988). The phenomenon of switching sug- gests that each tactic may have costs and benefits that vary with changing conditions.
A number of factors are thought to influence the frequency of alternative tactics. Male density is considered a positive correlate of satellite fre- quency (Alexander 1975; Wells 1977; Arak 1983a; Fairchild 1984). This prediction is based on the assumption that increased male density will be associated with a decrease in benefits and an increase in either competition for mates or cost of defending call sites, which should cause males at a competitive disadvantage to switch to a cheaper,
0003-3472/89/061035 +09 $03.00/O 0 1989 The Association for the Study of Animal Behaviour
1035

1036 Animal Behaviour, 37, 6
albeit less successful, tactic. In other words, those males least likely to mate should minimize their costs by being satellites.
A second factor that has been predicted to influence satellite frequency is variation in site or male quality (Waltz 1982). As variance or disparity in male quality increases, the number of satellites should also increase. This prediction is based on the assumption that satellites will adopt the non- calling tactic because they are not successful.
The third factor is that the availability of females should affect satellite frequency. Female avail- ability is measured as operational sex ratio (the ratio of the number of receptive females to the number of breeding males at a given time; Emlen & Oring 1977). Waltz (1982) and Fairchild (1984) provided contrasting predictions on how oper- ational sex ratio will influence satellite frequency. Waltz (1982) argued that all males have a higher probability of mating when females swamp the breeding site by arriving synchronously. Swamping in turn reduces intrasexual competition. This should prompt males to avoid the more costly calling tactic which is advantageous when competi- tion is high. By contrast, Fairchild (1984) predicted that a scarcity of females will result in a higher frequency of the alternative tactic. He reasoned that the more mobile non-callers can improve their probability of encountering relatively rare females. Conversely, callers are more likely to attract females (without the females being intercepted by non-callers) when females are more abundant. Thus Waltz (1982) predicts that as operational sex ratio becomes less male-biased, satellite frequency will increase, while Fairchild (1984) predicts that as operational sex ratio becomes more male-biased, satellite frequency will increase.
Bufo cognates is a rain-stimulated, explosive breeding anuran (Wells 1977) that breeds in tem- porary, rain-filled pools. Breeding can occur from late March to early July in Oklahoma. The pres- ence of non-calling males at breeding sites is well documented for this species: non-callers were reported for grassland populations in Oklahoma (Bragg 1936, 1937, 1940) and for desert popula- tions in Arizona (Brown & Pierce 1967; Sullivan 1982b, 1983a) and New Mexico (Sullivan 1982b, 1983a). Sullivan (1982b) labelled such non-callers as satellites and reported that, on average, they comprise 25% (range, O-75%) of the males per chorus. Females usually initiate amplexus with calling males; subsequently, callers acquire most of
the matings (Sullivan 1982b, 1983a). Furthermore, males that acquire matings tend to be those that had previously spent more time calling than unsuc- cessful males (77 versus 64%: Sullivan 1983a). Satellite frequency was thought to depend on male density (Brown & Pierce 1967) which Sullivan (1982b) corroborated. Specifically, Sullivan found a significant positive relationship between satellite frequency and the number of males per metre of shoreline.
The present study was conducted on the Great Plains toad, Bufo cognatus, and the purpose was three-fold. (1) To test the following predictions on satellite frequency: (a) prediction 1: satellite fre- quency increases as male density increases (Alex- ander 1975; Wells 1977; Arak 1983a; Fairchild 1984); (b) prediction 2: satellite frequency increases as the range in male ‘quality’ increases (Waltz 1982) thus some males should be more likely to attract satellites; and (c) prediction 3: satellite frequency is influenced either positively or nega- tively by operational sex ratio (Waltz 1982; Fair- child 1984). (2) To provide observations on the general behaviour of callers and satellites, and the flexibility these males exhibit by switching between tactics. (3) To compare the effectiveness of callers and satellites at acquiring mates.
METHODS
The behaviour of B. cognatus was observed in Norman, Oklahoma during the 1984 (April through June), 1985 (March through June) and 1986 (May) breeding seasons.
Twenty-six breeding sites were visited in 1984 and 1985. I conducted observations during the breeding bouts (periods of one to nine nights when toads mated) that followed rainstorms. Breeding activity did not occur at each site after every rain, resulting in opportunistic sampling of sites; how- ever, no site was sampled more than once during a single breeding bout. Fifty-one breeding choruses (defined as a congregation of breeding toads found in a pool on one night) were sampled during 30 nights over the two seasons.
At the onset of observations, I noted the number of nights since the last rainfall (e.g. number of nights into the breeding bout), then numbered flags were placed next to each toad in a pool. All toads were then quickly gathered and sorted into separ- ate buckets for: (1) calling males associated with

Krupa: Alternative mating tactics 1037
satellites (with satellite defined operationally as a non-calling male that sits within 1 m of a caller); (2) calling males not associated with satellites; and (3) satellites. The pool was then searched carefully for all amplectant pairs and for single females, both of which were similarly collected and held separa- tely. All toads were counted by category, measured for snout-vent length (SVL), and returned to the pool (usually within 15 min of gathering). The following day distances between flags and shoreline of each pool were measured. From this, distance between males and percentage of shoreline used by toads were quantified. The number of breeding males, satellite frequency (with satellite frequency defined as the percentage of all unmated breeding males that were satellites), and the area ofpool used by breeders changed over the course of an evening. As a result, only data from a single observation period of a chorus were used in the analyses. Because the males in most choruses lined the shore, male density data were calculated in terms of Sullivan’s (1982b) measurement (males per m of shoreline), with choruses where males spread over the surface of the pool excluded from these analyses.
Simple regression analyses were used to deter- mine the effects of 10 variables on satellite frequency. These variables were derived from the data collected above. They include: (1) male density (expressed as the number of umated males per m of shoreline); (2) total number of males in the chorus; (3) caller density (number of calling males per m of shoreline); (4) total number of calling males; (5) mean distance between closest calling males; (6) total number of females; (7) operational sex ratio; (8) number of nights after a rainstorm; (9) water temperature of breeding pool; and (10) variance in male SVL within each chorus. Variable 10 was considered a potential estimate of male quality, since females may be attracted more to certain size classes of males or to various call characteristics (Howard 1978; Davies & Halliday 1979; Arak 1983b; Sullivan 1983b) and because dominant frequency and pulse rate are correlated with SVL (Krupa, personal observation).
Twenty-five of the 51 choruses were observed during the 1985 season. Besides the information collected above, each chorus was searched for satellite-caller associations and for solitary callers. In large, dense choruses (where males tended to move frequently) a portion of the males were sampled along a transect or one shore to prevent
encountering the same calling male more than once. For each caller, samples of one to three calls were timed with a stopwatch, recorded with a Uher 4000 tape-recorder, and later analysed on a Uni- scan II audio-spectral analyser. Snout-vent lengths of the males were measured after their calls were recorded. Dominant frequency (Hz) was measured to the nearest 10 Hz and pulse rate (pulses per s) was recorded for each caller. Temperature was not considered a confounding variable on call para- meters. Only pulse rate was influenced by tempera- ture, but body temperatures in this study did not range more than 1.6”C within any chorus (Krupa, personal observation). Measurements were taken from the midpoints of two or three 1.6-s audio- spectrographs of consecutive calls. The means for these were calculated for callers with satellites and solitary callers, and compared within each chorus using the Wilcoxon signed-ranks test (Sokal & Rohlf 1981) and stepwise logistic regression (BMDP: Engelman 1979). Finally, during the 1986 breeding season, continuous recordings were made on 39 males’ calls for a 2-min period. Call duration and intercall interval were measured from Uniscan audio-spectrographs.
During the course of the study, the general behaviour of callers and satellites was recorded. Toads were watched at a breeding site during two breeding bouts. The first lasted three nights (28-30 May 1984); the other, four nights (1417 May 1986). Toads were individually marked with waist bands in 1984 and with pieces of numbered plastic measuring 5 x 5 mm glued to their backs in 1986. I periodically walked around the pool noting which males were callers versus satellites. From these data, the percentages of males that were observed as callers, satellites, or both during the course of a breeding bout could be determined.
The mating success of callers and satellites was determined via female-release observations that were conducted at a single pool during the 1986 breeding season. Observations were made of 13 females (23 female-release trials) during the only four nights of the entire breeding season when females occurred at that site (14-17 May). Paired females were separated from their mates and released at either end of the pool. The following information was then recorded: (1) the number of times females initiated amplexus versus the number of times females were approached and clasped by males; (2) the number of times satellites attempted to intercept females and the number of times they
1038 Animal Beha rviour, 37, 6
succeeded; (3) the number of females that spawned with satellites or callers; and (4) the general be- haviour of the females. In those trials where satellites clasped females, the pair was not pried apart for additional trials. Instead, the pair was checked sporadically during the evening and until morning when spawning began.
RESULTS
Chorus configurations of B. cognatus varied greatly. In 10 choruses, calling males dispersed over the surface of shallow pools; in 41 other choruses, callers were confined to shorelines. The fraction of shoreline used in these 41 choruses varied greatly, with males evenly distributed around the shores of 16 choruses, but aggregated as one or two ‘call groups’ (defined as aggregations of toads separated from other such aggregations by 15 or more m, see Arak 1983b) in 25 choruses. For those choruses where call groups occurred, only 48% of the shoreline was occupied by males (range = 13-93%). In 22 of the 26 pools, vegetation covered 100% of the shoreline and call groups occurred on different stretches on different nights. For the remaining four pools, some of the shoreline appeared unused because of steep banks or tall, dense vegetation.
The 51 choruses contained an average of 17 males (range=4-86 males) and males exhibiting the satellite tactic were found in 41 (82%) of them. On average, 27% (SE= 2.4%; range=O-57%) of unmated males were satellites in these 41 choruses. The mean SVL of satellites was less than that for callers in 31 of 39 choruses (T,=97.0, PcO.01; Wilcoxon signed-ranks test), although males of nearly all sizes were observed to exhibit satellite behaviour (range for satellites = 54-89 mm; range for callers = 60-99 mm). Callers were never within 1.0 m of each other (mean distance=4.7 m, N= 346). By contrast, the mean distance between callers and satellites was 0.37 m (range=O.Ol- 0.87 m, N= 159). Furthermore, stationary and silent males (i.e. those that might be categorized as satellites) were never observed beyond 1 m of a caller. In seven of these 41 linear choruses, every unmated male called. These seven c&oruses had low male densities (less than 0.43 male$m shoreline).
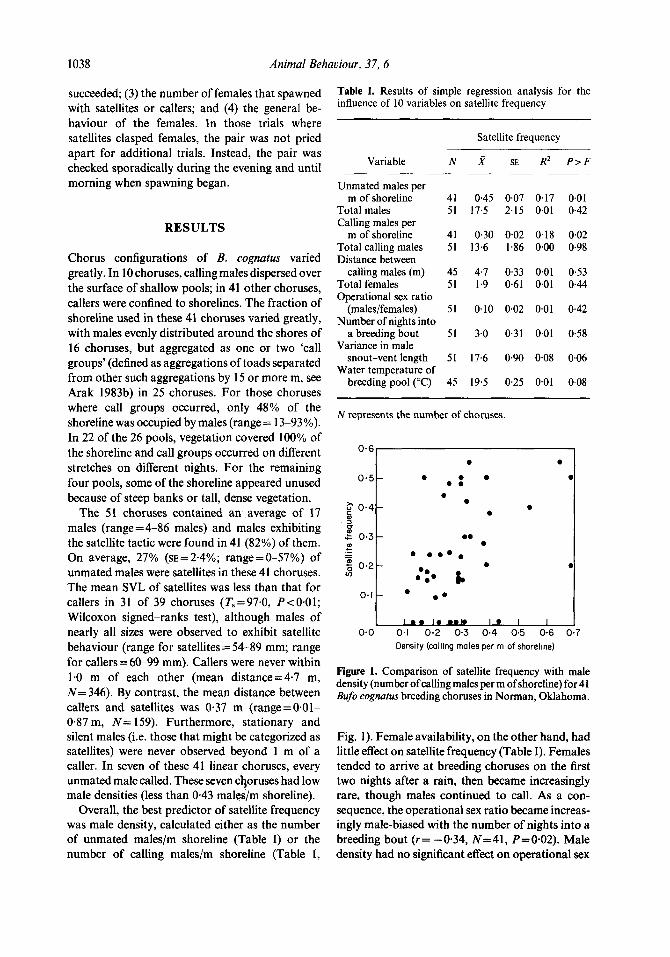
Overall, the best predictor of satellite frequency was male density, calculated either as the number of unmated males/m shoreline (Table I) or the number of calling males/m shoreline (Table I,
Table 1. Results of simple regression analysis for the influence of 10 variables on satellite frequency
Satellite frequency
Variable
Unmated males per m of shoreline
Total males Calling males per
m of shoreline Total calling males Distance between
calling males (m) Total females Operational sex ratio
(males/females) Number of nights into
a breeding bout Variance in male
snout-vent length Water temperature of
breeding pool (“C)
N x SE R2 PrF
41 0.45 0.07 0.17 0.01 51 17.5 2.15 0.01 0.42
41 0.30 0.02 0.18 0.02 51 13.6 1.86 0.00 0.98
45 4.7 0.33 0.01 0.53 51 1.9 0.61 0.01 044
51 0.10 0.02 0.01 0.42
51 3.0 0.31 0.01 0.58
51 17.6 0.90 0.08 0.06
45 19.5 0.25 0.01 0.08
N represents the number of choruses.
1 Im*Imm~I. I. I I I 0.0 0.1 o-2 0.3 0.4 0.5 0.6 0.7
Density (calling males per m of shoreline)
Figure 1. Comparison of satellite frequency with male density (number of calling males per m of shoreline) for 41 Bufo cognatus breeding choruses in Norman, Oklahoma.
Fig. 1). Female availability, on the other hand, had little effect on satellite frequency (Table I). Females tended to arrive at breeding choruses on the first two nights after a rain, then became increasingly rare, though males continued to call. As a con- sequence, the operational sex ratio became increas- ingly male-biased with the number of nights into a breeding bout (r= -0.34, N=41, P=O.O2). Male density had no significant effect on operational sex
Krupa: Alternative mating tactics
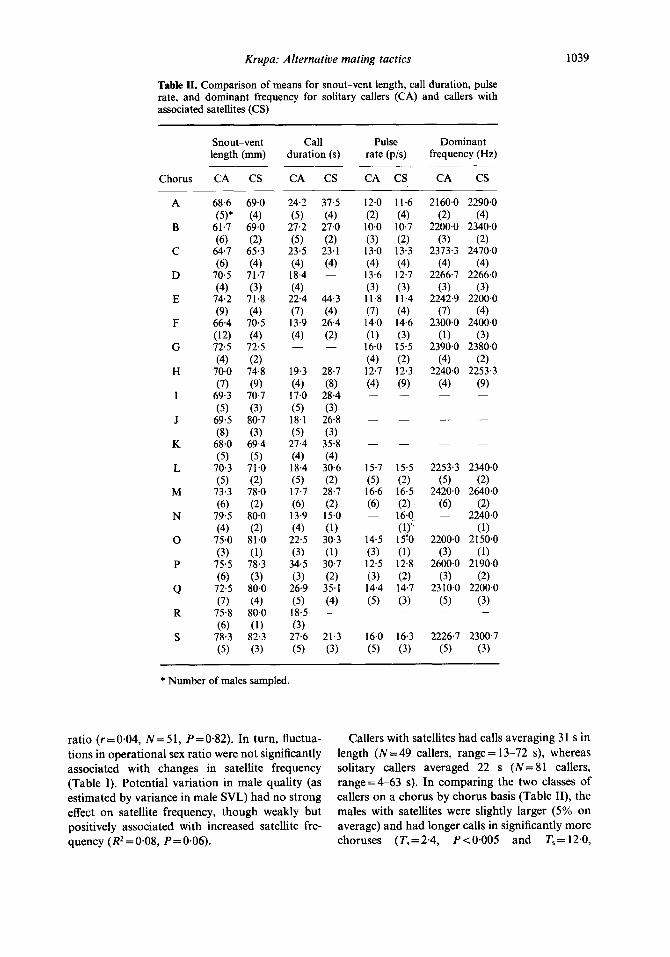
Table II. Comparison of means for snout-vent length, call duration, pulse rate, and dominant frequency for solitary callers (CA) and callers with associated satellites (CS)
1039
Chorus
A
B
C
D
E
F
G
H
I
J
K
L
M
N
0
P
Q
R
s
Snout-vent Call Pulse Dominant length (mm) duration (s) rate (p/s) frequency (Hz)
CA CS CA CS CA CS CA CS
68.6 69.0 (5)* (4)
61.7 69.0 (6) (2)
64.7 65.3 (6) (4)
70.5 71.7 (4) (3)
74.2 71.8 (9) (4)
66.4 70.5 (12) (4) 72.5 72.5
(4) (2) 70.0 74.8
(7) (9) 69.3 70.7
(5) (3) 69.5 80.7 (8) (3)
68.0 69.4 (5) (5)
70.3 71.0 (5) (2)
73.3 78.0 (6) (2)
79.5 80.0 (4) (2)
75.0 81.0 (3) (1)
75.5 78.3 (6) (3)
72.5 80.0 (7) (4)
75.8 80.0 (6) (1)
78.3 82.3 (5) (3)
24.2 37.5 (5) (4)
27.2 27.0 (5) (2)
23.5 23.1 (4) (4)
18.4 - (4)
22.4 44.3 (7) (4)
13.9 26.4 (4) (2)
19.3 28.7 (4) (8)
17.0 28.4 (5) (3)
18.1 26.8 (5) (3)
27.4 35.8 (4) (4)
18.4 30.6 (5) (2)
17.7 28.7 (6) (2)
13.9 15.0 (4) (1)
22.5 30.3 (3) (1)
34.5 30.7 (3) (2)
26.9 35.1 (5) (4)
18.5 - (3)
27.6 21.3 (5) (3)
12.0 11.6 (2) (4)
10.0 10.7 (3) (2)
13.0 13.3 (4) (4)
13.6 12.7 (3) (3)
11.8 11.4 (7) (4)
14.0 14.6 (1) (3)
16.0 15.5 (4) (2)
12.7 12.3 (4) (9)
2 160.0 2290.0 (2) (4)
2200.0 2340.0 (3) (2)
2373.3 2470.0 (4) (4)
2266.7 2266.0 (3) (3)
2242.9 2200.0 (7) (4)
2300.0 2400.0 (1) (3)
2390.0 2380.0 (4) (2)
2240.0 2253.3 (4) (9)
- -
15.7 15.5 (5) (2)
16.6 16.5 (6) (2) - 16.0
w 14.5 15’0 (3) (1)
12.5 12.8 (3) (2)
14.4 14.7 (5) (3)
16.0 16.3 (5) (3)
2253.3 2340.0 (5) (2)
2420.0 2640.0 (6) (2) - 2240.0
(1) 2200.0 2150.0
(3) (1) 2600.0 2190.0
(3) (2) 23 10.0 2200.0
(5) (3)
2226.7 2300.7 (5) (3)
* Number of males sampled.
ratio (r=0.04, N=51, P=O+32). In turn, fluctua- Callers with satellites had calls averaging 31 s in tions in operational sex ratio were not significantly length (N=49 callers, range= 13-72 s), whereas associated with changes in satellite frequency solitary callers averaged 22 s (N=81 callers, (Table I). Potential variation in male quality (as range=463 s). In comparing the two classes of estimated by variance in male SVL) had no strong callers on a chorus by chorus basis (Table II), the effect on satellite frequency, though weakly but males with satellites were slightly larger (5% on positively associated with increased satellite fre- average) and had longer calls in significantly more quency (R* = 0.08, P= 0.06). choruses (T, = 2.4, P~O.005 and T,=12.0,
1040 Animal Behaviour, 37. 6
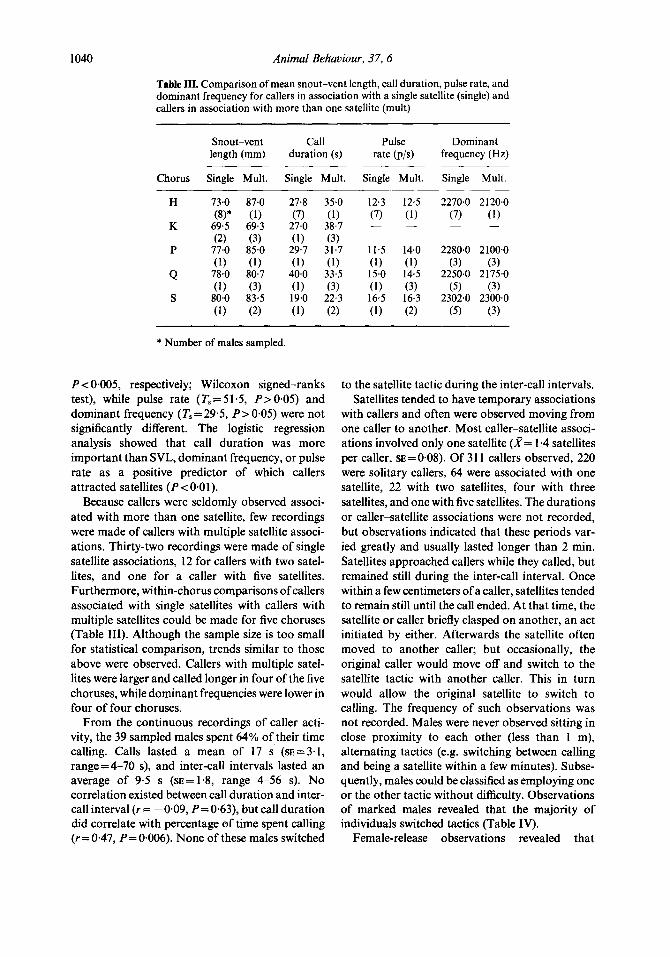
Table III. Comparison of mean snout-vent length, call duration, pulse rate, and dominant frequency for callers in association with a single satellite (single) and callers in association with more than one satellite (mult)
Snout-vent Call Pulse Dominant length (mm) duration (s) rate (p/s) frequency (Hz)
Chorus Single Mult. Single Mult. Single Mult. Single Mult.
H 73.0 (8)’
K 69.5 (2)
P 77.0 (1)
Q 78.0 (1)
S 80.0 (1)
87.0 (1)
69.3 (3)
85.0 (1)
80.7 (3)
83.5 (2)
27.8 350 12.3 12.5 (7) (1) (7) (1)
27.0 38.7 - - (1) (3)
29.7 31.7 11.5 14.0 (1) (1) (1) (1)
40.0 33.5 15.0 14.5 (1) (3) (1) (3)
19.0 22.3 16.5 16.3 (1) (2) (1) (2)
2270.0 2120.0 (7) (1)
2280.0 2100.0 (3) (3)
2250.0 2175.0 (5) (3)
2302.0 2300.0 (5) (3)
* Number of males sampled.
P < 0.005, respectively; Wilcoxon signed-ranks test), while pulse rate (T,=51.5, PpO.05) and dominant frequency (Ts = 29.5, P > 0.05) were not significantly different. The logistic regression analysis showed that call duration was more important than SVL, dominant frequency, or pulse rate as a positive predictor of which callers attracted satellites (P < 0.01).
Because callers were seldomly observed associ- ated with more than one satellite, few recordings were made of callers with multiple satellite associ- ations. Thirty-two recordings were made of single satellite associations, 12 for callers with two satel- lites, and one for a caller with five satellites. Furthermore, within-chorus comparisons of callers associated with single satellites with callers with multiple satellites could be made for five choruses (Table III). Although the sample size is too small for statistical comparison, trends similar to those above were observed. Callers with multiple satel- lites were larger and called longer in four of the five choruses, while dominant frequencies were lower in four of four choruses.
From the continuous recordings of caller acti- vity, the 39 sampled males spent 64% of their time calling. Calls lasted a mean of 17 s (SE = 3.1, range=470 s), and inter-call intervals lasted an average of 9.5 s (SE= 1.8, range 4-56 s). No correlation existed between call duration and inter- call interval (r = -0.09, P= 0.63), but call duration did correlate with percentage of time spent calling (r = 0.47, P= 0.006). None of these males switched
to the satellite tactic during the inter-call intervals. Satellites tended to have temporary associations
with callers and often were observed moving from one caller to another. Most caller-satellite associ- ations involved only one satellite (.X?= 1.4 satellites per caller, SE= 0.08). Of 311 callers observed, 220 were solitary callers, 64 were associated with one satellite, 22 with two satellites, four with three satellites, and one with five satellites. The durations or caller-satellite associations were not recorded, but observations indicated that these periods var- ied greatly and usually lasted longer than 2 min. Satellites approached callers while they called, but remained still during the inter-call interval. Once within a few centimeters of a caller, satellites tended to remain still until the call ended. At that time, the satellite or caller briefly clasped on another, an act initiated by either. Afterwards the satellite often moved to another caller; but occasionally, the original caller would move off and switch to the satellite tactic with another caller. This in turn would allow the original satellite to switch to calling. The frequency of such observations was not recorded. Males were never observed sitting in close proximity to each other (less than 1 m), alternating tactics (e.g. switching between calling and being a satellite within a few minutes). Subse- quently, males could be classified as employing one or the other tactic without difficulty. Observations of marked males revealed that the majority of individuals switched tactics (Table IV).
Female-release observations revealed that

Krupa: Alternative mating tactics 1041
Table IV, Percentage of marked males that were observed as callers only, as satellites only, and exhibiting both tactics
Observation period
Males in Exclusively Displayed Exclusively chorus callers both tactics satellites
28-30May1984 Number of toads Mean snout-vent length (mm) (range) Mean number of observations per male (range)
14-18 May 1986 Number of toads Mean snout-vent length (mm) (range) Mean number of observations per male Ww)
34 9 (25%) 22 (66%) 3 (9%) 75.2 74.9 69.5
(64-8 1) (68-88) (66-72) 4.6 4.4 4.0
(3-6) (3-6) (3-5)
27 6 (22%) 20 (74%) 1 (4%) 81.4 83.8 69.0
(77-84) (78-98) 4.3 3.8 7
(3-7) C-8)
females were usually capable of moving through choruses without being detected by males. They typically approached (then remained motionless) wihin l-2 m of a calling male. On some occasions, they then moved on to other males. Otherwise, the female quickly rushed’ the male as he resumed calling and initiated amplexus by circling around or beside the caller, then bumping him (sometimes repeatedly) until he stopped calling and clasped her.
Satellites rarely succeeded at obtaining mates, because females easily avoided most non-callers. On 11 occasions, non-callers not within 1 m of a caller (e.g. satellites moving from one caller to the next) attempted to clasp females. The females easily escaped these males by darting away. During 13 female-release trials, females approached callers with one to three satellites. In each case the satellites attempted to clasp the females, but only three (23%) were successful. Two of these females inflated immediately when clasped by satellites located 20 and 40 cm from the caller. Both of these males later were found without the females. In the third case, the female approached a caller closely attended by a satellite (4 cm away). The satellite clasped the female as she made contact with the caller. She did not inflate and was found laying eggs the following morning while still amplexed by the satellite. The other 12 females laid eggs with mates that were calling when females initiated amplexus during the night. Thus callers acquired 92% of the matings versus 8% for satellites.
DISCUSSION
The results of this study indicate that variation in satellite frequency was best predicted by variation in male density, specifically the number of unmated males/m shoreline as found also by Sullivan (1982b), and more importantly, the number of callers per m shoreline. Thus, of the three predic- tions generated from the published hypotheses on the determinants of satellite frequency, only predic- tion 1 (satellite frequency increases as male density increases) was supported by this study. In contrast, predictions 2 and 3 were not supported by the results; satellite frequency was weakly influenced by male quality (prediction 2) as measured by variance in SVL, and satellite frequency was neither positively nor negatively influenced by operational sex ratio (prediction 3).
The observed lack of an effect of operational sex ratio and female density on satellite frequency is in some respects not surprising. The models predict- ing such influences tacitly assume some mecha- nisms whereby males assess these variables, but that assumption does not seem to apply to this species. Bufo cognatus is a nocturnally breeding, acoustically oriented anuran. Females are silent, and are usually able to move through choruses without being detected by males. If males cannot detect even the closest individual female, it is improbable that they could be aware of the total number of females available. An argument could be made that males might respond according to

1042 Animal Behaviour, 37, 6
predictable patterns of female abundance (e.g. behave as if anticipating that females are more abundant on the first night after a rainstorm). Yet no evidence of such anticipation was found in the present study: satellite frequency remained fairly constant over the duration of breeding bouts.
The results offer equivocal support for the hypothesis that a steep gradient in male quality can increase satellite frequency. Quality could refer to a male’s attractiveness to females or to his ability to compete with other males. In this species, long calls attract satellites and possibly females if Waltz’s (1982) prediction holds true (i.e. satellites associate with males most attractive to females; as demon- strated in bullfrogs; Howard 1978). Therefore, the variance in males’ ability for long calls or for time spent calling may influence satellite frequency more strongly than range in SVL, even though the results of this study indicate that males with longer calls tend to be slightly larger. Perhaps this is a side- effect of larger males being physically able to call longer. Call duration could also be influenced by how much energy has already been spent earlier in the evening or on previous nights. Unfortunately, the true range in calling ability was not obtained since males do not call while employing the satellite tactic. However, such data could be obtained by removing callers, then recording calls of the toads that switched to calling.
The male density models (Alexander 1975; Wells 1977; Arak 1983a) assume that male-male compe- tition for mates varies directly with male density. An alternative view is that competition for mates varies directly with the degree of male-bias in the operational sex ratio (Emlen & Oring 1977). Poss- ibly male density and operational sex ratio interact, such that as male density increases (due to more males entering the chorus), operational sex ratio becomes more male-biased increasing male-male competition. However, operational sex ratio was independent of male density in this study. This being the case, how could male density alone influence satellite frequency when call sites appear available? A plausible explanation is that high density may cause those males with less intense calls to lose relative vocal conspicuousness (Fores- ter & Czarnowsky 1985; Schwartz 1987) and thus switch to a tactic that does not require attracting females with advertisement calls. In such a situa- tion, males with relatively weak calls will tend to be unnoticed by females if they are near louder males. Such disadvantaged males may be more successful
trying (as satellites) to intercept females attracted to the louder male.
The large number of males that switch between tactics suggests that the satellite tactic may provide a temporary relief from energetic costs associated with calling. The resting male could improve his mating opportunities by associating with an attrac- tive caller, rather than associating with an un- attractive caller or sitting silently by itself. For B. cognatus, the best tactic for a male taking a break from calling may involve associating with males that have long calls that are more likely to attract females, and in so doing possibly detecting and intercepting an approaching female. If a female is not obtained during this time, then the satellite can move away and resume the calling tactic. Thus, the energetic constraints on call effort could influence satellite frequency independently of den- sity.
ACKNOWLEDGMENTS
Thanks are extended to C. Davis, L. MacIvor, M. Paulissen, B. Ploger and S. Secor for their field assistance; to C. Carpenter, T. Lamey, D. Mock, F. Sonleitner, P. Schwagmeyer, G. Shugart and B. Vestal for their helpful suggestions on various drafts of this manuscript; and to the Oklahoma Biological Survey for financial support during the study. Additional funding was provided by the University of Oklahoma Associates’ Fund, the University of Oklahoma Graduate Student Associ- ation Grant-in-Aid Program and the Society of Sigma Xi.
REFERENCES
Alexander, R. D. 1975. Natural selection and specialized chorusing behavior in acoustical insects. In: Insects, Science, and Society (Ed by D. Pimental), pp. 37-77. New York: Academic Press.
Arak, A. 1983a. Male-male competition and mate choice in anuran amphibians. In: -Mare Choice (Ed. by P. Bateson), pp. 181-210. Cambridge: Cambridee University Press.
Arak, A. 1983b. Sexual selection by male-male competi- tion in natterjack toad choruses. Nature, Land., 306, 261-262.
Arak, A. 1988. Callers and satellites in the nattejack toad: evolutionary stable decision rules. Anim. Behau.. 36,416-432. -
Bragg, A. N. 1936. Notes on the breeding habits, eggs, and embryos of Bufo cognates with a description of the tadpole. Copeia, 1936, 14-20.

Krupa: Alternatit ye mating tactics 1043
Bragg, A. N. 1937. Observations on Bufo cognatus with special reference to breeding habits and eggs. Am. Mdl. Nat., 18, 273-284.
Bragg, A. N. 1940. Observations on the ecology and natural history of anura. I. Habits, habitat and breed- ing of Bufo cognatus Say. Am. Nat., 14,322-349,424- 438.
Brown, L. E. &Pierce, J. R. 1967. Male-male interactions and chorusing intensities of the Great Plains toad, Bufo cognatus. Copeia, 1967, 149-154.
Davies. N. B. & Hallidav. T. R. 1979. Competitive mate searching in male common toads. Anim. Behav., 27, 1253-1267.
Dawkins, R. 1980. Good strategy or evolutionarily stable strategies? In: Sociobiology: Beyond Nature/Nurture? (Ed. by G.W. Barlow & J. Silverberg), pp. 331-367. Boulder, Colorado: Westview Press.
Emlen, S. T. & Oring, L. W. 1977. Ecology, sexual selection, and the evolution of mating systems. Science, N.Y., 1!97,215-223.
Engelman, L. 1979. PLR: stepwise logistic program. In: Biomedical Computer Programs (Ed. by W. J. Dixon & M. B. Brown), pp. 517.1-517.13. Berkeley: University of California Press.
Fairchild, L. 1984. Male reproductive tactics in an explosive breeding toad population. Am. Zoot., 24, 407418.
Fellers, G. M. 1979. Aggression, territoriality, and mat- ing behaviour in North American treefrogs. Anim. Behav., 27, 107-I 19.
Forester, D. C. & Czarnowsky, R. 1985. Sexual selection in the spring peeper, Hyla crucifer (Amphibia, Anura): role of the advertisement call. Behaviour, 92, 112-l 28.
Gatz, J., Jr. 1981. Non-random mating by size in American toads, Bufo americanus. Anim. Behav., 29, 1004-1012.
Howard, R. D. 1978. The evolution of mating strategies in bullfrogs, Rana catesbeiana. Evolution, 32,850-871.
Perrill, S. A. & Magier, M. 1988. Male mating behaviour in Acris crepitans. Copeia, 1988,245-248.
Robertson, J. G. M. 1986. Male territoriality, fighting and assessment of fighting ability in the Australian frog Uperoleia rugosa. Anim. Behav., 34, 763-772.
Schwartz, J. J. 1987. The function of call alternation in anuran amphibians: a test of three hypotheses. Evolu- tion, 41, 461-471,
Sokal, R. R. & Rohlf, 1981. Biomefry. 2nd Edn. San Francisco: W. H. Freeman.
Sullivan. B. K. 1982a. Sexual selection in Woodhouse’s toad (Bufo woodhouset) I. Chorus organization. Anim. Behav., %I, 680-686.
Sullivan, B. K.,l982b. Male mating behaviour in the Great Plains foad (Bufo cognatus). Anim. Behav., 30, 939-940.
Sullivan, B. K. 1983a. Sexual selection in the Great Plains toad (Bufo cognatus). Behaviour, 84,258-264.
Sullivan, B. K. 1983b. Sexual selection and mating system variation in the Great Plains toad (Bufo cognatus Say) and Woodhouse’s toad (Bufo woodhousei australis Shannon and Lowe). Ph.D. thesis, Arizona State University, Tempe Arizona.
Waltz, E. D. 1982. Alternative mating tactics and the law of diminishing returns: the satellite threshold model. Behav. Ecol. Sociobiol., 10, 75-83.
Waltz, E. D. & Wolf, L. L. 1984. By Jove!! Why do alternative mating tactics assume so many different forms? Am. Zool., 24, 3333343.
Wells, K. D. 1977. The social behaviour of anuran amphibians. Anim. Behav., 25,666-693.
(Received 22 June 1988; revised 9 September 1988; MS. number ~5236)