alternaria alternata causes petal blight and flower drop in harvested geraldton waxflower
TRANSCRIPT

Australasian Plant Pathology (1998) 27:
Alternaria alternata causes petal blight and flower drop in harvestedGeraldton waxflower
M.N.Taylor, A.H.Wearing,D.C.Joyceand D.H.Simons
School of Land and Food, The University of Queensland, Gatton College, Queensland 4345 Australia
Corresponding author: A.H. Wearing (Email [email protected])
Abstract
Six isolates ofAltemaria alternata were tested and shown to be pathogenic on detached flowers ofGeraldton waxflowercv. Alba. The fungus caused petal blight and abscission ofthe pedicel from inoculated flowers. This finding strengthensthe hypothesis that fungal infection of Geraldton waxflower flowers commonly leads to premature flower drop.Furthermore, it shows that, in addition to Botrytis cinerea, which has been shown previously to cause flower abscission,A. altemata can also cause flower abscission.
Introduction
Geraldton waxflower(Chameiaucium uncinatumSchauer, Myrtaceae) occurs naturally in WesternAustralia (Blomberry 1973), andiswidely grownforthe cut flower export market (Tomas et al. 1995).Postharvest flowerandbudabscission in Geraldtonwaxflower is a serious problem, whichisexacerbatedby handling and long distance transport (Joyce1988). Postharvest flower abscission has mainlybeen associatedwith infectionbyBotrytis cinereaPers. (Tomas et ai. 1995), and the resultantproductionof endogenousethylene; which. in turn, triggersflowerabscission(Tomas et ai. 1992; WearingandJoyce 1994). By comparison, physicalinjury, waterstress and exogenous ethylene are not seriouscausesof flower abscission(Joyce 1993).
Other fungi including Alternaria, Colie totrichum and Penicillium spp. have been isolatedfrom Geraldtonwaxflowerflowers(J. Alcornpersonal communication; Joyce 1993; Tomas et al.1992; 1995). Duringa surveyofGeraldtonwaxflowercropsin June/July 1994,anAitemaria-like funguswas recovered from samples of flowers showingdecay symptoms in Queensland, NewSouthWales,Western Australia and the Northern Territory (unpublished data). The fungus was subsequentlyidentified as Alternaria alternata (Fr.) Keissler
Australasian Plant Pathology Vol. 27 (3) 1998
(J. Alcorn, personal communication). Symptomsincluded brownnecroticlesionswhichappearedtoadvance fromthepetalmargin. Inextreme cases,theflowers had shrivelledand were coveredwith darkmycelium. Similar symptoms have been observedby the authors in cartons of cut, packagedGeraldton waxflower flowers. Theobjectives ofthisstudy were to establish the pathogenicity ofA. alternata on harvested Geraldton waxflowerflowers, and to compare symptom developmentwith that of grey mould causedby B. cinerea.
Methods
Isolates Four isolates of A. alternata obtainedfromflowers (cvv Albaand PurplePride)collectedin Queensland ('Ebonybrook', Gatton; 27°33' S,152°17'E)andtwo(cvvIntermediate PinkandDarkPink) fromNewSouthWales ('Tamlaght', Dareton;240 I' S, 142°I' E) usingtechniquessimilar to thoseofTomaset al. (1995)wereused in this study. Oneisolate of B. cinerea was obtained from cv. Albaflowers collected from a farm near Greenbank,Brisbane (27.28S, 153.02E). Briefly, sections ofdiseasedflower tissue were surfaced sterilised in0.1% sodium hypochlorite (NaOel) for60 sec, rinsedoncein steriledistilledwater, platedon onequarter
207

strength potato-dextrose agar (Oxoid) (1/4 PDA) andincubated in the dark at 25°C. Single-spore cultureswere prepared and maintained on 1/
4PDA.
Pathogenicity test In April 1995, fully openflowers with green hypanthiums and approximately10 mm long pedicels were obtained from 3- to 6month-old potted Geraldton waxflower (cv. Alba)plants produced in a growth cabinet under photoperiod conditions known to induce flowering(Dawson and King 1993). The photoperiod andtemperature regimes were 8 and 16 h day/night withlight intensity of 331 umol/sec/m' at canopy leveland 23 and 18± 2°C day/night, respectively. Detachedflowerswere surfacesterilisedin 0.1% NaOCLrinsedtwice in sterile distilled water, and allowed to air dry.Spore suspensions (l x 104 spores/mL) were prepared by flooding 10- to 14-day-old Petri dish cultures with sterile distilled water (plus 0.05% Tween80, MERCK Pty.Ltd). Flowers were immersed for5 sec in spore suspensions of either A. alternata orB. cinerea, transferred to sterile 200 mL glass vialscontaining moistened filter paper, capped and sealedwith low density polyethylene film (11 11m thick,Glad Wrap; GLAD Products of Australia). Each vialcontained two flowers (samples), and four vials(replicates) were prepared for each isolate (i.e. n == 8samples per isolate). Control flowers were preparedby immersing flowers in sterile distilled water and0.05% Tween 80. The vials were incubated in thedark at 20°C for 4 days. Flowers were inspected forsymptom development and abscission of the pedicel at days 2, 4 and 6. Flowers showing symptomswere photographed using a Canon DeSI digitalcamera. Re-isolations were made using techniquespreviously described. The test was repeated threetimes and results presented are the combined resultsof the three tests.
Results and Discussion
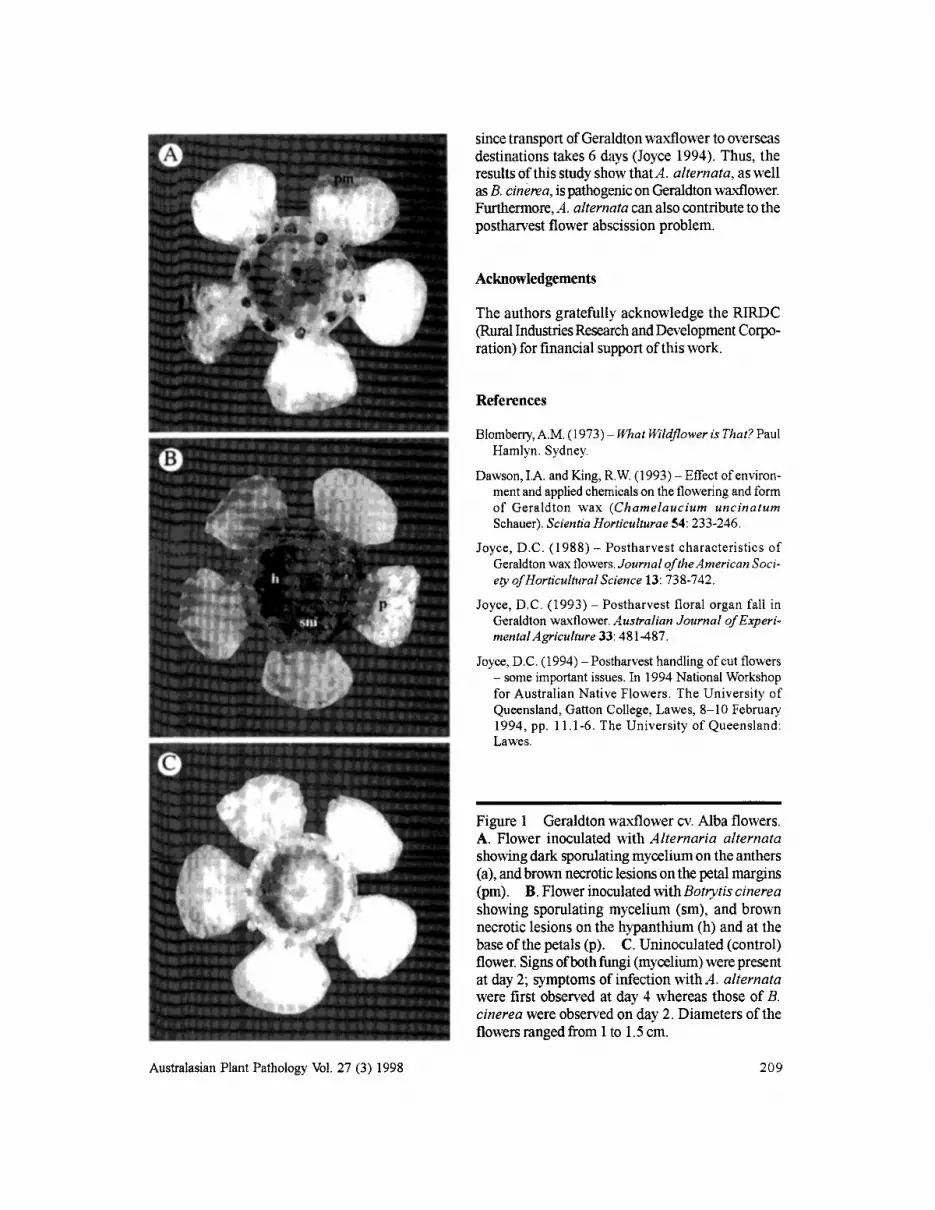
Within 48 h of inoculation with all isolates ofA. alternata, superficial mycelium was observedcolonising the stigma/style and anthers of mostflowers (n == 116). Four days after inoculation, allflowers (n == 144) showed signs of infection byA. alternata. Advancing necrotic brown lesions,initially starting at the petal margins, had also developed (n == 116). These signs and symptoms wereevident on all flowers (n == 144) inoculated with eachofthe six isolates ofA. alternata (n == 24 per isolate)
208
by day 6 (Figure lA). By day 4, abscission ofthepedicel had occurred in response to inoculationwith all isolates ofA. alternata (n == 76). No furtherabscission of pedicels was observed at day 6.
Within 48 h of inoculation with B. cinerea, allflowers (n == 24) showed signs of superficialmycelium and oflesion development. Water-soakedlesions started to develop at the base ofpetals andon the hypanthium, and advanced outwards.Abscission from the pedicel had occurred for mostflowers (n == 18) by day 2. By day 4, the lesions hadbecome necrotic-brown, and had worsened in thatmost lesions covered the entire hypanthium and allpetal surfaces (Figure lB). Mycelium had grown tocover most of the hypanthium by day 4, and wasfrequently sporulating. Abscission of the flowerfrom the pedicel had occurredfor all flowers (n == 24)by day 4.
Neither necrosis of the flower tissue nor abscission of the pedicel occurred in the uninoculated(control) flowers (n == 24; Figure 1C).
A. alternata and B. cinerea were re-isolatedfrom all flowers onto which they had each beeninoculated (n == 24 per isolate). However, neitherA. alternata nor B. cinerea was isolated from theuninoculated (control) flowers.
Cultures were lodged at the Department ofPrimary Industries, Queensland, Plant PathologyHerbarium, with the following accession numbers:BRIP 22823 to BRIP 22828 for the six isolates ofA. alternata, and BRIP 22936 for the isolate ofB. cinerea.
In summary, all six isolates ofA. alternata testedwere shown to be pathogenic on Geraldtonwaxflower cv. Alba flowers. Disease developmentand symptoms caused byA. altemata differed fromthose of B. cinerea in three ways. Firstly, necroticpetal lesions caused by A. alternata were initiatedfrom the petal margins and progressed inwards(Figure lA). In contrast, lesions caused by B. cinerea started from the base of the petal and grew outwards (Figure IB). Secondly, the superficialmycelium ofA. alternata was dark, greyish-brown,whereas the mycelium of B. cinerea was light grey.Mycelium ofboth fungi had appeared on flowersby day 2. Thirdly, the symptoms caused byA. alternata took longer to develop than thosecaused by B. cinerea (i.e. 4 compared to 2 days at20°C). Nevertheless, this relative delay in diseasedevelopment caused by A. alternata does notdecrease the probability that this pathogen contributes to the postharvest flower abscission problem,
Australasian Plant Pathology Vol. 27 (3) 1998
Australasian Plant Pathology Vol. 27 (3) 1998
since transportofGeraldton waxflower tooverseasdestinations takes 6 days (Joyce 1994). Thus, theresultsof this studyshowthatA. alternata, as wellasB. cinerea, ispathogenic onGeraldton waxflower.Furthermore, A. alternata canalsocontribute to thepostharvest flower abscission problem.
Acknowledgements
The authors gratefully acknowledge the RIRDC(Rural Industries Research andDevelopment Corporation) forfinancialsupport of this work.
References
B1omberry, A.M. (1973) - What Wildflower is That? PaulHamlyn. Sydney.
Dawson, LA. and King, R.w. (1993) - Effect ofenvironment and applied chemicals on the flowering and formof Geraldton wax (Chamelaucium uncinatumSchauer). Scientia Horticulturae 54: 233-246.
Joyce, D.C. (1988) - Postharvest characteristics ofGeraldton wax flowers. Journal ofthe American Society ofHorticultural Science 13: 738-742.
Joyce, D.C. (1993) - Postharvest floral organ fall inGeraldton waxflower. Australian Journal ofExperimental Agriculture 33: 481-487.
Joyce, D.C. (1994) - Postharvest handling ofcut flowers- some important issues. In 1994 National Workshopfor Australian Native Flowers. The University ofQueensland, Gatton College, Lawes, 8-10 February1994, pp. 11.1-6. The University of Queensland:Lawes.
Figure 1 Geraldton waxflower cv. Albaflowers.A. Flower inoculated with Alternaria alternatashowing dark sporulating mycelium on the anthers(a),andbrown necrotic lesions onthe petalmargins(pm). B. Flower inoculated withBotrytis cinereashowing sporulating mycelium (sm), and brownnecrotic lesionson the hypanthium(h) and at thebaseofthe petals (p). C. Uninoculated (control)flower. Signs ofbothfungi (mycelium) werepresentat day2; symptoms of infectionwithA. alternatawere first observed at day 4 whereas those of B.cinerea wereobserved on day 2. Diametersof theflowers rangedfrom1to 1.5em.
209

Tomas, A, Wearing, AH. and Joyce, D.C. (1992)Geraldton waxflower, flower abscission and Botrytis:a hypothesis. In High Quality Horticulture, Practicesand Products, pp. 17-19. Australian Society for Horticultural Science Inc. Regional Technical Meeting, 29August 1992, Brisbane.
Tomas, A, Wearing, AH. and Joyce, D.C. (1995)Botrytis cinerea: a causal agent of premature flowerdrop in packaged Geraldton waxflower. AustralasianPlant Pathology 24: 26-28.
210
Wearing, A.H. and Joyce, D.C. (1994) - Botrytis cinereaon Geraldton waxflower. In 1994 National Workshopfor Australian Native Flowers, pp. 11.22-11.24. TheUniversity ofQueensland, Gatton College, 8-10 February 1994, Lawes.
Manuscript received 22 December 1997, accepted 30Apri11998.
Australasian Plant Pathology Vol. 27 (3) 1998