aging and the myelinated fibers in prefrontal cortex and corpus callosum of the monkey
TRANSCRIPT

Aging and the Myelinated Fibers inPrefrontal Cortex and Corpus Callosum
of the Monkey
ALAN PETERS* AND CLAIRE SETHARES
Department of Anatomy and Neurobiology, Boston University School of Medicine,Boston, Massachusetts 02118
ABSTRACTIn the rhesus monkey, the myelin sheaths of nerve fibers in area 46 of prefrontal cortex
and in splenium of the corpus callosum show age-related alterations in their structure. Thealterations are of four basic types. Most common is splitting of the dense line of myelinsheaths to accommodate electron dense cytoplasm derived from the oligodendroglia. Lesscommon are splits of the intraperiod line to form balloons or blisters that appear to containfluid, the occurrence of sheaths with redundant myelin, and thick sheaths that are almostcompletely split so that one set of compact lamellae is surrounded by another set. But despitethese alterations in the sheaths, few nerve fibers show axonal degeneration. To quantify thefrequency of the age-related alterations in myelin, transversely sectioned nerve fibers fromthe splenium of the corpus callosum and from the vertical bundles of nerve fibers within area46 were examined in electron photomicrographs. The material was taken from 19 monkeys,ranging between 5 and 35 years of age. It was found that the frequency of alterations inmyelin sheaths from both locations correlates significantly with age. In area 46, the age-related alterations also significantly correlate (P � 0.001) with an overall assessment ofimpairment in cognition, i.e., the cognitive impairment index, displayed by individual mon-keys. The correlation is also significant when only the old monkeys are considered as a group.A similar result was obtained previously in our examination of the effects of age on the myelinsheaths of nerve fibers in primary visual cortex (Peters et al. [2000] J Comp Neurol.419:364–376). However, in the corpus callosum the myelin alterations correlate significantlywith only one component of the cognitive impairment index, namely the delayed nonmatchingto sample task with a 2-minute delay. It is proposed that age-related myelin alterations areubiquitous and that the correlations between their frequency and impairments in cognitionoccur because the conduction velocity along the affected nerve fibers is reduced, so that thenormal timing sequences within neuronal circuits break down. J. Comp. Neurol. 442:277–291, 2002. © 2001 Wiley-Liss, Inc.
Indexing terms: normal aging; myelin; rhesus monkeys; area 46; cognition; senescence
The rhesus monkey provides a very useful model forstudying the effects of age on the primate cerebral cortex,because as with normally aging humans, rhesus monkeysshow a cognitive decline as they age (e.g., Presty et al.,1987; Rapp and Amaral, 1989; Arnsten and Goldman-Rakic, 1990; Bachevalier et al., 1991; Albert and Moss,1996; Moss et al., 1997, 1999). However, rhesus monkeysdo not develop Alzheimer’s disease, and their cortical neu-rons do not form neurofibrillary tangles. Consequently, inthe rhesus monkey, the effects of normal aging can bestudied without the possibility of early stages of Alzhei-mer’s disease intruding into the population being exam-ined. Senile plaques do occur in some of the older mon-
keys, but there is no correlation between their frequencyand the extent of the cognitive decline exhibited by theolder monkeys (Sloane et al., 1997). Another advantage ofusing the rhesus monkey is that its life span is less than
Grant Sponsor: NIH National Institute on Aging; Grant number: 2POAG 00001.
*Correspondence to: Alan Peters, Department of Anatomy and Neurobi-ology, Boston University School of Medicine, 715 Albany Street, Boston MA02118. E-mail: [email protected]
Received 6 August 2001; Revised 11 October 2001; Accepted 23 October2001
THE JOURNAL OF COMPARATIVE NEUROLOGY 442:277–291 (2002)
© 2001 WILEY-LISS, INC.DOI 10.1002/cne.10099
that of the human. Rhesus monkeys reach sexual matu-rity at 5 years of age, and the maximum age they attain is35 years (Tigges et al., 1988), with only approximately25% of the monkeys surviving for 25 years and only some6% reaching 30 years of age. Furthermore, after monkeyshave been behaviorally tested their brains can be fixed toachieve the quality of preservation necessary for detailedmorphologic studies.
Examination of the cerebral cortex of monkeys hasshown that there is no significant loss of neurons with agefrom visual cortex (Vincent et al., 1989; Peters andSethares, 1993; Kim et al., 1997; Peters et al., 1997; Hof etal., 2000), motor cortex (Tigges et al., 1990), or area 46 ofprefrontal cortex (Peters et al., 1994). These studies, andothers, have led to the conclusion that loss of neurons fromthe cerebral cortex is not the basis of the age-relatedcognitive decline displayed by primates (Morrison andHof, 1997; Peters et al., 1998a). However, cerebral cortexis not unaffected by normal aging, because significantalterations occur in layer 1 in some parts of the cortex,such as area 46 and area 17 (Peters et al., 1998b, 2001a).
Age-related morphologic changes that appear to bemore ubiquitous are ones affecting myelin. Alterations inmyelin were first noticed in light microscopic studies. Forexample, Lintl and Braak (1983) carried out densitometricstudies on the intensity with which hematoxylin stainsmyelin in the line of Gennari in the striate cortex ofnormal humans between 18 and 96 years of age. Theyfound that, from the third decade onward, the intensity ofstaining is gradually reduced. Kemper (1994) also drewattention to the increased pallor of staining of intracorti-cal fibers in the human brain with age, especially in asso-ciation cortical areas, and in fiber tracts that myelinatelate in life. More recently, studies of white matter by usingaxial MRI scans on both human (Albert, 1993; Guttmannet al., 1998) and rhesus monkey brains (Lai et al., 1995)have shown that white matter is lost from the cerebralhemispheres during aging. By using stereology, Pakken-berg and Gundersen (1997) have also reported a loss ofwhite matter from human cerebral hemispheres with ageand Tang et al. (1997) have found that, compared withthose of young humans, the cerebral hemispheres of old
humans have 15% less white matter, with a 27% less totallength of myelinated fibers.
Electron photomicroscopy shows age-related alterationsin the structure of myelin sheaths in both the white andgrey matter of the cerebral cortex (Peters, 1991; Feldmanand Peters, 1998). With age, some myelin sheaths developblebs that balloon out from myelin internodes (Feldmanand Peters, 1998), whereas other sheaths in old monkeyshave splits containing dark cytoplasm. In addition, someof the thicker sheaths have extensive circumferentialsplits so that they appear to be composed of two sets oflamellae, although yet other sheaths have redundant my-elin, such that the sheath is much too large for the en-closed axon (Peters et al., 2001b). Concomitant with thesestructural alterations, there is an increase in the thick-ness of myelin sheaths in primary visual cortex with age,such that the mean number of lamellae increases from 5.6in young monkeys to 7 in old monkeys (Peters et al.,2001b). Much of this increase in the mean thickness isbecause thick myelin sheaths with more than 10 lamellaeare more common in old monkeys, indicating that eventhough there is some degeneration of myelin, with agethere is a continued production of myelin lamellae.
Other studies of monkey primary visual cortex haveshown that, although there is no loss of nerve fibers from
TABLE 1. Data on Age, Sex, Altered Myelin Sheaths, and Behavior1
AnimalAge(yr) Sex
Nerve fibers in area 46 Nerve fibers in corpus callosum
CIINumber
examinedNormal myelin
(%)Number
examinedNormal myelin
(%)
AM 58 4 M Not available 1543 99.8 No dataAM 16 5 M 981 98.7 Poor fixation 0AM 76 6 F 1,113 99.7 1413 99.9 0.09AM 77 6 F 759 99.1 1394 99.8 1.14AM 47 9 M 980 99.0 1369 99.6 �0.54AM 96 9 F 506 98.7 1815 99.7 2.46AM 53 10 M 944 98.2 1080 98.7 0.08AM 19 25 F 726 96.0 1346 92.6 1.68AM 100 25 F 677 94.8 1618 92.1 4.12AM 12 27 F 970 95.2 Poor fixation 4.05AM 15 27 F 873 94.3 1853 90.7 1.56AM 62 27 M 973 94.8 1521 96.1 4.26AM 27 28 M 999 96.0 Poor fixation 1.34AM 26 29 (est) F 970 96.0 1659 96.3 0.94AM 17 29 (est) F 977 91.6 Poor fixation 3.24AM 91 32 M 965 97.2 1389 95.5 0.17AM 23 32 F 984 94.0 Poor fixation 5.31AM 41 32 F 755 93.8 Poor fixation 4.81AM 13 35 M 1,028 93.7 1428 92.3 No data
1CII, Cognitive Impairment Index; est, estimated.
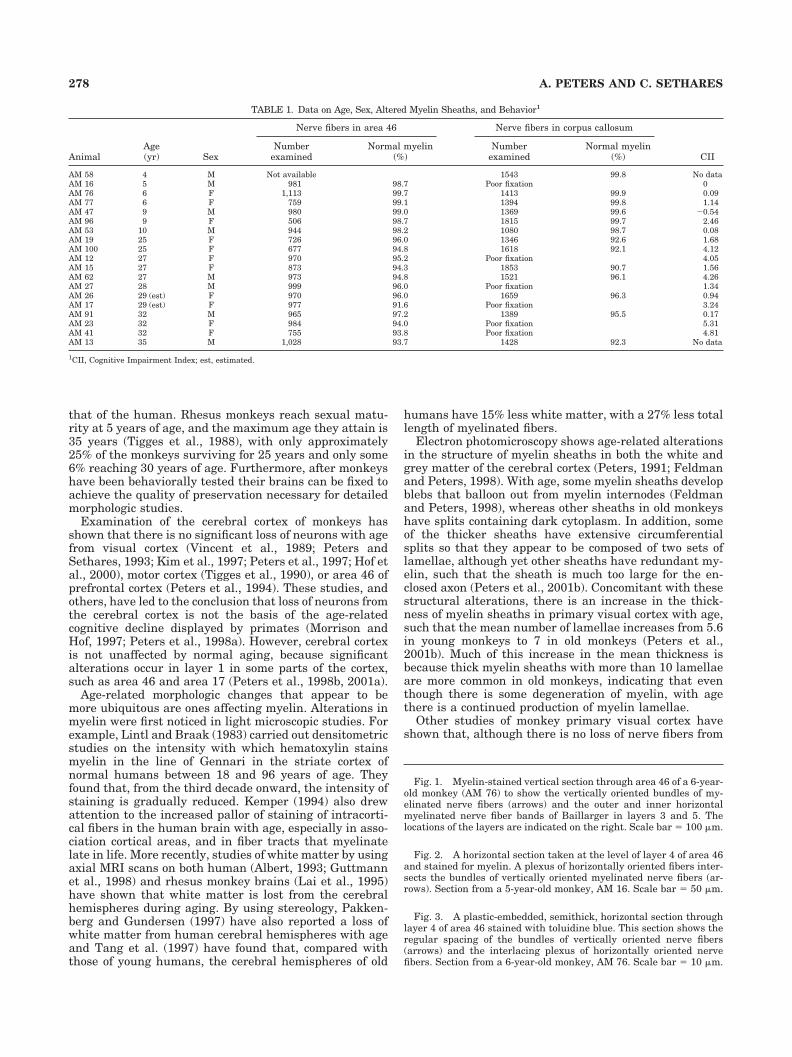
Fig. 1. Myelin-stained vertical section through area 46 of a 6-year-old monkey (AM 76) to show the vertically oriented bundles of my-elinated nerve fibers (arrows) and the outer and inner horizontalmyelinated nerve fiber bands of Baillarger in layers 3 and 5. Thelocations of the layers are indicated on the right. Scale bar � 100 �m.
Fig. 2. A horizontal section taken at the level of layer 4 of area 46and stained for myelin. A plexus of horizontally oriented fibers inter-sects the bundles of vertically oriented myelinated nerve fibers (ar-rows). Section from a 5-year-old monkey, AM 16. Scale bar � 50 �m.
Fig. 3. A plastic-embedded, semithick, horizontal section throughlayer 4 of area 46 stained with toluidine blue. This section shows theregular spacing of the bundles of vertically oriented nerve fibers(arrows) and the interlacing plexus of horizontally oriented nervefibers. Section from a 6-year-old monkey, AM 76. Scale bar � 10 �m.
278 A. PETERS AND C. SETHARES
Figures 1–3
279MYELIN AND AGING

the vertically oriented, intracortical myelinated fiber bun-dles with age (Nielsen and Peters, 2000), the frequency ofthe age-related alterations in the normal structure of themyelin sheaths of nerve fibers in these bundles correlatessignificantly with both increased age and increased cogni-tive impairment (Peters et al., 2000). However, it is notknown whether these correlations with myelin sheath al-terations are unique for striate cortex. The present studyis designed to partially answer that question by examin-ing the effects of increased age on the integrity of themyelin sheaths in area 46 of prefrontal cortex and in thesplenium of the corpus callosum. Area 46 is considered tobe pivotal in a network of functions that subserve workingmemory (e.g., Goldman-Rakic, 1996; Petrides, 1996), aswell as higher executive functions (Goldman, et al., 1971),whereas the corpus callosum is the principal commissureof the neocortex.
MATERIALS AND METHODS
Animals
Pieces of area 46 in prefrontal cortex and pieces of thesplenium of the corpus callosum were taken from 18 rhe-sus monkeys (Macaca mulatta) ranging from 5 to 35 yearsof age. Four of the monkeys were young (5–6 years), 3were middle aged (9–10 years), and 12 were old (over 25years). The sexes of the monkeys and their ages roundedto the nearest year are given in Table 1. Details of theprotocol for fixing the brains are given in an earlier pub-lication (Peters et al., 1994). The perfusions were carriedout in full accordance with the approved InstitutionalAnimal Care and Use Committee regulations. In sum-mary, the monkeys were preanesthetized with ketamine(6.5 mg/kg). Sodium pentobarbital was then administeredintravenously (approximately 35–45 mg/kg) until themonkey was deeply anesthetized and a state of areflexiawas attained. The monkeys were intubated into the tra-chea and artificially respired by using a mixture of CO2and O2. The chest was opened, and the monkeys wereperfused intra-aortically with a warm solution of 1% para-formaldehyde and 1.25% glutaraldehyde in 0.1 M cacody-late or phosphate buffer at pH 7.4. After this initial fixa-tion, the brain was removed and one hemisphere fixedfurther by immersion in a cold solution of 2% paraformal-dehyde and 2.5% glutaraldehyde in 0.1 M cacodylate orphosphate buffer at pH 7.4 for several days.
Tissue preparation
Tissue blocks, approximately 2 mm thick, were taken.The blocks from area 46 of prefrontal cortex were takenfrom the floor of the principal sulcus at the level of therostral end of the corpus callosum (see Peters et al., 1994).This portion of prefrontal cortex is part of area 8 of Brod-mann (1905) and is designated by Walker (1940) as be-longing to area 46. It was chosen because it has a rela-tively even thickness and a similar piece of tissue can betaken from each monkey. To ensure consistency, theblocks of corpus callosum were taken from the upper ordorsal half of the splenium. All blocks of cortex wereosmicated, dehydrated in an ascending series of alcohols,and then embedded in Araldite.
For the examination of the effects of age on myelinatednerve fibers in area 46, two blocks of cortex were randomlyselected from each monkey. Semithick (1 �m) sections
were taken in the vertical plane, stained with toluidineblue, and used to determine the depth of layer 4. Theblocks of cortex were then turned, sectioned in the tan-gential plane until layer 4 was reached, and a set ofsemithick sections stained for light microscopic evalua-tion. Thin sections were then taken to examine the cross-sectioned nerve fibers contained in the vertical fiber bun-dles. For examination of the nerve fibers in corpuscallosum the tissue blocks from the dorsal portion of thesplenium were oriented to obtain cross sections of thenerve fibers. All thin sections were stained with uranylacetate and lead citrate for examination in the electronmicroscope.
Quantitative analysis
After the thin sections of area 46 and of the corpuscallosum had been examined to determine the quality ofthe fixation and nature of the effects of age on the mor-phology of the nerve fibers, electron photomicrographswere taken at an initial magnification of 4,000� or 5,000�and printed to a final magnification of 10,000� or12,500�. For each monkey, at least 500 profiles of cross-sectioned nerve fibers in the vertical bundles of area 46and in the corpus callosum were examined to ascertain thepercentage of these nerve fiber profiles that displayedage-related changes in either their myelin sheaths or ax-ons. The results of these analyses were then correlatedwith age and with the behavioral data available for sev-enteen of the monkeys (see Table 1).
Behavioral testing
The behavioral tests used to determine the cognitivestatus of individual monkeys are described in detail inearlier publications (e.g., Herndon et al., 1997; Moss et al.,1999; Peters et al., 1996, 2000; Killiany et al., 2000). Thebattery of tests included three visual recognition tasks.They are (1) the delayed nonmatching to sample (DNMS)task, (2) a DNMS task with a 2-minute delay, and (3) thedelayed recognition memory span (DRS) task. From thescores achieved on these tasks by each monkey, an overallmeasure of cognitive impairment (Peters et al., 1998b),the Cognitive Impairment Index (CII), was derived. Spe-cifically, the individual scores on the three behavioralmeasures were transformed to scores normalized to a pop-ulation of 53 adult rhesus monkeys, as described by Hern-don et al. (1997), and a composite score, the CognitivePerformance Index, was derived. The Cognitive Perfor-mance Index has been shown to be a practical index ofglobal ability, and its inverse, the CII, is a measure ofcognitive impairment. We used the CII as a behavioralmeasure against which to relate the frequency of myelinsheaths that show age-related alterations.
Photomicrograph production
The photomicrographs used to illustrate this articlewere produced by using standard photographic and dark-room methods.
RESULTS
Prefrontal area 46
Light microscopy. In myelin stained preparations ofsections taken normal to the pial surface vertically ori-ented bundles of myelinated fibers are apparent (Fig. 1).
280 A. PETERS AND C. SETHARES
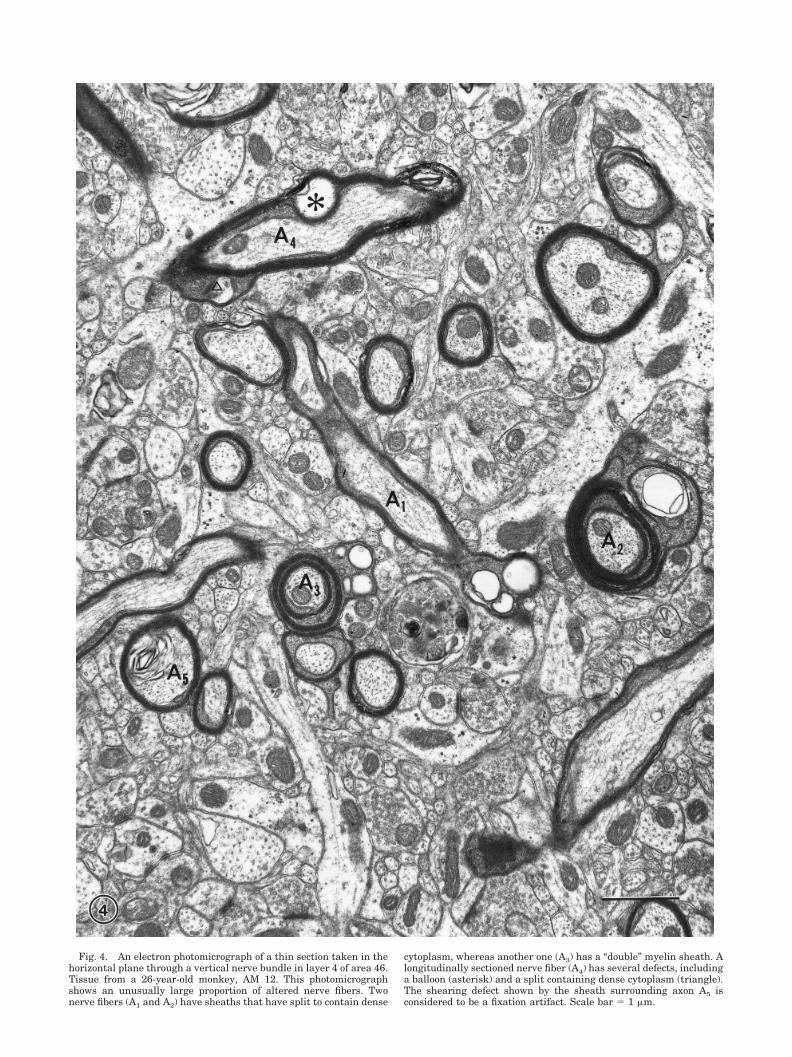
Fig. 4. An electron photomicrograph of a thin section taken in thehorizontal plane through a vertical nerve bundle in layer 4 of area 46.Tissue from a 26-year-old monkey, AM 12. This photomicrographshows an unusually large proportion of altered nerve fibers. Twonerve fibers (A1 and A2) have sheaths that have split to contain dense
cytoplasm, whereas another one (A3) has a “double” myelin sheath. Alongitudinally sectioned nerve fiber (A4) has several defects, includinga balloon (asterisk) and a split containing dense cytoplasm (triangle).The shearing defect shown by the sheath surrounding axon A5 isconsidered to be a fixation artifact. Scale bar � 1 �m.

These nerve fiber bundles make their appearance at thelevel of layer 3 and become gradually thicker as they passthrough the horizontal fibers in the outer and inner bandsof Baillarger to reach the white matter. However, horizon-tally oriented fibers are not confined to the bands of Bail-larger. Even at the level of layer 4, numerous horizontallyoriented myelinated fibers are visible in sections taken inthe horizontal plane and they criss-cross through the cor-tex, largely intersecting with the bundles of verticallyoriented nerve fibers (Fig. 2). This arrangement of thevertically and horizontally oriented myelinated fibers isseen to better advantage in semithick plastic sectionstaken in the horizontal plane at the level of layer 4 (Fig. 3).It is then evident that the vertically oriented bundles havea center-to-center spacing of approximately 40�m, andthat they are composed of rather loosely packed fibers,with horizontally oriented fibers passing through them.Between the bundles of nerve fibers the cell bodies ofneurons, astrocytes, and oligodendrocytes are visible.
Electron microscopy. It is rare to obtain perfect pres-ervation of the myelin sheaths of nerve fibers in cerebralcortex. Some profiles of myelin sheaths invariably showfixation defects and commonly these defects are manifestas a localized shearing of the lamellae in a segment of thesheath (see Fig. 4; A5). Such shearing causes the lamellaeto split apart so that they become detached from eachother, causing the widened sheath to bulge into the axonor into the surrounding extracellular space.
In the cortices of older monkeys, there are additionalalterations in the morphology of myelin sheaths, andthese are associated with aging. In monkeys over 25 yearsof age, usually between 3 and 6% of the sheaths of nervefibers profiles in layer 4 of area 46 show these age-relatedalterations. Examples are shown in Figure 4. This imageis from a 26-year-old monkey and is a field in which thereis an unusually high proportion of nerve fiber profiles withage-related alterations.
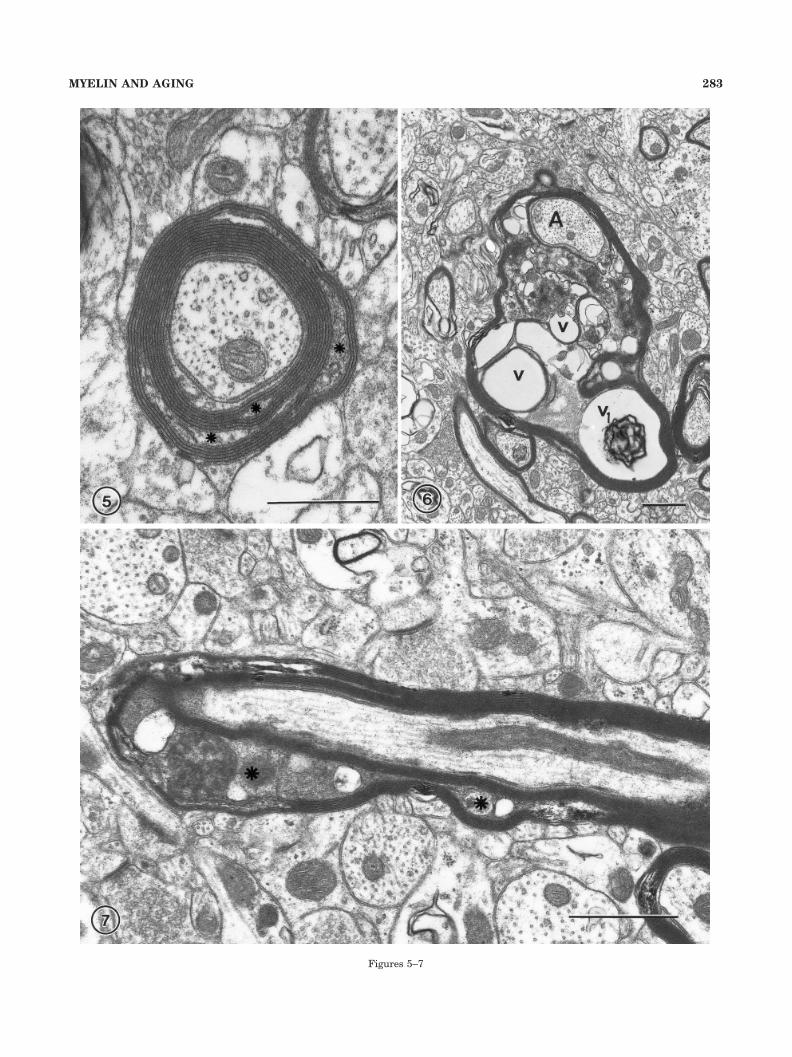
The age-related alterations are of various types. Themost common alteration involves a splitting of the majordense line on one side of the sheath to enclose electrondense cytoplasm (Figs. 4A2, 5–7). Approximately 80% ofaltered sheaths display this type of defect. Sometimes theamount of enclosed cytoplasm is small, as in the exampleshown in Figure 5, but in other examples the separation ofthe lamellae is more extensive and a voluminous densecytoplasm is accommodated within several splits (e.g.,Figs. 4, 6). In such cases, the dense cytoplasm usuallycontains vacuoles and dense bodies, some of which appearto be amorphous whereas others are bound by mem-branes. As shown in Figure 7, which is a longitudinalsection through a nerve fiber, several dense cytoplasmicinclusions can occur in the same internode and each onecan extend for several microns.
Another age-related alteration shown by approximately10% of altered sheaths is one in which the myelin balloonsout from the side of a sheath (Feldman and Peters, 1998).Such balloons are generated by splits in the intraperiodline of a sheath to accommodate large vesicles, whichappear to be fluid filled. Sometimes, as shown in Figure 8,the vesicle contains debris, but in examples in which theballoons are larger, the contents of the vesicle appear to beentirely fluid (Fig. 9). Essentially, the profiles of theseballoons are more or less circular in all planes of section,indicating that they are basically spherical; and, as shownin Figure 10, the ballooned segments of internodes can
occur adjacent to locations where there is dense cyto-plasm.
With age, there is also an increased frequency of nervefibers in which the myelin sheaths are too large for theenclosed axon. In these few cases the axon is located at oneend of the profile of a sheath that loops off to the side. Andfinally there is an increase in the frequency of occurrenceof so-called “double” myelin sheaths in which one set ofcompact lamellae is surrounded by a second, outer set.Sometimes cytoplasm occurs between the two sets of la-mellae (see Fig. 4; A3); but in thick sheaths, the spaceseparating the lamellae into inner and outer sets appearsto contain fluid.
Despite the changes in sheaths, the majority of axonsappear not to be affected by age. In the few altered axonsthat have been encountered, some have a homogeneouslydense axoplasm, but in others, the dense cytoplasm con-tains numerous vacuoles or it may be filled with lysosomes(Fig. 11). Such alterations are taken to indicate that theaxons are degenerating, and this process is consistentwith the occasional finding of myelin figures that appearto surround empty spaces, although some profiles of thistype can be sections through small myelin balloons.Lastly, a few axons have their cytoplasm filled with mito-chondria, and such profiles can be very large, suggestingthat the axon has become constricted in another location,causing the mitochondria to dam up.
To complete the picture of the effects of age on layer 4 ofarea 46, it should be mentioned that the only effect ageappears to have on the cell bodies of neurons is that theyhave an increased number of lipofuscin granules in theircytoplasm. Many of the neuroglia also have inclusions intheir cytoplasm and these are quite distinctive for theneuroglial cell type involved. Many of the oligodendrocytesin older monkeys have dense inclusions both in theirperikaryal cytoplasm and in swellings that occur alongtheir processes (Peters, 1996). Most of the perikarya ofmicroglia and astrocytes in old monkeys have phagocy-tosed debris within them and often this debris can be quitevoluminous (see Peters et al., 1991; Peters, 1999). Fur-thermore, myelin figures have been encountered in theperikarya and processes of several astrocytes (Fig. 12),suggesting that they have phagocytosed myelin from de-generating sheaths.
Corpus callosum
Being a tract of white matter, the corpus callosum con-sists of myelinated nerve fibers with the cell bodies of
Fig. 5. A nerve fiber with a sheath in which the major dense linehas split in several places to accommodate dense cytoplasm (aster-isks). Layer 4 in area 46 of a 35-year-old monkey, AM 13. Scale bar �0.5 �m.
Fig. 6. In this nerve fiber, the axon (A) is at one side of a verydistorted sheath that has split to contain dense cytoplasm in whichvacuoles (v) are embedded. One of the vacuoles (v1) contains membra-nous debris. Layer 4 in area 46 of a 35-year-old monkey, AM 13. Scalebar � 1 �m.
Fig. 7. A longitudinally sectioned nerve fiber to show that severalsplits of the myelin sheath that accommodate dense cytoplasm (as-terisks) can occur along the same fiber and extend for some distancealong the sheath. Layer 4 in area 46 of a 27-year-old monkey, AM 15.Scale bar � 1 �m.
282 A. PETERS AND C. SETHARES
Figures 5–7
283MYELIN AND AGING

astrocytes, oligodendrocytes, and microglial cells scat-tered between them. In the splenium of the corpus callo-sum of young monkeys, the nerve fibers range in diameterbetween approximately 0.5 and 5 microns (Fig. 13). Theyare closely packed together, with a few sheets of astrocyticprocesses partially separating the nerve fibers into fasci-cles. Unfortunately, the preservation of the corpus callo-sum is rarely good, and consequently, many of the myelinsheaths exhibit shearing defects of the type describedabove for nerve fibers in the area 46 of cerebral cortex (Fig.13, arrows). Also, it is not uncommon to encounter nervefibers with swollen axons and astrocytes with swollenperikarya and processes.
In the corpus callosum of older monkeys the nerve fibersare generally more loosely packed than in young monkeys(Fig. 14), suggesting that some nerve fibers might havebeen lost with age. This is consistent with the fact that, ina few nerve fiber profiles, the axons show signs of degen-eration like those described for area 46. Although some ofthe myelin sheaths exhibit shearing defects due to poorpreservation (Fig. 14; N1,N2), others display age-relatedalterations similar to those shown by myelin sheaths inarea 46. The most common age-related change shown byapproximately 75% of altered sheath profiles is the one inwhich myelin lamellae have split to accommodate densecytoplasm (Fig. 14; N3–N7). The next most common defectshown by approximately 15% of sheaths, and especiallythe thick sheaths of large diameter axons, is the splittingof the sheath so that it appears to be double. Less com-monly encountered are myelin balloons and sheaths toolarge for the enclosed axons. As in area 46, the neurogliain the corpus callosum of old monkeys also exhibit age-related alterations. The astrocytes are more voluminousthan in young monkeys, and their processes are moreobvious, because they occupy the spaces between the morewidely separated nerve fibers. Also, as in the cerebralcortex the oligodendrocytes often have dense inclusions intheir cell bodies and most of the microglia contain phago-cytosed material.
Unfortunately when the nerve fibers are poorly pre-served, as they are in the corpora callosa of several the oldmonkeys, and even in young animals (see Table 1), it canbe difficult to assess whether alterations in myelinsheaths are to be attributed to poor preservation or toage-related changes. Consequently, the corpora callosa ofsuch monkeys cannot be used for quantitative analyses.
Quantification of age-related alterationin nerve fibers
To generate a measure of the effects of age on thestructure of myelin sheaths, electron photomicrographswere taken of cross-sectioned nerve fibers both in thevertical bundles that traverse layer 4 of area 46, and inthe corpus callosum. Nerve fibers were considered to becross-sectioned if the lengths of their profiles were notmore than twice their widths and if the microtubules andneurofilaments were not obviously sectioned obliquely.The cross-sectioned nerve fibers were then examined in-dividually to determine whether their sheaths appeared tobe normal, other than for the presence of the shearingdefects which are regarded as artifacts of fixation, or if thesheaths showed any of the age-related defects describedabove. As pointed out, in several old monkeys, the preser-vation of the nerve fibers in the corpus callosum was sopoor that it was not possible to be certain that defects in
myelin sheaths were due to inferior fixation or to agechanges. These corpora callosa of such monkeys, listed inTable 1, had to be omitted from the study.
It is realized that the quantification of these age-relateddefects, by using only profiles of transversely sectionednerve fibers, does not reveal the true number or proportionof nerve fibers with defective sheaths, because the defectscan extend from several microns along the length of aninternode (see Figs. 7, 10). However, such quantificationdoes provide a comparative measure by which it can bedetermined whether the myelin sheaths in one monkeydisplay more or fewer age-related defects than those inothers. The results of this analysis are given in Table 1,which lists the number of nerve fiber profiles examined inboth area 46 and in the corpus callosum of each monkeyand the percentage of those profiles with myelin sheathsthat are normal.
Correlation of myelin sheath changes withage and behavior
By using the data given in Table 1, the plot in Figure 15shows the percentage of nerve fibers that display normalsheaths both in area 46 and in the corpus callosum versusthe ages of the monkeys. For both structures, there is asignificant correlation between age and the increase in theproportion of nerve fibers that possess abnormal sheaths(area 46: r � -0.86, p � 0.001; corpus callosum: r � -0.84,p � 0.001).
In Figure 16, the percentage of nerve fibers with normalsheaths is plotted versus the cognitive impairment index(CII), a measure of the overall learning deficiencies dis-played by the monkeys. For area 46, there is a significantcorrelation between the frequency of age-related myelindefects and the CII (r � -0.74; p � 0.001). This findingsuggests that cognition in monkeys is dependent on theintegrity of the myelin sheaths in area 46. Furthermore,when only the 12 old monkeys over 25 years of age areconsidered as a group, there is also a significant correla-tion between the proportion of abnormal sheaths in area46 and their cognitive impairment indices (r � 0.62; p �0.05), as well as with the three individual components thatmake up the CII.
Figure 16 also shows the plot of the percentage of nor-mal sheaths in the corpus callosum against the CII, and inthis case, there is not a significant correlation between thetwo measures (r � -0.45, p NS), perhaps partly due to thesmall number of monkeys (n � 11) for which data on boththe CII and the percentage of normal sheaths are avail-able. However, when the frequency of normal sheaths inthe corpus callosum is plotted against the three separate
Fig. 8. A nerve fiber in which the sheath has formed a smallballoon (b) that contains debris. Layer 4 in area 46 of a 35-year-oldmonkey, AM 35. Scale bar � 1 �m.
Fig. 9. An axon (A) with a sheath that has a large balloon (b), 4 �min diameter, protruding from one side. Layer 4 from area 46 of a29-year-old monkey, AM 26. Scale bar � 1 �m.
Fig. 10. The sheath of this longitudinally sectioned axon (A) has asplit that contains dark cytoplasm (asterisk) as well as a seconddefective area that has formed a balloon (b). Layer 4 in area 46 of a33-year-old monkey, AM 41. Scale bar � 1 �m.
284 A. PETERS AND C. SETHARES
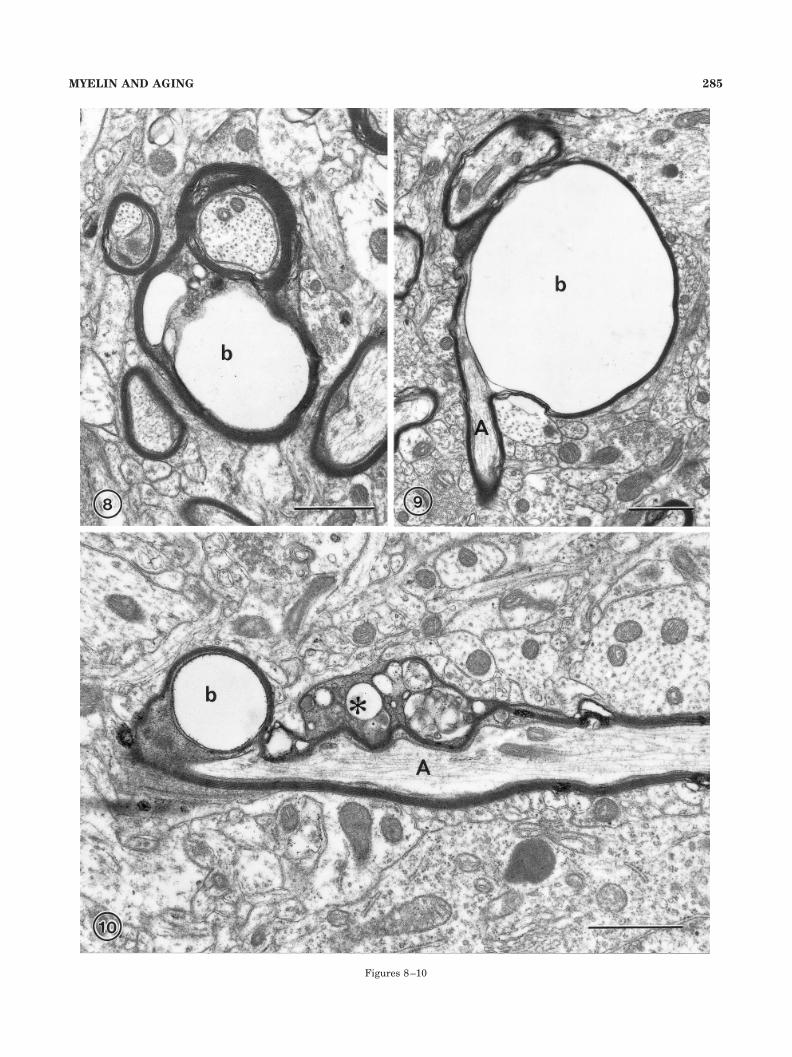
Figures 8–10
285MYELIN AND AGING

components that make up the CII, it is found that there isa correlation between the proportion of abnormal sheathsand the DNMS task with a 2-minute delay (r � 0.63, p �0.05) but no correlation with either the DNMS task with-out the delay or the delayed recognition memory span(DRS) task.
Age changes in axons
As pointed out, some axon profiles have a morphologythat differs from that shown by normal axons (Fig. 11).However, not more than approximately 0.5% of axons inthe old monkeys show age-related alterations and in mostof the monkeys the frequency is much lower, being be-tween 0.1 and 0.3%.
DISCUSSION
Myelin changes
The age-related changes in the myelin of area 46 aresimilar to those encountered in our earlier study of myelinin primary visual cortex (Peters et al., 2000). The mostcommonly occurring age-related alteration is the accumu-lation of dense material in splits of some of the myelinsheaths and, as pointed out previously, because this densecytoplasm is accommodated by splits of the major denseline, the dense cytoplasm must be derived from the oligo-dendrocytes. Moreover, because the dense cytoplasm inthe largest accumulations contain lysosomes, vesicles, andamorphous dense bodies, the cytoplasm appears to bedegenerating. Indeed, the dark cytoplasm resembles thatinduced in Cuprizone poisoning, in which it can be asso-ciated with oligodendroglial degeneration (Ludwin, 1978,1995). Dark cytoplasm is also present within the sheathsof mice with myelin-associated glycoprotein (MAG) defi-ciency (e.g., Lassmann et al., 1997; Biffiger et al., 2000).However, in the old monkeys, there are no indications thatthe oligodendroglia are degenerating, although they doaccumulate dense inclusions in their cell bodies and pro-cesses (Peters, 1996).
The myelin balloons that appear during aging can be aslarge as the cell bodies of neurons. They appear to be dueto the focal accumulation of fluid in splits of the intrap-eriod line (e.g., Ludwin, 1995; Feldman and Peters, 1998).In their review of cellular pathology, Kreutzberg et al.(1997) state that myelin sheath vacuolation is a form ofstatus spongiosis and suggest that it is due to exposure toa primary myelinotoxic agent. This agrees with the factthat such balloons have been described in a variety ofconditions such as spongiform degeneration in the gerbilcochlear nucleus (Faddis and McGinn, 1997), experimen-tal toxicity with Cuprizone (Ludwin, 1995), triethyl tin(Hirano, 1969; Malamud and Hirano, 1973), and by lyso-lecithin (Blakemore, 1978). Myelin balloons have alsobeen described in cases of severe diabetes (Tamura andParry, 1994), and they are formed in genetically alteredmice that lack or have increased proteolipid protein (e.g.,Monuki and Lemke, 1995; Anderson et al., 1998). They arealso present in mice deficient in galactolipid galactocere-broside. These mice develop apparently normal myelinsheaths that subsequently break down (Coetzee et al.,1996, 1998).
The so-called double sheaths that become increasinglycommon in old monkeys appear not to be due to the for-mation of one sheath around another but to an almost
complete separation of the inner from the outer lamellae,especially in thick sheaths of large diameter nerve fibers(Peters et al., 2001b). And, because thick sheaths becomeincreasingly common in aged monkeys, this presumablyleads to the increased incidence of apparently doublesheaths with age (Peters et al., 2001b). The other age-related myelin defect, namely the occurrence of sheathswith redundant myelin, seems to have been first noted bySturrock (1976) in the anterior commissures of aged mice,and Anderson et al. (1998) have noted that redundantmyelin sheaths are the earliest of the abnormalities en-countered in areas in which demyelination occurs in adultmice with an increased dosage of the proteolipid proteingene.
Nerve fiber changes
In old monkeys, only a small proportion of axons showalterations in their morphology, and there is no correla-tion between the ages of the monkeys and the frequency ofoccurrence of altered axons, either in area 46 and corpuscallosum, or in area 17 (Peters et al., 2000). But, there isa suggestion that some nerve fibers might be degenerat-ing, because in corpus callosum the packing density ofnerve fibers decreases with age, and occasional myelinfigures have been encountered in the cytoplasm of astro-cytes. However, unlike the optic nerves (Sandell and Pe-ters, 2001), there is no indication in the corpus callosumthat large numbers of nerve fibers are simultaneouslyundergoing degeneration with age, and for primary visualcortex, counts of nerve fibers in the vertical bundles sug-gest that significant numbers of nerve fibers are not lostwith age (Nielsen and Peters, 2000). This is surprising inview of the fact that axial MRI scans of both human(Albert, 1993; Guttmann et al., 1998) and rhesus monkey(Lai et al., 1995), as well as stereologic analyses (Pakken-berg and Gundersen, 1997; Tang et al., 1997), indicatethat some white matter is lost from the cerebral hemi-spheres during aging.
As we have seen in our examination of optic nerves fromsome of these monkeys (Sandell and Peters, 2001), thepathologic changes associated with nerve fiber degenera-tion are much more pronounced than any changes that wehave encountered in either frontal and visual cortex orcorpus callosum. Most of the optic nerves from old mon-keys show age-related changes in the myelin sheaths of
Fig. 11. A nerve fiber in which the axon (A) appears to be degen-erating, because it contains dense cytoplasm in which there are lyso-somes. Adjacent to this fiber is a profile of a myelin figure (asterisk)that surrounds vacuolated dark cytoplasm. This could be either adegenerating fiber or a balloon protruding from the sheath surround-ing the degenerating axon (A). Layer 4 in area 46 of a 35-year-oldmonkey, AM 13. Scale bar � 1 �m.
Fig. 12. An astrocytic process (As) containing of myelin figure (m),which suggests that the astrocyte has phagocytosed a degeneratingmyelin sheath. From layer 4 in area 46 of a 29-year-old monkey, AM26. Scale bar � 1 �m.
Fig. 13. In the corpus callosum of young monkeys, the nerve fibersare closely packed and their sheaths show no defects other than thoseproduced by localized shearing of lamellae (arrows). This field fromthe corpus callosum of a 10-year-old monkey, AM 53, contains anoligodendrocyte (O). Scale bar � 5 �m.
286 A. PETERS AND C. SETHARES
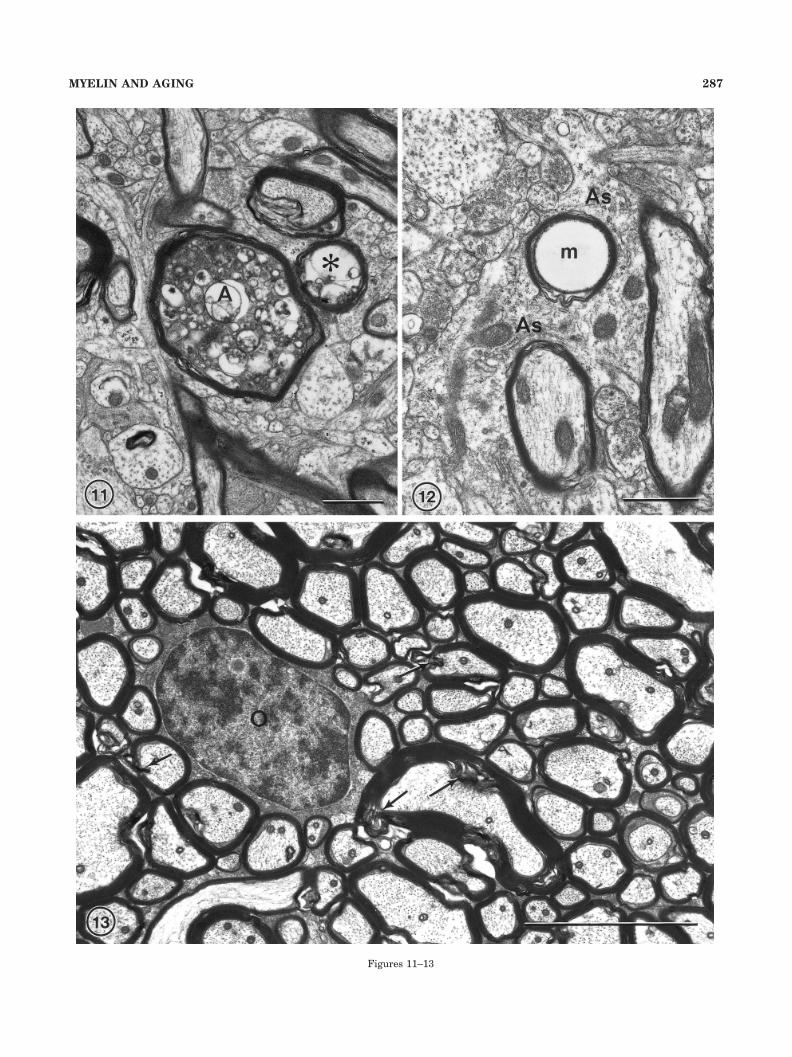
Figures 11–13
287MYELIN AND AGING
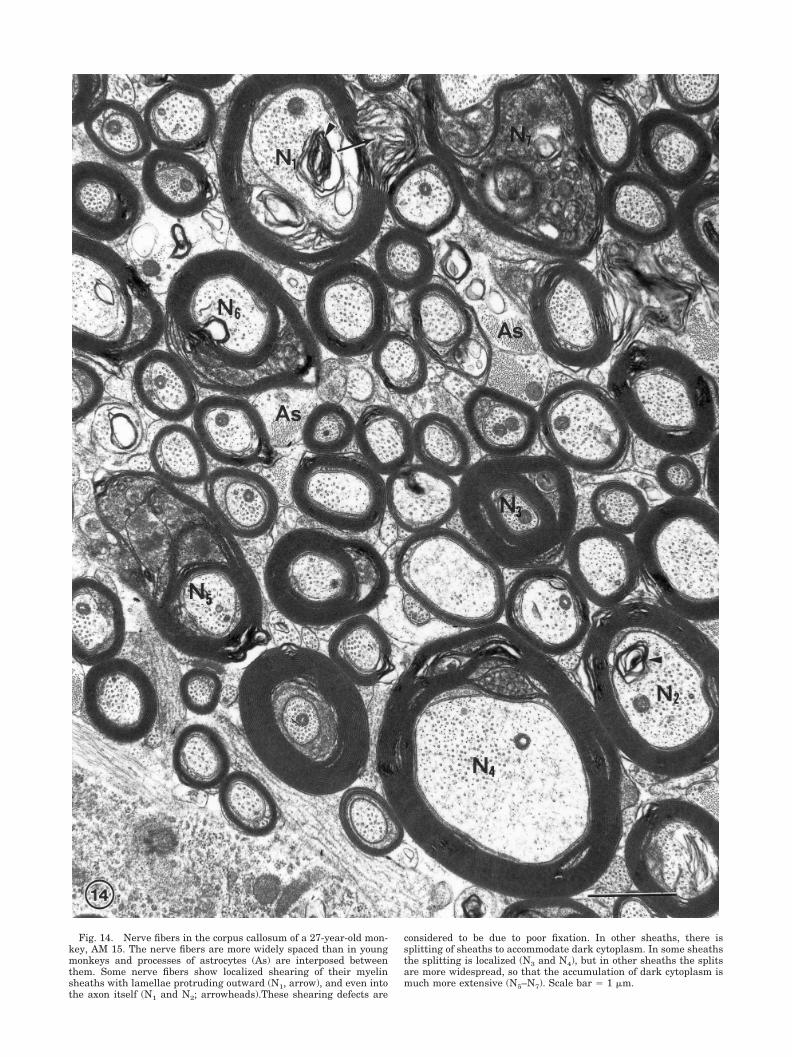
Fig. 14. Nerve fibers in the corpus callosum of a 27-year-old mon-key, AM 15. The nerve fibers are more widely spaced than in youngmonkeys and processes of astrocytes (As) are interposed betweenthem. Some nerve fibers show localized shearing of their myelinsheaths with lamellae protruding outward (N1, arrow), and even intothe axon itself (N1 and N2; arrowheads).These shearing defects are
considered to be due to poor fixation. In other sheaths, there issplitting of sheaths to accommodate dark cytoplasm. In some sheathsthe splitting is localized (N3 and N4), but in other sheaths the splitsare more widespread, so that the accumulation of dark cytoplasm ismuch more extensive (N5–N7). Scale bar � 1 �m.
their nerve fibers, and these changes are similar to thosereported here. But in addition, those optic nerves withextensive nerve fiber loss also show obvious degenerationof some axons and their myelin sheaths, and this is ac-companied by extensive phagocytosis of the degeneratingmaterial by activated microglial cells.
How these changes correlate with the observationsmade by Lintl and Braak (1983) and by Kemper (1994)that there is a decrease in the intensity of myelin stainingof nerve fibers with age is not clear. To them, the loss of
staining intensity suggested there is some loss of nervefibers with age, but it is equally possible that the decreasein myelin staining intensity reflects an alteration in struc-ture and composition of myelin with age, a question thatseems not to have been investigated.
It is also worth pointing out that, in humans, thereseems to be a correlation between some white matterchanges, such as myelin rarefaction and gliosis in thecerebral hemispheres, and cognitive decline (Inzitari,2000). Some of these white matter changes are visible inMRI scans, and de Groot et al. (2000) have recently exam-ined the correlation between the frequency of occurrenceof cerebral white matter lesions and cognitive function ofapparently normal elderly subjects in domains such asmemory and psychomotor speed. Their data suggest thatthe speed of cognitive processing seems to be most affectedby lesions in the periventricular white matter, where longassociation fibers are located and less affected by subcor-tical white matter lesions.
Alterations in sheaths and correlations withage and cognition
In both the present study of the effects of age on themyelin in layer 4 of area 46 and in corpus callosum, andthe previous study on the myelin sheaths in layer 4C ofarea 17 (Peters et al., 2000), correlations have been foundbetween the increasing frequency of altered myelinsheaths and age. All of the monkeys listed in Table 1, withthe exception of AM 23 and AM 100, were also included inthe study of age-related effects on myelin sheaths in area17 (Peters et al., 2000) or the frequency of myelin defectswithin those monkeys not included in that early study(i.e., AM 13, AM 58, AM 77, AM 96) have been determinedsubsequently. Consequently, the frequencies of myelinage-related changes in the two cortical areas and in corpuscallosum can be compared, each with the other. When thedata from areas 46 and 17 are compared, it is seen that, foreach monkey, the percentage of sheath profiles displayingage-related myelin changes is similar in both areas (r �0.87, p � 0.001). The correlation between frequency ofage-related changes in each of the cortical areas and in thecorpus callosum is also good (corpus callosum vs. area 46:r� 0.89, PI� 0.001; corpus callosum vs. area 17: r� 0.75,p � 0.01). This finding indicates that age-related alter-ations in myelin occur with comparable frequency in area46, area 17, and corpus callosum, and suggests the likeli-hood that with age alterations in myelin sheaths becomeincreasingly common throughout the entire brain.
In addition to the correlations with age, the data fromboth area 17 and area 46 on the frequency of occurrence ofsheaths defects correlates significantly with cognitive dys-function, as indicated by the Cognitive Impairment Index(CII). More importantly, when only the old monkeys over25 years of age are considered as a group, and the corre-lation between the cognitive impairment index and thefrequency of occurrence of age-related myelin alterationsis examined, for both area 46 (P� 0.001) and area 17 (P�0.001; Peters et al., 2000), there is a significant correlationbetween the two measures. However, for corpus callosum,there is only a significant correlation between the fre-quency of myelin defects and the behavior of the monkeyson the 2-minute delayed DNMS task. The DNMS task is arecognition memory task that has been used by severallaboratories to assess memory in aged monkeys and tocharacterize memory impairments in human subjects, and
Fig. 15. Plots of the percentages of normal sheath profiles in area46 and corpus callosum vs. the ages of the monkeys.
Fig. 16. Plots of the percentages of normal sheath profiles in area46 and corpus callosum against the Cognitive Impairment Indices(CII) of individual monkeys.
289MYELIN AND AGING

it is made more difficult by the imposition of the 2-minutedelay (e.g., see Moss et al., 1997; Herndon et al., 1997;Peters et al., 1998b; Killiany et al., 2000). The reason forthe difference between corpus callosum and the two corti-cal areas with respect to the correlations between myelinalterations and behavior is not known, but there are twoobvious factors that could affect any correlations. One isthat inferior fixation of the nerve fibers in corpus callosumin old monkeys can make it difficult to distinguish be-tween those defects caused by poor fixation and those dueto age. Another factor is that data about myelin defects inthe corpus callosum may be available from an insufficientnumber of monkeys to generate significant correlationswith the other behavioral tasks. But in any case, whenthey are considered in their entirety, the data stronglyindicate that age-related alterations in myelin sheathscontribute significantly to the cognitive decline evidencedby old monkeys.
Role of myelin sheaths in cognition
Myelin provides insulation to axons and makes salta-tory conduction possible. Consequently, it is likely thatdamage to myelin sheaths would decrease the rate ofconduction along nerve fibers, and there is evidence forsuch a decrease. Although the structure of the myelin wasnot examined, there are several recorded examples of re-duced conduction velocity in old animals. Thus, Aston-Jones et al. (1985) found that, in rats, the conductionlatencies from nucleus basalis to frontal cortex are signif-icantly longer in aged than in young animals, and theysuggest that impairment in the fidelity of the nucleusbasalis-cortical system could disrupt normal cognitivemechanisms. Morales et al. (1987) have also shown areduction in the conduction velocities of lumbar motoneu-rons in the spinal cords of old cats, whereas Xi et al. (1999)found that, in old cats, only 26% of pyramidal tract neu-rons are fast conducting, compared with 51% in youngcats. Xi et al. (1999) also records that, in old cats, there isan overall reduction of 43% in the median conductionvelocity of these neurons. Other examples of myelinchanges altering conduction rates, have been provided byGutierrez et al. (1995), who have shown that, in proteo-lipid protein (PLP) deficient mice, the decompaction ofmyelin leads to a reduced conduction velocity. A decreasein conduction velocity also occurs in demyelinating dis-eases (see Waxman et al., 1995; Felts et al., 1997).
The available data suggest that the age-related changesin myelin sheaths may be ubiquitous, because we havenow shown such changes to occur in prefrontal cortex andcorpus callosum, visual cortex (Peters et al., 2000), parscompacta of the substantia nigra (Siddiqi and Peters,1999), and optic nerve (Sandell and Peters, 2000). Becausethere is evidence that age-related changes in the structureof myelin lead to a reduction in conduction velocities alongthe affected nerve fibers, it can be conjectured that thiswould lead to changes in the timing of sequential events inneuronal circuits. Consequently, it is proposed that suchdisruptions, produced by the age-related alteration in my-elin sheaths, offer one basis for the cognitive decline ex-hibited by older primates.
ACKNOWLEDGMENTS
We thank Drs. Ron Killiany, Jim Herndon, and MarkMoss for providing the behavioral data on the effects of
age on the cognition of monkeys used in this study, andDr. Douglas Rosene for perfusing the monkey brains formicroscopic examination. We also thank Dr. Howard Ca-bral for his comments on the appropriate statistical anal-yses to be used in this study.
LITERATURE CITED
Albert M. 1993. Neuropsychological and neurophysiological changes inhealthy adult humans across the age range. Neurobiol Aging 14:623–625.
Albert M, Moss MB. 1996. Neuropsychology of aging: findings in humansand monkeys. In: Schneider EL, Rowe JW, Morris JH, editors. Hand-book of the biology of aging. 4th ed. San Diego, CA: Academic Press. p217–233.
Anderson TJ, Schneider A, Barrie LA, Klugman M, McCulloch MC, Kirk-man D, Kyriades E, Nave KA, Griffiths IR. 1998. Late onset neurode-generation in mice with increased dosage of the proteolipid proteingene. J Comp Neurol 394:506–519.
Arnsten AFT, Goldman-Rakic PS. 1990. Analysis of alpha-2 adrenergicagonist effects on the delayed nonmatching to sample performance ofaged rhesus monkeys. Neurobiol Aging 11:583–590.
Aston-Jones G, Rogers J, Shaver RD, Dinan TG, Moss DE. 1985. Age-impaired impulse flow from nucleus basalis to cortex. Nature 318:462–464.
Bachevalier J, Landis LS, Walker LC, Brickson M, Mishkin M, Price DL,Cork LC. 1991. Age monkeys exhibit behavioral deficits indicative ofwidespread cerebral dysfunction. Neurobiol Aging 12:99–111.
Biffiger K, Bartsch S, Montag D, Aguzzi A, Schachner M, Bartsch U. 2000.Severe hypomyelination of the murine CNS in the absence of myelin-associated glycoprotein and Fyn tyrosine kinase. J Neurosci 20:7430–7437.
Blakemore WF. 1978. Observations on remyelination in the rabbit spinalcord following demyelination induced by lysolecithin. NeuropatholAppl Neurobiol 4:47–59.
Brodmann K. 1905. Beitrage zur histologischen Lokalization derGrosshirnrinde. IIIte Mittlung: Die Rindfelder der niederen Affen.J Psychol Neurol 4:177–226.
Coetzee T, Fujita N, Dupree J, Shi R, Blight A, Susuki K, Popko B. 1996.Myelination in the absence of galactocerebroside and sulfatide: normalstructure with abnormal function and regional instability. Cell 86:209–219.
Coetzee T, Susuki K, Popko B. 1998. New perspectives on the function ofmyelin galactolipids. Trends Neurosci 21:126–130.
De Groot JC, de Leeuw F-E, Oudkerk M, van Gijn J, Hofman A, Jolles J,Breteler M. 2000. Cerebral white matter lesions and cognitive function:the Rotterdam scan study. Ann Neurol 47:145–151.
Faddis BT, McGinn MD. 1997. Spongiform degeneration of the gerbilcochlear nucleus: an ultrastructural and immunohistochemical evalu-ation. J Neurocytol 26:625–635.
Feldman ML, Peters A. 1998. Ballooning of myelin sheaths in normallyaged macaques. J Neurocytol 27:605–614.
Felts PA, Baker TA, Smith KJ. 1997. Conduction along segmentally de-myelinated mammalian central axons. J Neurosci 17:7267–7277.
Goldman PS, Rosvold HE, Vest B, Galkin TW. 1971. Analysis of thedelayed-alteration deficit produced by dorsofrontal lesions in the rhe-sus monkey. J Comp Physiol Psychol 77:212–220.
Goldman-Rakic PS. 1996. The prefrontal landscape: implications of func-tional architecture for understanding human mentation and the cen-tral executive function. Philos Trans R Soc Lond Biol Sci 351:1445–1453.
Gutierrez R, Bioson D, Heinemann U, Stoffel W. 1995. Decompaction ofCNS myelin leads to a reduction of the conduction velocity of actionpotentials in optic nerve. Neurosci Lett 195:93–96.
Guttmann CRG, Jolesz FA, Kikinis R, Killiany RJ, Moss MB, Sandor T,Albert MS. 1998. White matter changes with normal aging. Neurology50:972–978.
Herndon J, Moss MB, Killiany RJ, Rosene DL. 1997. Patterns of cognitivedecline in early, advanced and oldest of the old aged rhesus monkeys.Behav Res 87:25–34.
Hirano A. 1969. The fine structure of the brain in edema. In: Bourne GH,editor. The structure and function of nervous tissue. Vol. 2. New York:Academic Press. p 69–135.
Hof RP, Nimchinsky EA, Young W, Morrison JH. 2000. Numbers of Mey-
290 A. PETERS AND C. SETHARES

nert and layer IVB cells in area V1; stereological analysis in young andaged macaque monkeys. J Comp Neurol 420:113–126.
Inzitari D. 2000. Age-related white matter changes and cognitive impair-ment. Ann Neurol 47:141–143.
Kemper TL. 1994. Neuroanatomical and neuropathological changes duringaging and dementia. In: Albert ML, Knoefel JE, editors. Clinical neu-rology of aging. New York: Oxford University Press. p 3–67.
Killiany RJ, Moss MB, Rosene DL, Herndon J. 2000. Recognition memoryfunction in early senescent rhesus monkeys. Psychobiology 28:45–56.
Kim CBY, Pier LP, Spear PD. 1997. Effects of aging on numbers and sizesof neurons in histochemically defined subregions of monkey striatecortex. Anat Rec 247:119–128.
Kreutzberg GW, Blakemore WF, Graeber MB. 1998. Cellular pathology ofthe central nervous system. In: Graham DI, Lantos PL, editors. Green-field’s neuropathology. 6th ed. London: Arnold. p 85–156.
Lai ZC, Rosene DL, Killiany RJ, Pugliese D, Albert MS, Moss MB. 1995.Age-related changes in the brain of the rhesus monkey: MRI changes inwhite matter but not gray matter. Soc Neurosci Abstr 21:1564.
Lassmann H, Bartsch U, Montag D, Schachner M. 1997. Dying-back oli-godendrogliopathy: a late sequel of myelin-associated glycoprotein de-ficiency. Glia 19:104–110.
Lintl P, Braak H. 1983. Loss of intracortical myelinated fibers: a distinctivealteration in the human striate cortex. Acta Neuropathol 61:178–182.
Ludwin SK. 1978. Central nervous system demyelination and remyelina-tion in the mouse. An ultrastructural study of Cuprizone toxicity. LabInvest 39:597–612.
Ludwin SK. 1995. Pathology of the myelin sheath. In: Waxman SG, KocsisJD, Stys PK, editors. The axon: structure, function and pathophysiol-ogy. New York: Oxford University Press. p 412–437.
Malamud N, Hirano A. 1973. Atlas of neuropathology. Berkeley: Univer-sity of California Press.
Monuki ES, Lemke G. 1995. Molecular biology of myelination. In: WaxmanSG, Kocsis JD, Stys PK, editors: The axon: structure, function andpathophysiology. New York: Oxford University Press. p 144–163.
Morales FR, Boxer PA, Fung SJ, Chase MH. 1987. Basic electrophysiolog-ical properties of spinal cord motoneurons during old age in the cat.J Neurophysiol 58:180–194.
Morrison JH, Hof PR. 1997. Life and death of neurons in the aging brain.Nature 278:412–419.
Moss MB, Killiany RJ, Lai ZC, Rosene DL, Herndon JG. 1997. Recognitionspan in rhesus monkeys of advanced age. Neurobiol Aging 18:13–19.
Moss MB, Killiany RJ, Herndon JG. 1999. Age-related cognitive decline inrhesus monkey. In: Peters A, Morrison JH, editors. Neurodegenerativeand age-related changes in structure and function of the cerebralcortex. Cerebral cortex. Vol. 14. New York: Kluwer Academic/PlenumPubishers. p 21–48.
Nielsen K, Peters A. 2000. The effects of aging on the frequency of nervefibers in rhesus monkey striate cortex. Neurobiol Aging 21:621–628.
Pakkenberg B, Gundersen HJG. 1997. Neocortical neuron number in hu-mans: effect of sex and age. J Comp Neurol 384:312–320.
Peters A. 1991. Aging in monkey cerebral cortex. In: Peters A, Jones EG,editors. Normal and altered states of function. cerebral cortex. Vol. 9.New York: Plenum Press. p 485–510.
Peters A. 1996. Age-related changes in oligodendrocytes in monkey cere-bral cortex. J Comp Neurol 371:153–163.
Peters A. 1999. Normal aging in the cerebral cortex of primates. In: PetersA, Morrison JH, editors: Neurodegenerative and age-related changes instructure and function of cerebral cortex. Cerebral cortex. Vol. 14. NewYork: Kluwer Academic/ Plenum Publishers. p 49–80.
Peters A, Sethares C. 1993. Aging and the Meynert cells in rhesus monkeyprimary visual cortex. Anat Rec 236:721–729.
Peters A, Josephson K, Vincent SL. 1991. Effects of aging on the neuroglialcells and pericytes within area 17 of the rhesus monkey cerebral cortex.Anat Rec 229:384–398.
Peters A, Leahu D, Moss MB, McNally KJ. 1994. The effects of aging onarea 46 of the frontal cortex of the rhesus monkey. Cereb Cortex6:621–635.
Peters A, Rosene DL, Moss MB, Kemper TL, Abraham CR, Tigges J, AlbertMS. 1996. Neurological bases of age-related cognitive decline in therhesus monkey. J Neuropathol Exp Neurol 55:861–874.
Peters A, Nigro NJ, McNally KJ. 1997. A further evaluation of the effectsof age on striate cortex of the rhesus monkey. Neurobiol Aging 18:29–36.
Peters A, Morrison JH, Rosene DL, Hyman BT. 1998a. Are neurons lostfrom the primate cerebral cortex during aging? Cereb Cortex 8:295–300.
Peters A, Sethares C, Moss MB. 1998b. The effects of aging on layer 1 inarea 46 of prefrontal cortex in the rhesus monkey. Cereb Cortex 8:671–684.
Peters A, Moss MB, Sethares C. 2000. The effects of aging on myelinatednerve fibers in monkey primary visual cortex. J Comp Neurol 419:364–376.
Peters A, Moss MB, Sethares C. 2001a. The effects of aging on layer 1 ofprimary visual cortex in the rhesus monkey. Cereb Cortex 11:93–103.
Peters A, Sethares C, Killiany RJ. 2001b. Effects of age on the thickness ofmyelin sheaths in monkey primary visual cortex. J Comp Neurol 435:241–248.
Petrides M. 1996. Specialized systems for the processing of mnemonicinformation within the primate frontal cortex. Philos Trans R Soc LondB Biol Sci 351:1455–1461.
Presty SK, Bachevalier J, Walker LC, Struble RG, Price DL, Mishkin M,Cork LC. 1987. Age differences in recognition memory of the rhesusmonkey (Macaca mulatta). Neurobiol Aging 8:435–440.
Rapp PR, Amaral DG. 1989. Evidence for a task-dependent memory dys-function in the aged monkey. J Neurosci 9:3568–3576.
Sandell JH, Peters A. 2001. Effects of age on nerve fibers in the rhesusmonkey optic nerve. J Comp Neurol 429:541–553.
Siddiqi ZA, Peters A. 1999. The effects of aging on pars compacta of thesubstantia nigra in rhesus monkey. J Neuropathol Exp Neurol 58:903–920.
Sloane JA, Pietropaolo MF, Rosene DL, Moss MB, Peters A, Kemper T,Abraham CR. 1997. Lack of correlation between plaque burden andcognition in the aged monkey. Acta Neuropathol 94:471–478.
Sturrock RR. 1976. Changes in neuroglia and myelination in the whitematter of aging mice. J Gerontol 31:513–522.
Tamura E, Parry GJ. 1994. Severe radicular pathology in rats with long-standing diabetes. J Neurol Sci 127:29–35.
Tang Y, Nyengaard JR, Pakkenberg B, Gundersen HJG.1997. Age-inducedwhite matter changes in the human brain: a stereological investigation.Neurobiol Aging 18:609–615.
Tigges J, Gordon TP, McClure HM, Hall EC, Peters A. 1988. Survival rateand life span of rhesus monkeys at the Yerkes Regional Primate Re-search Center. Am J Primatol 15:263–272.
Tigges J, Herndon JG, Peters A. 1990. Neuronal population of area 4during life span of the rhesus monkey. Neurobiol Aging 11:201–208.
Vincent SL, Peters A, Tigges J. 1989. Effects of aging on the neurons withinarea 17 of rhesus monkey cerebral cortex. Anat Rec 223:329–341.
Walker AE. 1940. A cytoarchitectural study of the prefrontal area of themacaque monkey. J Comp Neurol 73:59–86.
Waxman SG, Kocsis JD, Black JA.1995. Pathophysiology of demyelinatedaxons. In: Waxman SG, Kocsis JD, Stys PK, editors. The axon: struc-ture, function and pathophysiology. New York: Oxford UniversityPress. p 438–461.
Xi M-C, Liu R-H, Engelhardt KK, Morales FR, Chase MH. 1999. Changesin the axonal conduction velocity of pyramidal tract neurons in theaged cat. Neuroscience 92:219–225.
291MYELIN AND AGING