age of tertiary formations on spitsbergen
TRANSCRIPT

Age of Tertiary formations SVEIN B. MANUM AND TORBJBRN THRONDSEN
on Spitsbergen
Manum, S. B. & Throndsen, T. 1986: Age of Tertiary formations on Spitsbergen. Polar Research 4 n.s., 103-131.
Biostratigraphic evidence for the age of Tertiary formations on Spitsbergen is critically reviewed and new evidence from dinoflagellates is presented. The shift in basin geometry and sediment source area observed in the Gilsonryggen Formation is dated as latest Palaeocene, coinciding with the initial opening of the Norwegian Sea at anomaly 24-25 time. An Upper Eocene age is proposed for sediments from Forlandsundet (Sarsbukta), indicating that the Forlandsundet graben was created during the phase of shear movement between Greenland and Spitsbergen before anomaly 13 time.
Svein B. Manum, Department of Geology, University of Oslo, P. 0. Box 1047, Blindem, N-0316 Oslo 3, Norway; Torbj0rn Throndsen, Institute for Energy Technology, P. 0. Box 40, N-2007 Kjeller, Norway; February 1986 (revised August 1986).
Tertiary sediments cover the greater part of Spits- bergen south of Isfjorden (Fig. 1). They are pre- served in a north-south trending trough with an estimated maximum thickness exceeding 2300 metres (Harland et al. 1976). Lithostratigraph- ically they are divided into six formations (Major & Nagy 1972), starting with deltaic coal-bearing strata resting unconformably on late Lower Cre- taceous sediments (Fig. 2j. The succession con- tinues through shallow marine deposits followed by deeper marine sediments and ends with mass- ive deltaic sandstones including some coals (Steel et al. 1981). There is evidence from vitrinite reflectance studies that approximately 1500-2000 metres of sediments have overlain the presently preserved formations (Manum & Throndsen 1978).
Outside and to the west of the central basin of Mesozoic and Tertiary sediments, there are four small areas with Tertiary rocks preserved: For- landsundet, Ny Alesund, Renardodden, and 0yr- landet. Opinions vary considerably with regard to the correlation of these isolated outcrops with the formations of the central basin.
The age of the sediments has been recognized as Tertiary ever since the rich angiosperm floras of the basal and top formations of the central basin (and also of some of the isolated areas) were first discovered (Heer 1868, 1870, 1876; Nathorst 1910). However, more precise age deter- minations for these formations have remained controversial to the present day. The main reason for this is the poor yield of biostratigraphically
diagnostic marine fossils in spite of impressive field and laboratory efforts by many scientists for more than a century.
There is now a greater need than ever before to solve the age problem of the Spitsbergen Tertiary. Recent attempts to correlate the observed changes in depositional environments with the established plate-tectonic events in the area suffer from lack of precise age determinations for the sediments (Steel et al. 1986). Moreover, the Ter- tiary events and the geology of Spitsbergen on the whole have recently attracted renewed interest because of Spitsbergen’s key role as reference area for the off-shore geology of the Barents Sea.
In this paper we offer firstly a review of the status regarding the dating of the Tertiary for- mations, and secondly we present the results of our own efforts over more than a decade to estab- lish a better biostratigraphy based on marine palynomorphs.
Material and Methods Sample locations
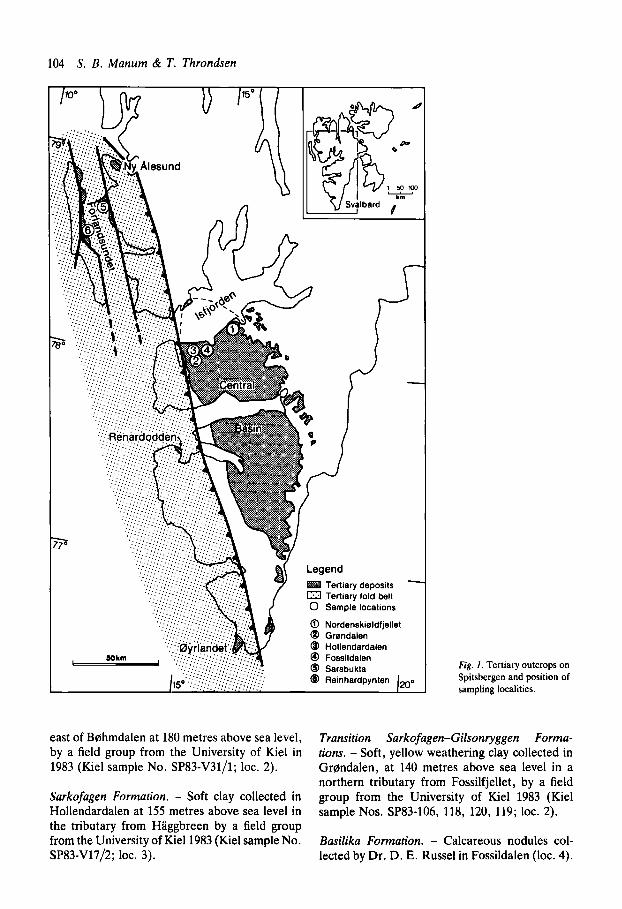
Locality numbers used below refer to Fig. 1.
Gilsonryggen Formation. - Soft, yellow weath- ering clay, probably an ash-layer, collected by T. Throndsen 30 metres above the base of the formation at Nordenskiddfjellet (loc. 1).
Similar lithology, collected in Gr~ndalen, 1 km
104 S. B. Manum & T. Throndsen
east of Bohmdalen at 180 metres above sea level, by a field group from the University of Kiel in 1983 (Kiel sample No. SP83-V31/1; loc. 2).
Sarkofagen Formation. - Soft clay collected in Hollendardalen at 155 metres above sea level in the tributary from Haggbreen by a field group from the University of Kiel1983 (Kiel sample No. SP83-V17/2; loc. 3).
Fig. 1. Tertiary outcrops on Spitsbergen and position of sampling localities.
Transition Sarkofagen-Gilsonryggen Forma- tions. - Soft, yellow weathering clay collected in Grmdalen, at 140 metres above sea level in a northern tributary from Fossilfjellet, by a field group from the University of Kiel 1983 (Kiel sample Nos. SP83-106, 118, 120, 119; loc. 2).
Basilika Formation. - Calcareous nodules col- lected by Dr. D. E. Russel in Fossildalen (loc. 4).

STEE
L €l
M.
1981
M
AJOR
AN0
NAGY
197
2
ASPE
LINT
OPP
EN
FM
BA
llFJE
UE
T FM
GILS
ONRY
GGEN
FM
SARK
OFAG
EN
FM
BA
SlLl
U
FM
FlRl
UllT
EW
FM
HEER
186
8.70
.76
NATH
ORST
191
0
plan
ts
MIO
CENE
MIO
CENE
RAVN
192
2
mol
lusc
s
EOCE
NE ?
+ M
IOO
LE
I- UP
PER?
) P
AU
EO
EN
E
I V
AK
UW
KO
. UVsnnS 1
970.
197
4 OS
ENKR
AIITZ
194
2 M
AN
UM
196
2 lO
NOER
8ANK
197
0
PI- 1-1 UTE
EOCE
NE
dlm
u
WO
CE
NE
EWEM
E
Fig.
2. L
ithos
trat
igra
phy,
dep
ositi
onal
env
iron
men
ts a
nd b
iost
ratig
raph
ic e
vide
nce f
or a
ges o
f th
e ce
ntra
l bas
in T
ertia
ry s
eque
nce o
n Sp
itsbe
rgen
.

106 S. B . Manum & T. Throndsen
Forlandsundet. :c Micaceous clay from Sarsbukta, collected by R. W. Feyling-Hanssen (F.-H- sample Nos. 248 and 260, locality description in Feyling-Hanssen & Ulleberg 1984); samples from same locality collected by T. Throndsen (loc. 5).
Reinhardpynten. - Black shale samples collected by T. Throndsen (loc. 6).
Laboratory treatment
Standard palynological processing, using hydro- chloric and hydrofluoric acids to dissolve mineral components, was used. Oxidation was occasion- ally used to remove pyrite and to clear the residue of amorphous matter.
Sample productivity and palynomorph preservation Our search for marine palynomorphs that could aid the dating of the Tertiary sequence has been going on since the first dinoflagellates were dis- covered in these sediments (Manum 1960, Sarsbukta loc. 5). Hundreds of samples have been processed, although mostly with disappointing results. If samples were at all productive, pres- ervation was usually too poor to allow identi- fications of stratigraphic significance. Only a few samples have yielded diagnostic palynomorphs, and the present paper aims at documenting the most reliable records of dinoflagellates that we have been able to make so far.
The most useful palynological samples from the central basin have been collected from relatively thin and soft, usually yellowish weathering clay beds. They are apparently of volcanic ash origin (Dypvik & Nagy 1979). Ash deposition appears to have provided an environment that has been less destructive to palynomorphs. However, also in such samples preservation is highly variable. A typical degradation feature is a bubble-like structure which severely distorts the fossils’ origi- nal morphology. Dinoflagellates have also been obtained from calcareous nodules of the Basilika Formation; these apparently have suffered little compression, indicating that the nodules were formed prior to compaction of the sediments. Specimens from this lithology, however, appear to have thinner walls than normal for the species present, and fragmentation is common.
In the samples from Sarsbukta - one of the localities outside the central basin - the pre- servation is considerably better and compression is negligible. The palynomorph productivity, however, is not particularly high and only a few specimens have been found of most of the species recorded. On the other hand, some of the samples from the central basin are quite productive, but even in the richer ones well preserved specimens are not very common. The fossils are often highly compressed and folded and otherwise degraded.
Previous age determinations No radiometric age determinations for the Spits- bergen Tertiary exist so far. Biostratigraphic cor- relations can be made only with sequences that occur at considerably lower latitudes. While making such correlations one always assumes that the biostratigraphic criteria are valid over great palaeo-latitudinal spans. However, existing dis- crepancies between ages proposed by various authors for the Spitsbergen Tertiary indicate that the criteria are not always reliable. The incon- sistencies are particularly noticeable for ages based on terrestrial floras. However, there are also problems with marine fossils; this is clearly demonstrated by our results from Forlandsundet which differ from previous conclusions based on foraminifera.
The lithostratigraphic terminology proposed by Major & Nagy (1972) for sediments of I he central basin has been adopted in this paper. Correlation to other lithostratigraphic schemes for the same sediments poses no serious problem, and we refer the reader to the correlation table in Major & Nagy (1972, p. 22).
The central basin (Fig. 2 )
Heer (1868,1870,1876) described rich floras from the basal and topmost formations (Firkanten and Aspelintoppen respectively) which he assigned to the Miocene because of their similarity to Swiss Miocene floras which he knew well. This age was also adopted by the Swedish explorer, geologist and palaeobotanist A. G . Nathorst in the first comprehensive monograph on the geology of the Svalbard archipelago to which Spitsbergen belongs (Nathorst 1910).
The concept of a Miocene age remained unchal- lenged until Ravn (1922) proposed a Palaeocene

Age of Tertiary formations 107
made earlier by Rosenkrantz (1942) based on the find of the bivalve Thyasira (Conchocele) conradii in the Basilika Formation, a species which has been recorded in Greenland in strata referred to the Danian.
Palynological studies by Manum (1962), par- ticularly of pollen and spores from coals in the basal and topmost units, offered no more precise age determinations than those of Ravn (1922). They appear, however, to support a Palaeocene to Eocene age for the entire sequence. The macro- as well as microfloras of the basal and the topmost units were not seen to be significantly different, suggesting that they are not of widely different ages.
In more recent years, Soviet scientists (Vak- ulenko & Livshits 1971; Livshits 1974) have contributed considerably to the palaeontology of the Spitsbergen Tertiary. Vakulenko & Livshits (1971) identified pollen and spores from all for- mations except the Basilika Formation, and a correlation to Soviet Arctic Tertiary was made on the basis of identifications of spore/pollen ‘complexes’. A Palaeocene complex was ident- ified in the Firkanten Formation, an Oligocene or possibly even Miocene complex was reported for the Aspelintoppen Formation, and Eocene was indicated for the intervening units. However, Vakulenko & Livshits (loc. cit.) were also con- strained in their conclusions by poor preservation of the palynomorphs. This is reflected by the fact that out of more than 50 recorded pollen and spore taxa, only six were identified beyond the generic level. They also stated that the differences between the three identified complexes were not very pronounced. Their Oligocene or possibly Miocene age for the youngest formation was based on the somewhat impoverished palynoflora recovered from it. However, Manum (1962) con- cluded that there appeared to be no biostrati- graphically significant differences between the macrofloras of the youngest and oldest units. The reduced palynoflora noticed in the youngest unit by Vakulenko & Livshits (loc. cit.) may well reflect different preservational and depositional conditions rather than real differences in the flora.
Livshits (1974) also reported the results of stud- ies of plant macrofossils and of molluscs on the basis of which a Palaeocene age was indicated for the Firkanten Formation. For the Aspelintoppen Formation, the molluscs, identified only to gen- eric level, were considered to indicate an Oli- gocene age; the plants, on the other hand, were
to Eocene age for the lower parts of the sequence based on marine and fresh-water molluscs, par- ticularly from the Sarkofagen and Battfjellet For- mations. In spite of the fact that Ravn expressed considerable reservation with regard to his age indications, mainly because of the poor quality of his specimens, his conclusion quickly replaced the palaeobotanically founded concept of a Miocene age for the sequence. Ravn’s determination of ‘a Middle Palaeocene or perhaps Upper Palaeocene age’ for the Sarkofagen Formation was based on the identification of a single bivalve species, Cyrena (Corbicula) angustidens, which allowed him to make comparisons with identifications from the Palaeogene of France and Belgium. Other fossils from the Sarkofagen Formation could only be referred to the generic level or else to a new taxon. The fossils from the Battfjellet Formation were even more difficult to determine, and Ravn indicated an Eocene age with great reservations. It needs mentioning here that the stratigraphic significance of Ravn’s identification of Corbicula angustidens has subsequently been played down considerably by Vonderbank (1970, p. 100). The study by Ravn (1922) offers a good example of the problems that palaeontologists and biostratigraphers have faced in the Spits- bergen Tertiary. Owing to poor preservation it becomes difficult to take identifications further than to the generic level, and when species can be distinguished, they are often described as new taxa. Obviously, this provides a poor basis for stratigraphic conclusions, and these therefore tend to lack precision.
Vonderbank (1970) studied extensive col- lections of foraminifera, molluscs, and crus- taceans from several formations, but he was also severely limited in his taxonomic and stratigraphic conclusions by unsatisfactory preservation. Only 30 of the 72 taxa of foraminifera listed by Von- derbank were identified to known species, and these were of little stratigraphic significance. The crustacean remains were referred to a new species, as yet endemic. The biostratigraphically most significant among Vonderbank’s identi- fications is one bivalve (Modiolus hauniensis) and two gastropod species (Cylichna discifera and Chenopus gracilis) from the Basilika Formation (based on fragmented or otherwise damaged specimens). These species occur in the Danian in Denmark (‘Faxekalk’), and on this basis Von- derbank proposed a Danian age for the Basilika Formation. This age agrees with a suggestion

108 S. B . Manum & T. Throndsen
UVSHITS 1014
UPPER EOCENE proposed wr- relation with Gilsonryggen
Frn
OLIGOCENE 7
proposed wr- relation wtth
Aspelintoppen '
Fm
EOCENE/ OLIGOCENE
FORLANDSUNDET ( SARSBUUA ) dinoflagellates
THlEOlG L II. 1900
proposed cor- relation wlth Firkanten Fm
molluscs
7
FEVUNQ- HANSSEN & U 1 LE B E R G
1W MIDDLE/
I P E R OLIGOCENE
foraminifera
UPPER EOCENE
dinoflagellates
UPPER EOCENE / LOWER OLIGOCENE
dinoflagellates
Fig. 3. Stratigraphic assignments for Tertiary outcrops outside the central basin (only papers presenting new biostratigraphic evidence have been includedl
considered to indicate an age no younger than late Eocene.
Thus, to summarize the biostratigraphic statfls for the formations of the central basin, an early Tertiary age (Danian/Palaeocene) appears to be generally well agreed upon for the two oldest formations. For the youngest formations, however, the age indications range from Eocene to Oligocene, and possibly even Miocene; for the intervening formations evidence is indeed vague or lacking.
Areas outside the central basin (Fig. 3) Plant macrofossils are common and coal seams occur at the Bellsund locality (Renardodden) and in the Ny Alesund (Kings Ray) area. The floras do not seem to differ significantly from those found in the basal and topmost formations of the central basin. Unfortunately, they are of very little biostratigraphic value.
Renardodden. - Heer (1876) and subsequently Nathorst (1910) found the flora from Renard- odden to be most similar to that of the Aspe- lintoppen Formation of the central basin. Livshits (1974) arrived at a similar conclusion based on the pollen and spores recovered from the sequence and proposed an Oligocene age. More recently, Head (1984) indicated a Late Eocene to Early Oligocene age based on dinoflagellates, which would also appear to support a correlation with the uppermost plant-bearing formation of the central basin (the Aspelintoppen Formation) rather than with the basal formation. (Head listed
among others the following dinoflagellates: Areo- sphaeridium arcuatum, A. diktyoplokus, Cer- atiopsis depressa, Cordosphaeridium funiculatum, Corrudinium incompositum, Deflandrea spinu- losa, Glaphyrocysta semitecta, 'Millioudodinium' giuseppei, Phthanoperidinium comatum, P . echi- natum, P. geminatum, Wetzeliella articulata, W. oualis, and W. symmetrica.) Thiedig et al. (1980), on the other hand, considered a correlation with the lower parts of the central basin more likely, based on their finding of Conchocele conradii, a mollusc species first reported by Rosenkrantz (1942) in the Basilika Formation (cf. the discus- sion above of Vonderbank's study on this forma- tion). In view of the available evidence, we would tend to favour Head's dinoflagellate-based cor- relation of the Renardodden sequence rather than rely on a single mollusc species whose range appears uncertain.
N y Alesund. - The Tertiary sequence of the Ny hesund area is generally considered to be a northerly extension of the central basin (Hjelle & Lauritzen 1982). However, there is no other biostratigraphic evidence available than the ter- restrial flora, which is inconclusive. Vakulenko & Livshits (1971) correlated the sequence with the Aspelintoppen Formation without offering new biostratigraphic evidence.
Sarsbukta. - Manum (1960) reported the first find of dinoflagellates in the Spitsbergen Tertiary from the Sarsbukta locality at Forlandsundet; these were indicative of an Eocene to Oligocene age. The preservation of palynomorphs at this locality

Age of Tertiary formations 109
Apectodinium homomorphum (Deflandre & Cookson) L. & W. P1. 2, Fig. 2. Specimens corresponding to the illustrations in Harland (1979, P1. 1, Figs. 1-8) are recorded under this species. However, there appears to be a range of variability present (e.g. P1. 2, Fig. 6), corresponding to the range in morphology exhibited by the A . homomorphum-A. sumis- sum-A. quinquelatum group of species (cf. Har- land 1979).
was found to be better than in the formations of the central basin, and a few plant taxa were recorded which were unknown from the central basin (e.g. Tsuga, Manum 1962). Recently, Feyling-Hanssen & Ulleberg (1984) reported for- aminifera in Tertiary samples from Sarsbukta. They concluded that the sequence is ‘most prob- ably Middle to Upper Oligocene in age’ (op. cit. p. 83). The same sequence was proposed by Livshits (1974) to be of Upper Eocene age and to correlate with the Gilsonryggen Formation, based on a combination of faunistic and floristic evidences, although not particularly decisive.
0yrlandet. -No biostratigraphic evidence is avail- able from this locality.
Dinoflagellates Dinoflagellate taxa are listed below in alpha- betical order. The nomenclature follows Lentin & Williams (1985) and we also refer to their list of references for author citations.
Division Pyrrhophyta Class Dinophyceae Order Peridiniales Apectodinium Costa & Downie
Apectodinium augustum (Harland) L. & W. P1. 1, Figs. 1-6; P1. 2, Fig. 1 This species forms a conspicuous component of samples from the Gilsonryggen Formation. Indi- viduals present a range in variability much like that observed by Harland (1979). Some specimens show finer processes than the typical A . augustum (Pl. 1, Fig. 6 and PI. 2, Fig. 1, the latter appears to approach A . paniculatum).
Spitsbergen record. - Gilsonryggen Fm., localities 1. 2.
Stratigraphic range. - Latest Palaeocene, NP9 equivalent (Harland 1979; Costa & Downie 1979b: Into NP10).
Spitsbergen record. - Gilsonryggen Fm., locality 2.
Stratigraphic range. -Latest Palaeocene to Lower Oligocene (Harland 1979; Costa & Downie 1979b: To Middle Oligocene).
Apectodinium cf. A . quinquelatum (W. & D.) Costa & Downie P1. 2, Figs. 3-5
A distinctive group of specimens comes close to A . quinquelatum sensu Harland (1979, p. 67), except for their more prominent apical horn, which may place them somewhere between that species and A . sumissum (Harland) L. & W. Resemblance to cysts identified as Wetzeliella (Apectodinium) hyperacantha in Costa & Downie (1976, PI. 92, Fig. 6), cf. that species by Harland (1979, P1. 2, Fig. 19), is also exhibited in this group.
Spitsbergen record. - Sarkofagen and Gilson- ryggen Fms., localities 2, 3.
Stratigraphic range. - A . quinquelatum: From latest Palaeocene to Lower Eocene, NPSNP13 equivalent according to Harland (1979), extend- ing into Upper Eocene according to Costa & Downie (1979b).
Cribroperidinium Neale & Sarjeant
Cribroperidinium giuseppei (Morgenroth) Helenes P1. 2, Figs. 7-8 Spitsbergen record. - Forlandsundet, locality 5 .
Stratigraphic range - Eocene, top in NP19 equi- valent strata (Williams & Bujak 1985); London

110 S . B. Manum & T. Throndsen
Clay through Barton Beds, southern England (Bujak et al. 1980).
Deflandrea Eisenack Nomenclatural note. - Of the species recorded below, D. dartmooria and D. striata belong to a group which formerly has been separated from Deflandrea and placed under Ceratiopsis Vozz- hennikova, likewise in Lentin & Williams (1985). However, Lindgren (1984, p. 154) pointed out that the latter name is a junior homonym of a genus of fungi, Ceratiopsis De Wilde. The re- attribution here to Deflandrea therefore rep- resents a deviation from Lentin & Williams (1985).
Deflandrea dartmooria Cookson & Eisenack PI. 3, Figs. 1-2 Spitsbergen record. - Basilika Fm., locality 4.
Stratigraphic range. - Upper Palaeocene to Lower Eocene, NP4-NP13 equivalent (Williams & Bujak 1985); North Sea: Upper Palaeocene- basal Lower Eocene (Ioakim 1979).
Deflandrea heterophlycta Deflandre & Cookson sensu Gocht P1. 4, Figs. 1, 3-4
01960 Depandrea phosphoririca Eisenack: Manum, p. 18. Figs. 4-6, text-fig. 1. 01969 Deflandrea cf. hererophlycra Deflandre & Cookson: Gocht, p. 1 1 , text-fig. 4. 01980 Deflandrea cf. D . hererophlycra Deflandre & Cookson sensu Gocht: Bujak, in Bujak et al . , p. 50. P1. 15, Fig. 4.
This form, which is characterized by a denticulate periphragm in a more or less clearly penitabular pattern and a warty endophragm, matches closely the Oligocene form referred to as D. cf. hetero- phlycta by Gocht (1969). Subsequently, Bujak (in Bujak et al. 1980, p. 50) recorded the same form in the Barton Beds of southern England. (Bujak listed the Spitsbergen form illustrated by Manum (1960) under D . spinulosa synonymy, but according to our present identification it should be assigned to D. heterophlycta sensu Gocht.)
Spitsbergen record. - Forlandsundet, locality 5 .
Stratigraphic range. -Barton Beds, southern Eng- land (Bujak et al. 1980; Middle Eocene, NP16- 17 equivalent acc. to Williams & Bujak 1985);
Middle Oligocene, Germany (Gocht 1969). Wil- liams & Bujak (1985) give the range of D. het- erophlycta as Middle Palaeocene through Oli- gocene, NP16-NP25 equivalent.
Deflandrea oebisfeldensis Alberti PI. 3 , Figs. 3 4
Spitsbergen record. - Gilsonryggen Fm., locality 2.
Stratigraphic range. -Upper Palaeocene to Lower Eocene, NP7-NP12 equivalent (Williams & Bujak 1985); North Sea and Labrador: Palaeo- cene to Lower Eocene (Ioakim 1979).
Deflandrea striata Drugg P1. 3 , Figs. 5-7
Spitsbergen record. - Basilika Fm., locality 4.
Stratigraphic range. - Offshore eastern Canada: Maastrichtian - Lower Palaeocene (Williams & Bujak 1977); Denmark: Danian - Lower Seland- ian (Heilmann-Clausen 1985); Switzerland: Danian - Lowermost Thanetian (Jan du ChCne 1977); North Sea and Labrador: Lower Palaeo- cene (Danian) (Ioakim 1979).
Diphyes Cookson
Diphyes colligerum (Deflandre & Cookson) Coo kson P1. 2, Fig. 9 Spitsbergen record. - Gilsonryggen Fm., locality 1.
Stratigraphic range. - Eocene (Williams & Bujak 1985); North Sea and Labrador: Lower Eocene (Ioakim 1979); southern England: London Clay through Barton Beds (Bujak et al. 1980); Denmark: Danian - Lower Ypresian (Heilmann- Clausen 1985).
Fibrocysta Stover & Evitt
Fibrocysta axialis (Eisenack) Stover & Evitt P1. 2, Fig. 10
Spitsbergen record. - Gilsonryggen Fm., locality 1.

Age of Tertiary formations 111
Stratigraphic range. - Lower Eocene to Lower Stratigraphic range. - Late Middle Eocene to Oligocene (Williams & Bujak 1985); North Sea: Lower Oligocene, NP16/17-NP23 equivalent L. Eocene-L. Oligocene (Ioakim 1979); southern (Williams & Bujak 1985); Barton Beds, S. Eng- England: London Clay through Barton Beds land (Bujak et al. 1980). (Bujak et al. 1980).
Lentinia wetzelii (Morgenroth) Bujak Glaphyrocysta Stover & Evitt P1. 5 , Figs. 4-6
Spitsbergen record. - Gilsonryggen Fm., locality Glaphyrocysta exuberans (Defl. & Cooks.) 1. Stover & Evitt P1. 6, Figs. 2-3 Stratigraphic range. - London Clay and Brackles- This species is a common and conspicuous ham Beds, S. England (Bujak et al. 1980; Lower element in the assemblage from the Gilsonryggen Eocene-earlY Middle Eocene act. to Williams Formation. Bujak 1985).
Spitsbergen record. - Gilsonryggen Fm., locality 2.
Stratigraphic range. - From the Upper Palaeocene (Pyrenees, Car0 1973; offshore eastern Canada, Williams & Bujak 1977; Labrador, Ioakim 1979) to Upper Eocene (S. England, Eaton 1976; off-shore eastern Canada, Williams & Bujak 1977).
Lentinia sp./spinidinium sp P1. 5 , Figs. 7-9 Some specimens with wide archeoPYles differ from L. serrata in their share and development of horns (Figs- 7,8); others appear to be more related to SPinidinium (Fig. 9).
Spitsbergen record. - Gilsonryggen Fm., locality 1.
Kisselouia Vozzhennikova
Kisselouia crassoramosa (Williams & Downie) Damassa P1. 4, Fig. 8 The figured specimen does not exhibit the general shape well because of folding. However, the pro- cesses with trabeculae characteristic of this species are well shown.
Spitsbergen record. - Forlandsundet, locality 5.
Stratigraphic range. - Lower Eocene (London Clay, S. England, Williams & Downie 1966; N. Germany, W. meckelfeldensis zone, Costa & Downie 1976; Labrador, Ioakim 1979); Upper Eocene (offshore eastern Canada, Williams & Bujak 1977).
Lentinia Bujak
Lentinia serrata Bujak P1. 5 , Figs. 1-3 Spitsbergen record. - Gilsonryggen Fm., locality 1; Forlandsundet, locality 5.
Palaeoperidinium Deflandre
Palaeoperidinium pyrophorum (Ehrenberg) Sarjeant P1. 6 , Figs. 5-6 The specimens appear to be smaller than usual for this species.
Spitsbergen record. - Basilika Fm., locality 4.
Stratigraphic range. - Campanian to Upper Palaeocene, top in NP8 equivalent strata (Wil- liams & Bujak 1985).
? Palaeoperidinium sp . P1. 6 , Figs. 7-9 Identification of this Palaeoperidinium-like form is uncertain because it lacks distinct ornament- ation, and the archeopyle has not been observed.
Spitsbergen record. - Gilsonryggen Fm., locality 1.
Phthanoperidinium Drugg & Loeblich

112 S . B . Manum & T. Thronaken
Phthanoperidinium echinatum Eaton P1. 4, Figs. 5-7 Epicyst of illustrated specimen distorted. However, the characteristic parasutural double line of pin-like projections is shown.
Spitsbergen record. - Forlandsundet, locality 5.
Stratigraphic range. - Lower Eocene to Lower Oligocene (Williams & Bujak 1985); Brackles- ham Beds to Barton Beds 3, S. England (Bujak et al. 1980; NP14NP17 equivalent acc. to Wil- liams & Bujak 1985); Norwegian Sea and North Sea: Middle and Upper Eocene (Manum 1976 and Ioakim 1979 respectively).
Selenopemphix Benedek
Selenopemphix nephroides Benedek P1. 5 , Fig. 10 Spitsbergen record. - Forlandsundet, locality 5.
Stratigraphic range. - Late Middle Eocene to Upper Miocene, NP16NN11 equivalent (Wil- liams & Bujak 1985); Germany: Middle to Upper Oligocene (Benedek 1972); S. England: Barton Beds (Bujak et al. 1980, NP16/17 equivalent acc. to Williams & Bujak 1985).
Svalbardella Manum
Svalbardella cooksoniae Manum P1. 5 , Figs. 11-15 The figured specimens show some of the varia- bility exhibited by this species; they are all from the type material. For comparison one specimen from Upper Eocene sediments of the Norwegian Sea (DSDP Leg 38) is shown in PI. 4, Fig. 1.
Spitsbergen record. - Forlandsundet, locality 5.
Stratigraphic range. - North Sea: Middle Eocene to Lower Oligocene (Ioakim 1979); Norwegian Sea: Upper Eocene (Johnsen 1981; Elde 1985).
Thalassiphora Eisenack & Gocht
Thalassiphora pelagica (Eisenack) Eisenack & Gocht PI. 6, Fig. 4
Spitsbergen record. - Forlandsundet, locality 5.
Stratigraphic range. - Maastrichtian to Lower Oligocene (Williams & Bujak 1985); Norwegian Sea and Rockall Plateau: Eocene (Manum 1976; Costa & Downie 1979a; North Sea: Lower Eocene to Lower Miocene (Ioakim 1979).
Turbiosphaera Archangelsky
Turbiosphaera cf. T. filosa Archangelsky P1. 6 , Fig. 1 Preservation does not permit a definitive identification.
Spitsbergen record. - Gilsonryggen Fm., locality 2.
Stratigraphic range. -(For T. filosa) Maastrichtian to Middle Eocene (Williams & Bujak 1985).
Discussion The recorded dinoflagellates are listed in Fig. 4, together with their stratigraphic ranges, forming a basis for discussion of the age of the individual formations.
Basilika Formation: LowerlUpper Palaeocene transition
Only Deflandrea dartmooria, D. striata and Palaeoperidinium pyrophorum have been ident- ified from the Basilika Formation. Based on their known ranges we can only propose an age near the Lower to Upper Palaeocene transition. Thus, there is a fair amount of agreement with Rosen- krantz (1942) and Vonderbank (1970) who indi- cated a Danian age, and also with the (Middle?-) Upper Palaeocene age based on molluscs by Livshits (1974).
Sarkofagen Formation: Late Upper Palaeocene
In the upper part of this formation we have observed Apectodinium. For discussion of the age, see below under Gilsonryggen Formation.
Gilsonryggen Formation: Late Upper Palaeocene
A varied assemblage of Apectodinium species was recovered from apparent ash layers in this for-
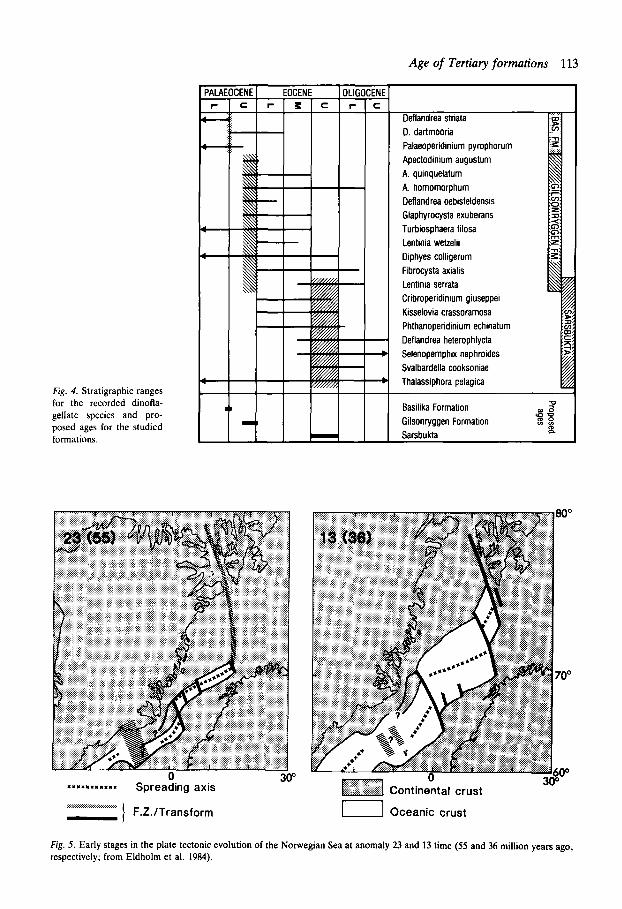
Age of Tertiary formations 113
Fig. 4. Stratigraphic ranges for the recorded dinofla- gellate species and pro- posed ages for the studied formations.
Deflandrea striata D dartmooria Palaeoperidinium pyrophorum Apectodinium augustum A quinquelatum A homomorphum Deflandrea oebisfeldensis Glaphyrocysta exuberans Turbiosphaera filosa Lentinia wetzelii Diphyes colligerum Fibrocysta axialis Lentinia serrata Cribroperidinium giuseppei Kisselovia cnssoramosa Phthanoperidinium echinatum Deflandrea heterophlycta Selenopemphix nephroides Svalbardella cooksoniae Thalassiphora pslagica
9 go"
Gilsonryggen Formation r n r n 8
Basilika Formation
Sarsbukta
(Do
I I Oceanic crust
Fig. 5. Early stages in the plate tectonic evolution of the Norwegian Sea at anomaly 23 and 13 time (55 and 36 million years ago, respectively; from Eldholrn et al. 1984).

114 S. B. Manum & T. Throndren
mation. Partkular attention is drawn to Apecto- dinium augustum which has a narrow strati- graphic range in Northwest Europe and the North Sea, restricted to the later part of the Upper Palaeocene (equivalent to NP9-10 strata). In addition, the assemblage presents a wide range of morphological variations corresponding to the ’ Wetteliella (Apectodinium) homomorphum plexus’ described by Harland (1979). This com- plex of morphotypes is characteristic of the Apectodinium hyperacanthum zone (= W. hyper- acantha zone) of Costa & Downie (1976, text- fig. 5 ; 1979b). In NW Europe it is equivalent to NP9 and earliest NP10. The first appearance of Apectodinium in NW Europe is in strata equiva- lent to NP9 (Harland 1979). On the basis of this Apectodinium association we assign the Gilson- ryggen Formation and also the upper part of the Sarkofagen Formation to late Upper Palaeo- cene.
Forlandsundet (Sarsbukta): Upper Eocene
The identifications of Cribroperidinium giuseppei and Kisselooia crassoramosa indicate that the Sarsbukta sediments are no younger than the Upper Eocene. The possibility of a somewhat greater age cannot be excluded, since most of the recorded species range down into Middle and Lower Eocene. However, we base the assignment to Upper Eocene on the presence of Sualbardella cooksoniae, which does not appear to range down into Middle Eocene. It is worth noting that all the recorded species occur in the Barton Beds (Upper Eocene) of southern England (Bujak et al. 1980). Our conclusion is at variance with that of Feyling- Hanssen & Ulleberg (1984: M.-U. Oligocene) but supports the more loosely founded Upper Eocene age proposed for the Sarsbukta sequence by Livshits (1974). Consequently, Livshits’ proposed correlation with the Gilsonryggen Formation of the central basin cannot be supported.
Geological Synthesis The Tertiary basin in Spitsbergen is situated close to the margin of the Barents Sea plate, and its development is clearly controlled by the plate tectonic events which led to the formation of the Norwegian-Greenland Sea and the connection between the North Atlantic and the Arctic Oceans
(Fig. 5). The opening of the Norwegian Sea started at anomaly 24-25 time in the latest Palaeocene to earliest Eocene according to Palmer (1983). In Fig. 6 we have presented a time-stratigraphic correlation between the Ter- tiary deposits in the central basin, Forlandsundet (Sarsbukta), Renardodden, and plate tectonic events in the Norwegian-Greenland Sea.
The central basin
The sedimentological studies by Steel et al. (1981) have demonstrated three main stages of depo- sitional development in the central basin which have been related to Palaeogene tectonic events in the Norwegian-Greenland Sea. The tectonic control of the depositional pattern was discussed in more detail by Steel et al. (1986).
The first stage, according to Steel et al. (1981), is represented by the Firkanten and Basilika For- mations and presents a period of transgression with sediment supply from the east and north- east. This development took place in the Palaeo- cene. The subsidence probably resulted from an early transtensional regime between the Green- land and the Barents Sea blocks.
The second stage, represented by the Sarko- fagen and the lower part of the Gilsonryggen Formations, shows a period of regression. During the ultimate phase of this development (repre- sented by the Hollendardalen Formation), sedi- ment influx was from the west. This incipient sediment supply from the west is taken by Steel et al. (op. cit.) as evidence of initiation of western uplift and onset of a transpressional regime along the sheared margin. This shift in basin geometry is dated by our dinoflagellates from the Gilson- ryggen Formation to late Upper Palaeocene, thus associating it in time with starting of sea floor spreading in the Norwegian Sea at anomaly 24- 25 time.
The third stage of sedimentary development according to Steel et al. (1981) is represented by the upper part of the Gilsonryggen Formation and the Battfjellet and Aspelintoppen Forma- tions, and this presents a second period of regression with sediment supply from the west and north. According to our age assignment for the Gilsonryggen Formation, this development took place from the earliest Eocene onwards, when a sheared margin and a transpressional regime still existed between the Greenland and the Barents Sea blocks. During the latest part of

STEE
L E
l AL
. 19
81
MU
OR
AND
NAGY
191
2
THE
CENT
RAL
BASI
N AC
CEPT
EO A
GE
SEDI
MEN
T SU
PPLY
SE
DIM
ENT
DETE
RMIN
ATIO
NS
Slee
l el a
l. 19
81
ACCU
MUU
TIO
N PA
TER
N
Man
urn
and
Thm
ndrs
n 19
78
*___h
ASPE
LINT
OPP
EN
FM
UlT
FJE
UE
T FM
GIL
SONR
VGG
EN
FM
..
TR
ANST
ENSI
ON
1500
m e
rode
d (
MAN
UM
AN
0 TH
RONO
SEN
1978
1
, -sw
NN
i
(MIO
OLE
? -
) UP
PER
SIRN
OFA
GEN
PA
UEO
CENE
UT
E U
PPER
PU
EOC
ENE
FlR
lWrr
m
FM
1 LOW
ER-U
PPER
I
0 0 0 0
SUPP
LY 1 FROM W
. NW
. N
FIRS
T SI
GN
OF
CHAN
GE IN
AR
EA O
F SU
PPLY
SUPP
LY IT FROM E.
ME
0 0 0 o i-l
EAST
WAR
D M
IGRA
TIO
N OF
ARE
A OF
PMM
OC
ENE
PILI
SDC
ENE
(OAN
IAN)
IIMEN
T riom
NORW
EGIA
N. G
REEN
LAND
SE4
EVE
NTS
iEA.
FLO
OR S
PREM
ING
M
agna
lic c
amla
lion
au. I
D Pa
lmer
198
3
PRE
3 A
NO
MM
V 13
STAR
TING
OF
SEA.
FLO
OR
ANOM
ALY
24 - 2
5
smm
oiwc
IPITS
BERG
EN F
RLCT
URE
,?OM
S
lml e
l al.
1981
TRAI
(SPR
ESSI
ON
AND
DEVE
LOPM
ENT
OF M
IGR
AM
IG R
IFI
WN
S W
NG
TH
E W
EST
CO
W
OF S
PWSB
LRG
EN L
Fig. 6
. Pro
pose
d sy
nthe
sis o
f Sp
itsbe
rgen
Ter
tiary
bio
stra
tigra
phy,
bas
inal
evo
lutio
n, a
nd m
ajor
pla
te t
ecto
nic
even
ts in
the
Nor
weg
ian-
Gre
enla
nd S
ea.

116 S. B. Manum & T. Throndsen
this stage there was an increasingly rapid uplift and high sediment influx as evidenced by the great thickness of sandstones of the Aspelintoppen For- mation. Vitrinite reflectance studies (Manum & Throndsen 1978; Throndsen 1982) indicate that another 1500 metres of sediments were deposited on top of the existing formations. These missing sediments were conceivably also deposited during this stage of high accretion. There is no reliable indication so far that they extended into the Oligocene.
N~ttvedt (1985, p. 44) and Steel et al. (1986, p. 342) proposed a correlation between the initia- tion of the regressive cycle observed at the base of the Sarkofagen Formation and the global eustatic drop in sea level at the Early-Late Palaeocene transition proposed by Vail et al. (1977). This would seem to agree well with the dinoflagellate based age proposed here for the Basilika For- mation (Lower-Upper Palaeocene transition). However, in our opinion this regressive event may equally well be controlled by the local tectonic setting.
The numerous yellowish weathering clays in the Gilsonryggen Formation, some of which have yielded useful dinoflagellate assemblages, appear to be ash layers. Apparently there was con- siderable volcanic activity in or near the Spits- bergen area at the time of the Palaeocene-Eocene transition, at about the same time as the ‘ash- markers’ occur in the North Sea.
Forlandsundet
The Tertiary sequence of Forlandsundet is pre- served in a narrow, elongated graben structure within the West Spitsbergen fold belt (Fig. 1) which is isolated from the central Tertiary basin. The graben cuts into pre-Devonian strata along steeply dipping faults on either side of For- landsundet (Atkinson 1962; Livshits 1974). It displays a sequence of alluvial fan, fan delta, nearshore marine, and submarine fan deposits with an estimated stratigraphic thickness of more than 5 kilometres (Rye-Larsen 1982).
Vitrinite reflectance measurements on Tertiary samples from either side of Forlandsundet, from Sarsbukta and Reinhardpynten respectively, show highly contrasting values. The measure- ments from Sarsbukta give values of around Rm = 0.3, suggesting a modest overburden in the order of maximum 2 kilometres (depending on the geo-
thermal regime in the area). The measurements from Reinhardpynten, on the other hand, give very much higher values, Rm = 4.0, indicating the anthracite stage of organic maturation. This is an abnormally high maturation level for the Spitsbergen Tertiary and indicates either a very thick overburden (6-8km), or a lesser over- burden in combination with a higher than normal heat flow (possibly from igneous activity). The latter explanation appears more likely in view of the modest maturation level recorded at Sarsbukta only a few kilometres away.
Steel & Worsley (1984) and subsequently Steel et al. (1985) proposed that the Forlandsundet graben originated in the Late Palaeocene-Eocene and developed as a southerly migrating trough within the mobile belt under a transpressional strike-slip regime. Our identification of an Upper Eocene dinoflagellate assemblage in sediments from Sarsbukta supports this timing of the event.
The Middle to Upper Oligocene age indicated by Feyling-Hanssen & Ulleberg (1984), which is not supported by our study, would imply that the Forlandsundet graben and its sediments were formed under quite a different tectonic regime, namely when rifting had already started between the Greenland and the Barents Sea blocks (post anomaly 13 time).
Renardodden
The Tertiary deposits at Renardodden consist of conglomerates (at base), coal-bearing shales and sandstones resting unconformably on rocks of the Hecla Hoek basement complex (Precambrian). Total thickness of the sequence is estimated at around 300 metres by Atkinson (1963) and Thie- dig et al. (1980), whereas Vonderbank (1970) estimated 130 metres.
As discussed earlier, age assignment and cor- relation with formations of the central basin are equivocal. We favour Head’s (1984) indication of an Upper Eocene to Lower Oligocene age. It would follow from this that this sub-basin devel- oped during the tectonic stage of shear movement and transpressional regime between the Green- land and the Barents Sea blocks (Fig. 5). This is in agreement with the interpretation offered by Kellogg (1975) and Harland et al. (1976) that the sequence was deposited in a local basin faulted into the structures of the West Spitsbergen oro- genic belt at about the same time as the formation of the Forlandsundet basin.

Age of Tertiary formations 117
Dartmouth, for helpful discussions; he is also grateful to Dr. Williams and the Atlantic Geoscience Center for the hospitality extended to him on his visit in 1985. A travel grant from the Norwegian Research Council for Science and the Humanities is also gratefully acknowledged. Throndsen acknowledges financial support towards field work in Spitsbergen from the Norwegian Research Council for Science and Technology (the Svalbard-projgct, University of Oslo).
Conclusion A review of biostratigraphic evidence for the age of the Tertiary formations on Spitsbergen reveals that remarkably few useful fossil assemblages have been found. Proposed ages are generally vague and often equivocal.
An early Palaeocene age (Danian) for the Fir- kanten Formation and part of the Basilika For- mation appears to be well established and is sup- ported by the present study. However, a more precise age for the initiation of deposition of the Firkanten Formation following the Upper Cre- taceous hiatus is still lacking.
The lower part of the Gilsonryggen Formation is dated herein to late Upper Palaeocene; conse- quently, this also becomes the age of the change in basin geometry and sediment source area observed in the formation. The timing of this event is thus associated with initiation of sea floor spreading in the Norwegian Sea at anomaly 24- 25 time.
The younger formations of the central basin (Battfjellet and Aspelintoppen Formations) have so far yielded very little of biostratigraphic value. An Eocene age appears most acceptable from available evidence from palaeontology as well as general basinal considerations; no clear evidence exists for an Oligocene or younger age.
The ages of the Tertiary sub-basins outside the central basin and their correlation with formations of the central basin as proposed by various authors are equivocal. An Upper Eocene age is proposed herein for the sequence at Sarsbukta, whereas no new evidence is available for Prins Karls Forland itself. Head (1984) indicated an Upper Eocene to Lower Oligocene age for the Renardodden sub- basin based on dinoflagellates. These ages fit a model which places the formation of these sub- basins before anomaly 13 time, that is, under a transpressional regime along the shear-zone before the two plates started to separate.
Acknowledgements. - Wc wish to thank the following for pro- viding samples: Dr. R. w. Feyling-Hansscn, Universitetct i Aarhus; Dr. D. E. Russell, Museum National d’Histoirc Natu- relle, Paris; Professor J . Thicdc, Gcologisch-Palaontologisches lnstitut und Museum der Universitat Kiel, and his students’ field group to Spitsbergen 1983, financially supported by DEMINEX (Norge) A.S.
The skilful processing of samples by MIS. A. Dugan, Geology Department, Oslo, is gratefully acknowledged.
Manum is grateful to Dr. J . P. Bujak, Bujak Research Ltd., Calgary, and Dr. G. L. Williams, Atlantic Geoscience Center,
References Atkinson, D . J . 1962: Tectonic control and sedimentation and
the interpretation of sediment alternation in the Tertiary of Prince Charles Foreland, Spitsbergen. Bull. Geol. Soc. Am. 73, 343-364.
Atkinson, D. J . 1963: Tertiary rocks of Spitsbergen. Bull. Am. Ass. Pefr. Geol. 47, 302-323.
Benedek, P. N. 1972: Phytoplanktonen aus dem Mittel- und Oberoligozan von Tonisberg (Niederrheingebiet). Palae- ontographica I37 B , 1-71, 16 PIS.
Bujak, J. P., Downie, C., Eaton, G. L. & Williams, G . L. 1980: Dinoflagellate cysts and acritarchs from the Eocene of southern England. Palaeont. Ass. London, Spec. Pap. Palaeont. 24. 100 pp., 22 PIS.
Caro, Y. 1973: Contribution B la connaiscance des dinoflagellks du Palakoche-Eockne infkrieurs des Pyrknkes Espagnoles. Reuista Espan. Micropal. 5 , 329-312, 5 pls.
Costa, L. & Downie, C. 1976: The distribution of the dino- flagellate Wetzeliella in the Palaeogene of north-western Europe. Palaeontology 19, 519-614. 1 pl.
Costa, L. & Downie. C. 1979a: Cenozoic dinocyst stratigraphy of Sites 403 to 406 (Rockall Plateau), IPOD, Leg 48. Init. Rep. DSDP 48. 513-529.
Costa, L. & Downie, C. 1979b: The Wefzeliellaceae; Palaeogene dinoflagellates. IV Intern. Palynol. Conf., Lucknow (1976
Dypvik, H. & Nagy J . 1979: Early Tertiary bentonites from Svalbard. Geol. Mag. 116. 457-468.
Eaton. G. L. 1976: Dinoflagellate cysts from the Bracklesham Beds (Eocene) of the Isle of Wight, southern England. Bull. Brit. MuFeum (Nat. Hist.) Geol. 269 227-332, 21 pls.
Elde, T. 1985: Dinoflagellafloraen i mure eocen i DSDP Site 338, Vmringplatdet. - Unpubl. thesis (Geol.), Univ. Oslo. 142 PP.
Eldholm, E.. Myhre, A. M., Sundvor, E. & Faleide, J. I. 1984: Cenozoic evolution of the margin off Norway and Svalbard. Pp. 3-18 in Spencer, A. M. et al. (eds.): Pefroleum geology of the North European margin. Graham & Trotman Ltd., London.
Feyling-Hanssen, R. W. & Ulleberg, K. 1984: A Tertiary- Quaternary section at Sarsbukta, Spitsbergen, Svalbard, and its foraminifera. Polar Res. 2, 77-106, 4 pls.
Gocht, H. 1969: Formengemeinschaften alttertiaren Mikro- planktons aus Bohrproben des Erdolfeldes Meckclfeld bei Hamburg. Palaeontographica B126. 1-100. 11 pls.
Harland, R. 1979: The Wetzeliella (Apectodinium) homo- morpha plexus from the Palaeogene/earliest Eocene of North West Europe. IV Intern. Palyn. Conf., Lucknow (1976-77) 2 , 59-70, 2 pls.
Harland, W. B.. Pickton, C. A. G., Wright, N. J. R., Croxton, C. A, , Smith, D. G. , Cutbill, J. L. & Henderson, W. G.
77) 2 , 3643.

118 S. B. Manum & T. Throndsen
1976: Some bal-bearing strata in Svalbard. Norsk Polarinst. Skr. 164. 89 pp.
Head, M. 1984: A palynological investigation of Tertiary strata at Renardodden, W. Spitsbergen. Abstr. 6th Intern. Palyn. Conf. Calgary 1984. 61 pp.
Heer, 0. 1868: Florafossilis arctica. 1. Zurich. 192 pp., 50 pls. Heer, 0. 1870 Flora fossilis arctica. I1 ( 3 ) . In K. Suenska
Vetensk. Akad. Handl. 8 (7). 98 pp.. 16 pls. Heer, 0. 1876: Flora fossilis arcrica. IV (1). In K . Svenska
Vetensk. Akad. Handl. 14 (5). 141 pp.. 32 pls. Heilmann-Clausen, C. 1985: Dinoflagellate stratigraphy of the
uppermost Danian to Ypresian in the Viborg 1 borehole, central Jylland, Denmark. Danm. Geol. Llnders. Ser. A (7). 69 pp., 15 pls.
Hjelle, A. & Lauritzen, 0. 1982: Geological map of Svalbard 1:5W OOO. Sheet 3G, Spitsbergen northern part. Norsk Polar- insr. Skr. 154C. 15 pp.. 1 map.
Jan du Chine, R. E. 1977: Palynostratigraphie (Maastrichtien- Eocene infkrieur) des flysch du Schlieren (Canton D’Obwald, Suisse Centrale). Reu. Micropal. 20, 147-156.
Ioakim, Ch. 1979: Etude comparative des dinojlagell.?s du Ter- tiaire inferieur de la Mer du Labrador et de la Mer du Nord. These Geol., Univ. P. & M. Curie, Paris. 204 pp., 23 pls.
Johnsen, K. 1981: Palynologisk undersgkelse i tertirerpd Island- F~royryggen (Site 336) DSDP materialer. Unpubl. thesis (Geol.), Univ. Oslo. 126 pp.
Kellogg, H. E. 1975: Tertiary stratigraphy and tectonism in Svalbard and continental drift. Bull. Am. Assoc. Petrol. Geol. 59. 465-385.
Lentin. J. K. & Williams, G. L. 1985: Fossil dinoflagellates: Index to genera and species. 1985 edition. Can. Techn. Rep. Hydrogr. Ocean Sci. 60. 451 pp.
Lindgren. S . 1984: Acid resistant peridinoid dinoflagellatesfrom the Maastrichtian of Trelleborg. southern Sweden. Stockholm Contrib. Geol. 39. 145-201.
Livshits, Ju. Ja. 1974: Palaeogene deposits and the platform structure of Svalbard. Norsk Polarinst. Skr. 159. 50 pp.
Major, H. & Nagy, J. 1972: Geology of the Adventdalen map area. Norsk Polarinst. Skr. 138. 58 pp., 1 map.
Manum. S . B. 1960: Some dinoflagellates and hystricho- sphaerids from the lower Tertiary of Spitsbergen. Nya. Mag. Bot. 8 , 17-26. 1 pl.
Manum. S. B. 1962: Studies in the Tertiary flora of Spitsbergen, with notes on Tertiary floras of Ellesmere Island, Greenland, and Iceland. A palynological investigation. Norsk Polarinst. Skr. 125. 127 pp., 20 pls.
Manum. S . B. 1976: Dinocysts in Tertiary Norwegian-Green- land Sea sediments (Deep Sea Drilling Project Leg 38). with observations on palynomorphs and palynodebris in relation to environment. Pp. 897-919 in Talwani, M., Udintsev, G. et al.: Initial Reports of the Deep Sea Drilling Project, 38. U.S. Government Printing Office, Washington D.C.
Manum, S. B. & Throndsen, T. 1978: Rank of coal and dis- persed organic matter and its geological bearing in the Spitsbergen Tertiary. Norsk Polarinst. Arbok 1977,159-177.
Nathorst. A . G. 1910: Beitrage zur Geologie der Bareninsel,
Spitzbergens und des Konig-Karl-Landes. Bull. Geol. lnst. Uppsala 10, 261-416.
N~ttvedt , A. 1985: Askeladden delta sequence (Palaeocene) on Spitsbergen - sedimentation and controls on delta formation. Polar Research 3 , 21-48.
Palmer, A. R. 1983: The decade of North American geology - 1983 geologic time scale. Geology 11, 502-504.
Ravn, J. P. J. 1922: On the Mollusca of the Tertiary of Spits- bergen. Result. Norske Spitsbergeneksp. 1 (2). 28 pp.. 2 pls.
Rosenkrantz, A. 1942: Slregten Thyasira’s geologiske Optrae- den. Meddel. Dansk Geol. For. 10, 277-278.
Rye-Larsen, M. 1982: Forlandsundet Graben (Paleogen) - Sual- bard’s vestmargin - sedimentasjon og tektonbk utuikling au et basseng ued en transform plategrense. Cand. real. thesis (Geol.), Univ. Bergen. 380 pp.
Steel, R., Dalland, A, , Kalgraff, K. & Larsen, V. 1981: The central Tertiary basin of Spitsbergen: Sedimentary develop- ment of a sheared-margin basin. Can. Soc. Petrol. Geol. Mem. 7, 647-664.
Steel, R. J . & Worsley, D. 1984: Svalbard’s post-Caledonian strata: An atlas of sedimentational patterns and palaeo- geographic evolution. Pp. 109-135 in Spencer, A. M. et al. (eds.): Petroleum geology of the North European margin. Graham & Trotman Ltd., London.
Steel, R., Gjelberg, J., N~ttvedt , A,, Helland-Hansen, W., Kleinspehn, K. & Rye-Larsen, M. 1985: The Tertiary strike- slip basins and orogenic belt of Spitsbergen. Soc. Econ. Paleont. Mineral. Spec. Publ. 37, 339-359.
Thiedig, F., Pickton, C. A. G., Lehmann. U. , Harland, W. B. & Anderson, H. J . 1980: Das Tcrtiar van Renardodden (Ostlich Kapp Lyell, Westspitzbergen, Svalbard). Mittlg. Geol.-Palaeonr. lnst. Uniu. Hamburg 49, 135-146.
Throndsen, T. 1982: Vitrinite reflectance studies of coals and dispersed organic matter in Tertiary deposits in the Advent- dalen area, Svalbard. Polar Research 2, 77-91.
Vail. P. R., Mitchum Jr., R. M. & Thompson, S. 1977: Global cycles of relative changes of sea level. Am. Ass. Petrol. Geol. Mem. 26, 62-97.
Vakulenko, A. S. & Livshits, Ju. Ja. 1971: Palinologiceskaja charakteristika paleogenovych otlozenij Spicbergena (Paly- nological characteristics of Palaeogene deposits of Spitsbergen). NIIGA Uc. Zap. Paleont. Biostrat. 31, 39-50, 4 pls.
Vonderbank, K. 1970: Geologie und Fauna der tertiaren Ablagerungen Zentral-Spitzbergens. Norsk Polarimt. Skr. 153. 119 pp, 21 pls.
Williams, G. L. & Bujak, J. P. 1977: Cenozoic palyno- stratigraphy of offshore eastern Canada. AASP Contrib. Ser. 5A , 14-48, 5 pls, 2 charts.
Williams, G. L. & Bujak, J . P. 1985: Mesozoic and Cenozoic dinoflagellates. Pp. 847-964 in Bolli et al. (eds.): Planktonic stratigraphy. Cambridge UNv. Press.
Williams, G. L. & Downie, C. 1966: Wetzeliella from the London Clay. Pp. 182-198 in Davey et al.: Studies on Meso- zoic and Cainozoic dinoflagellate cysts. Bull. Brit. M u . (Nat. Hist.) Geology Suppl. 3 .

Age of Tertiary formations 119
Plates 1-6 All magnifications are 550x unless otherwise stated. Slides are kept in the Palaeontological Museum of the University of Oslo; the indicated prep. numbers refer to the Museum’s PA series of types. Specimen locations in the slides have been indicated by means of ‘England Finder’ coordinates.

120 S . B . Manum & T. Throndsen
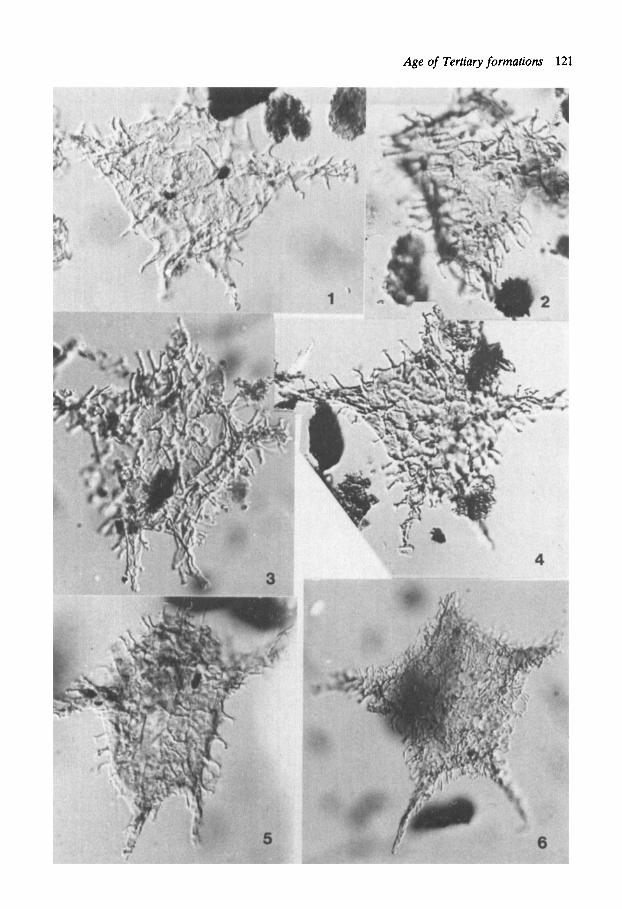
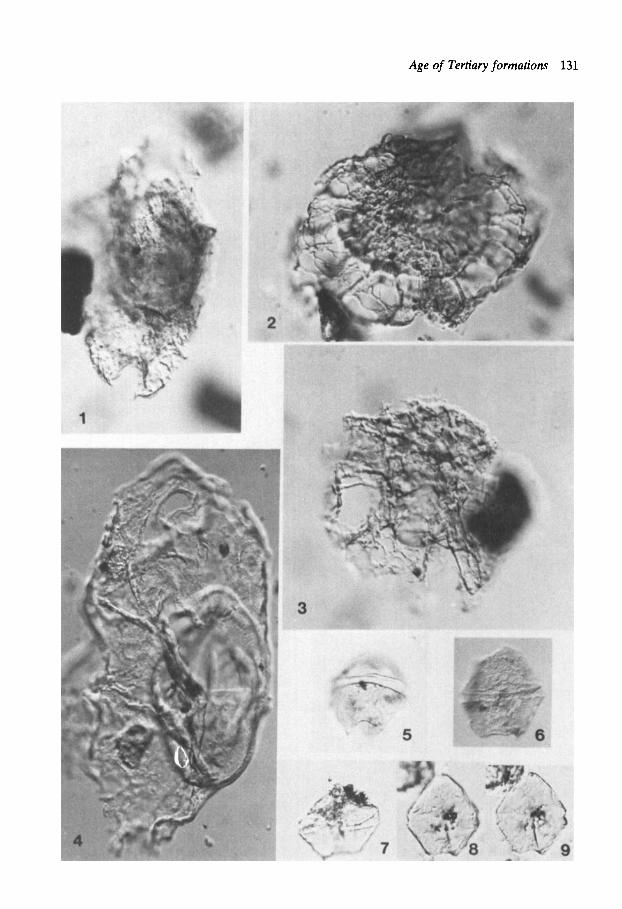
Plate 1 Figs. 1-6. Apectodinium augusrum (Harl.) L. & W., specimens in Figs. 1-5 are from the Gilsonryggen Fm., specimen in Fig. 6 is from the Sarkofagen Fm. Fig. 1: Prep. 4383: T38/1, loc. 1 . Fig. 2: Showing state of preservation with partly distorted and partly broken horns; prep. 4385: R34/4, loc. 1. Fig. 3: Prep. 4384: P53, loc. 1 . Fig. 4: Prep. 4383: L30/1, loc. 1 (SOOX). Fig. 5: Prep. 3890: R49, loc. 2. Fig. 6: Showing variety with finer and more closely spaced ornamentation; prep. 3889: J 4 W / 1-2, loc. 2.
Age of Tertiary formations 121

122 S. B. Manum & T. Thronhen
Plate 2 Fig. I : Apecrodinium augwmm (Harl.) L. & W. Specimen with less developed horns, somewhat approaching A. paniculurum (Costa & Downie) L. & W. Prep. 3889: U2/2-4, Gilsonryggen Fm. loc. 2. Fig. 2: Apecrodinium homomorphum (Defl. & Cooks.) L. & W. Prep. 3890: W44/1, Gilsonryggen Fm. loc. 2. Figs. 3-5: Apectodinium cf. A . quinquelatum (W. & D.) Costa & Downie. Fig. 3: Prep. 3888: M38-39, SarkofagenFm. loc. 3. Figs 4-5: Prep. 3890: VW40/3-1, Gilsonryggen Fm. loc. 2. Fig. 6: Apectodinium aff. A . homomorphum/sumissum/quin- quelarum-complex. Prep. 3890: QR48-49, Gilsonryggen Fm. loc. 2 . Figs. 7-8: Cribroperidinium giuseppei (Morg.) Helenes. Prep. 3882: J38-39/1-2, Sarsbukta loc. 5. Fig. 9: Diphyes colligerum (Defl. &Cooks.) Cooks. Prep. 4383: T40, Gilsonryggen Fm. loc. 1. Fig. 10; Fibrocysta axialis (Eis.) Stover & Evitt. Prep. 4383: D38/4, Gilsonryggen Fm. loc. 1.

Age of Tertiary formations 123

124 S. B. Manum & T. Throndsen
Plate 3 Figs. 1-2: Deflandrea dartmooria Cooks. & Eis. Prep. 3884: 034-35, Basilika Fm. loc. 4. Figs. 3-4: Deflandrea oebkfeldensis Alb. Prep. 3889: MN40/1- 3, Gilsonryggen Fm. loc. 2. Figs. 5-7: Deflandrea striata Drugg. Prep. 3885. Basilika Fm. loc. 4. Figs. S 6 : T45. Fig. 7: S41.

Age of Tertiary formations 125

126 S. B. Manum & T. Throndsen
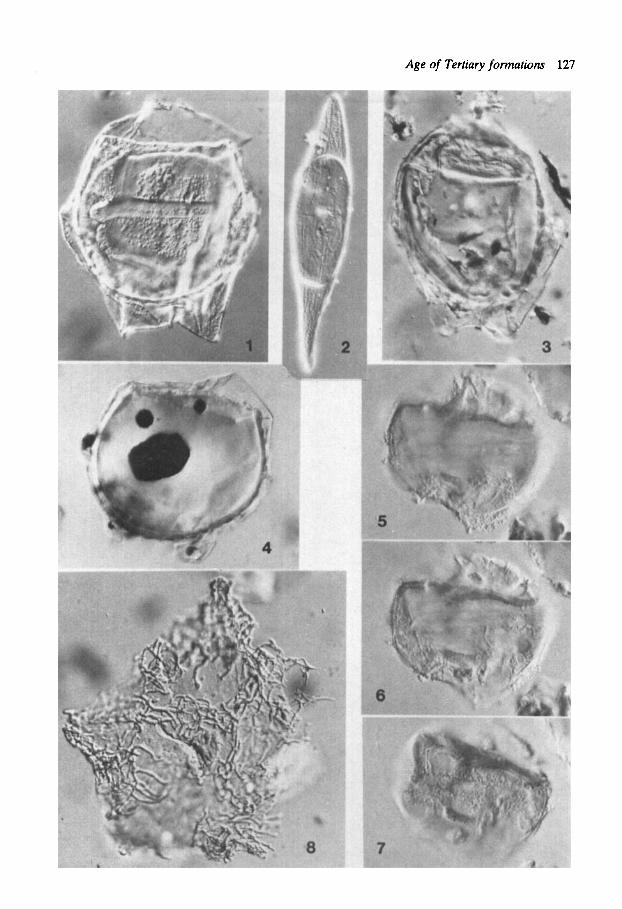
Plate 4 Figs. I , 3-4: Deflandrea hererophlycta Defl. & Cooks. sensu Gocht. Sarsbukta loc. 5 . Fig. 1: Prep. 3883: P42/1. Fig. 3: Prep. 3881: K42/2. Fig. 4: Prep. 3880: R42/1. Fig. 2: Svalbardella cooksoniae Man. Showing specimen from Upper Eocene Norwegian Sea sediments, DSDP 336-23-4. Figs. 5-7: Phthanoperidinium echinarum Eaton. Prep. 3880: T47/3, Sarsbukta loc. 5 (85OX). Fig. 8: KLrsclooia crmsoramosa (Wd. & Downie) Dam. Prep. 3879: W32/2-4, Sarsbukta loc. 5.
Age of Tertiary formations 127

128 S. B. Manum & T. Throndsen
Plate 5 Figs. 1-3: Lenrinia serrafa Bujak. Figs. 1-2: Prep. 4385: 036/ 4, Gilsonryggen Fm. loc. 1 . Fig. 3: Prep. 3878: EF/29, Sarsbukta loe. 5 . Figs. 46: Lenfinia wetzelii (Morg.) Bujak. Prep. 4383: Gil- sonryggen Fm. loc. l; Fig. 4: P37/3. Figs. 5 6 : M44. Figs. 7-9: Lentinia sp./Spinidinium sp. Prep. 4383: Gil- sonryggen Fm. l ~ . l . Fig. 7: K40/3. Fig. 8: K34. Fig. 10: Selenopemphix nephroides Ben. Prep. 3878: K43/1-3, Sarsbukta loc. 5. Figs. 11-15: Soalbardella cooksoniae Man. Sarsbukta loc. 5 . Figs 11-12: Prep. 3880: 034/4. Fig. 13: Prep. 3353. Figs. 14- 15: Prep. 3351.

Age of Tertiary formations 129

130 S. B. Manum & T. Throndsen
Plate 6 Fig. 1: Turbiosphaera cf. T. filosa Arch. Prep. 3890: R41/2, Gilsonryggen Fm. loc. 2. Figs. 2-3: Glaphyrocysta exuberans (Defl. & Cooks.) Stover & Evitt. Prep. 3890: Gilsonryggen Fm. loc. 2. Fig. 2: N43/3. Fig. 3: M4212-4. Fig. 4: Thalassiphorapelagica (Eis.) Eis. & Gocht. Prep. 3878: DE37/1-3, Sarsbukta loc. 5. Figs. 5-6: Palaeoperidinium pyrophorum (Ehrenb.) Sarj. Basi- lika Fm. loc. 4. Fig. 5: Prep. 3886: 042-43/3-4. Fig. 6 Prep.
Figs. 7-9: lPalaeoperidinium sp. Gilsonryggen Fm. loc. 1. Fig. 7: Prep. 4383: 034. Figs. &9: Prep. 4385: 534.
3887: U3637/1-2.
Age of Tertiary formations 131