absence of lytic virus in two species of symbiotic algae within the sea anemone anthopleura...
TRANSCRIPT

Absence of Lytic Virus in Two Species of Symbiotic Algae within the Sea AnemoneAnthopleura xanthogrammica (Coelenterata: Anthozoa)Author(s): Timothy L. O'Brien, R. MacLeod and M. C. MacleanSource: Transactions of the American Microscopical Society, Vol. 103, No. 3 (Jul., 1984), pp.228-232Published by: Wiley on behalf of American Microscopical SocietyStable URL: http://www.jstor.org/stable/3226183 .
Accessed: 30/06/2014 11:30
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley and American Microscopical Society are collaborating with JSTOR to digitize, preserve and extendaccess to Transactions of the American Microscopical Society.
http://www.jstor.org
This content downloaded from 109.241.198.166 on Mon, 30 Jun 2014 11:30:11 AMAll use subject to JSTOR Terms and Conditions

Absence of Lytic Virus in Two Species of Symbiotic Algae within the Sea Anemone Anthopleura xanthogrammica
(Coelenterata: Anthozoa)l
TIMOTHY L. O'BRIEN,2 R. MACLEOD, AND M. C. MACLEAN
School of Basic Medical Sciences and Department of Genetics and Development, University of Illinois, Urbana, Illinois 61801, U.S.A.
Abstract. The production of lytic algal viruses, or virus-like particles, by symbiotic algae isolated from Hydra viridis and Anthopleura xanthogrammica was monitored by sucrose-gradient fractionation of the algal lysates and by transmission electron microscopy. Viral particles were detected readily in algae isolated from H. viridis; however, virus-like
particles were absent from either of the two species of symbiotic algae isolated from A. xanthogrammica. We propose the following hypothesis: the presence of lytic algal viruses in fresh-water algal-invertebrate symbiosis and the apparent absence of such viruses in marine algal-invertebrate symbiosis may represent a fundamental distinction, not previ- ously recognized, between fresh-water and marine algal-invertebrate symbiosis.
Numerous aquatic invertebrates maintain unicellular, eukaryotic algae as intracellular symbionts in what constitutes a mutually beneficial symbiosis (Trench, 1979). Algal-invertebrate symbiosis has been thought to be an inte- grated system consisting solely of the interactions between the alga and the animal host. Recent reports in the literature have established that the assump- tion above is invalid within certain fresh-water algal-invertebrate symbiotic associations. Van Etten and coworkers demonstrated that the fresh-water Hy- dra viridis-algal symbiosis also includes a double-stranded DNA virus (Meints et al., 1981; Van Etten et al., 1981). This virus never has been observed within the intact association-yet upon removal of the algae from hydras, viral par- ticles begin to appear within the algal cytoplasm outside the chloroplast within approximately 2 h. Lysis of the population of isolated algae is complete within approximately 24 h. A similar lytic algal virus is described from the symbiotic algae of fresh-water Paramecium bursaria (Van Etten et al., 1982).
Fresh-water associations represent only a small proportion of the algal-in- vertebrate symbiotic associations known to exist in nature-most associations occur in shallow marine waters. In light of the recent discovery of lytic algal viruses within fresh-water hydras and paramecia, it is of interest to know if marine algal-invertebrate symbiotic associations might also maintain lytic algal viruses.
Viral production in symbiotic algae isolated from Hydra viridis was moni- tored by sucrose-gradient fractionation of algal lysates and by transmission
1 We thank the staff of the Bamfield Marine Station for supplying the sea anemones used in this
study. This investigation was supported by a Biomedical Research Support Grant (NIH RR-7030, 1-5-21748).
2 Address for reprint requests: Department of Physiology and Cell Biology, University of Kansas, Lawrence, Kansas 66045, U.S.A.
TRANS. AM. MICROSC. SOC., 103(3): 228-232. 1984. ? Copyright, 1984, by the American Microscopical Society, Inc.
This content downloaded from 109.241.198.166 on Mon, 30 Jun 2014 11:30:11 AMAll use subject to JSTOR Terms and Conditions

VOL. 103, NO. 3, JULY 1984
electron microscopy (TEM). Also, two distinct species of marine symbiotic algae were isolated from the sea anemone Anthopleura xanthogrammica and examined as above for the presence of virus-like particles. Viral particles were detected readily in algae isolated from H. viridis; however, virus-like particles were absent from either of the two species of symbiotic algae isolated from A. xanthogrammica.
MATERIALS AND METHODS
Animals. Hydra viridis (Florida strain-obtained from Dr. Leonard Mus- catine, University of California at Los Angeles) containing Chlorella-like sym- biotic algae were grown at 23?C in "M" solution (1 mM CaC12, 1 mM NaHCOs, 0.1 mM MgCl2, and 0.1 mM KC1 adjusted to pH 7.6) without Tris buffer (Muscatine & Lenhoff, 1965) under continuous lighting maintained with Syl- vania fluorescent tubes (Gro-lux). Hydras were fed three times each week with freshly hatched brine shrimp.
Anthopleura xanthogrammica (Brandt) with two species of symbiotic al- gae-the dinoflagellate Symbiodinium (=Gymnodinium) microadriaticum Freudenthal (zooxanthellae) and Chlorella-like algae (zoochlorellae) (Musca- tine, 1971; O'Brien, 1978)-were collected from the intertidal zone near Bam- field, British Columbia, Canada, and transported to the University of Illinois. The sea anemones were placed in illuminated aquaria containing artificial seawater (specific gravity, 1.025 at 15?C) maintained at 12?C. Algae within the sea anemones were isolated and examined for the presence of virus-like par- ticles within 24 h following their arrival in Illinois.
Isolation of algae. Hydras were disrupted in a glass-Teflon tissue homoge- nizer containing "M" solution. The algae were removed from the resulting homogenate by centrifugation at 650 x g for 2 min. The pelleted algae were washed three times in "M" solution and then resuspended in "M" solution containing penicillin G (50 U/ml) and streptomycin sulphate (1 ug/ml) to a concentration of approximately 1 x 107 cells/ml as determined with a hema- cytometer. The algal suspension was allowed to stand at 23?C for 24 h.
Tentacles were removed from sea anemones with scissors and the algae teased out of the severed end of each tentacle with the aid of forceps and needles. The algal slurries thus obtained were dispersed gently in a glass-Teflon tissue homogenizer containing artificial seawater. The algae were pelleted at 360 x g for 2 min and washed three times in artificial seawater. The algae were resuspended in artificial seawater containing penicillin G (50 U/ml) and streptomycin sulphate (1 Aug/ml) to a concentration of approximately 2 x 107 cells/ml. The algal suspensions were allowed to stand at 16?C for 24 or 48 h.
Sucrose-gradient fractionation (MacLeod, 1967). Algal suspensions were
centrifuged at 650 x g for 5 min. The supernatant was decanted and centri-
fuged at 25,000 x g for 1 h. The supernatant was discarded and the pellet resuspended in TNE buffer (10 mM Trizma-HCl, 100 mM NaCl, and 1 mM EDTA, pH 7.8). This crude extract was layered on a 5-25% (w/w) linear sucrose rate-density gradient equilibrated with TNE buffer and centrifuged at 33,000 x g for 10 min. Various zones within the gradients were collected
229
This content downloaded from 109.241.198.166 on Mon, 30 Jun 2014 11:30:11 AMAll use subject to JSTOR Terms and Conditions
230 TRANS. AM. MICROSC. SOC.
d^ . a
1 1,
100nm
? . .,
*
4 , ^ .
-1 . * 4 i
1 60n
? "- .' n k ... .
100nm i
2
This content downloaded from 109.241.198.166 on Mon, 30 Jun 2014 11:30:11 AMAll use subject to JSTOR Terms and Conditions

VOL. 103, NO. 3, JULY 1984
through the top of each gradient tube, layered on a 35-55% (w/w) linear sucrose quasi-equilibrium gradient equilibrated with TNE buffer, and centri- fuged at 58,000 x g for 30 min. Various zones within the gradients were col- lected by puncturing the bottom of each gradient tube with a syringe needle. Various fractions thus collected were mixed 1:1 with TNE buffer and centri- fuged at 80,000 x g for 1 h. Resulting pellets were resuspended in 0.5 ml of TNE buffer and examined on Formvar- and carbon-coated copper grids using 0.5% (w/v) ammonium molybdate (pH 7.0) as a negative stain for TEM.
TEM. Algae were pelleted in a clinical centrifuge and fixed in 3% glutar- aldehyde buffered with 0.1 M sodium cacodylate for 1 h at 20?C and for an additional hour at 4?C. Post-fixation was in 1% OsO4 buffered with 0.1 M sodium cacodylate for 1 h at 4?C. Algal pellets then were dehydrated in an acetone series and embedded in Spurr's low-viscosity resin. Ultrathin sections were stained with aqueous uranyl acetate and lead citrate and examined at 80 kV using a Siemens 102 transmission electron microscope.
RESULTS
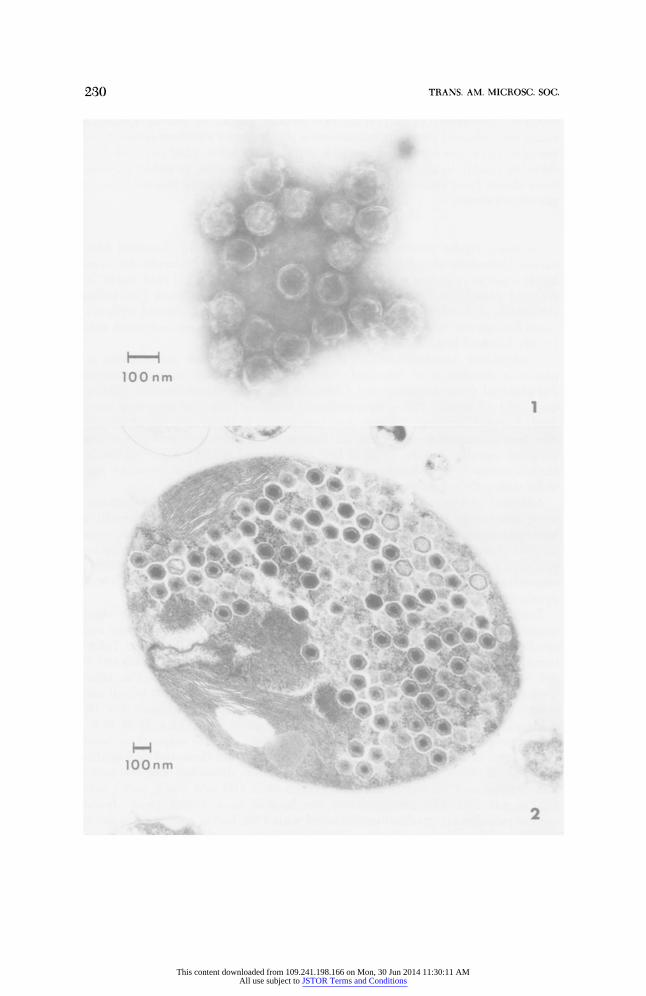
Extracts of algae isolated from hydras for 24 h produced a broad, but dis- tinct, light-scattering zone within the rate-density gradients. This zone, when collected and centrifuged through a quasi-equilibrium gradient, produced a sharp light-scattering zone 1 cm into the gradient (46% sucrose as measured with a Zeiss refractometer). The presence of viral particles within this band, similar in size and morphology to those described by Van Etten et al. (1981), was confirmed by electron microscopy (Fig. 1).
By contrast, extracts of zoochlorellae and zooxanthellae from sea anemones isolated for either 24 or 48 h did not produce any discernible light-scattering zone within rate-density gradients. Aliquots collected from rate-density gra- dients and then centrifuged through quasi-equilibrium gradients also failed to produce a discernible light-scattering zone. Virus-like particles were not de- tected by electron microscopy within pellets of material collected from rate- density gradients.
Ultrathin sections of algae isolated from hydras 24 h prior to fixation re- vealed that the algae were either lysed or, if still intact, contained numerous viral particles within the algal cytoplasm outside the chloroplast (Fig. 2). The size and morphology of the viral particles were similar to that described by Meints et al. (1981).
By contrast, profiles of virus-like particles never were seen in ultrathin sec- tions of zooxanthellae and zoochlorellae isolated from sea anemones 24 or 48
FIG. 1. Electron micrograph of negatively stained viruses obtained from isolated algae of Hydra viridis. Viruses were purified by sequential centrifugation on a sucrose rate-density gradient and a sucrose quasi-equilibrium gradient prior to staining with 0.5% ammonium molybdate. FIG. 2. Ultrathin section of an alga isolated from Hydra viridis 24 h prior to fixation for electron microscopy. Viral particles are present throughout the algal cytoplasm exluding the chloroplast.
231
This content downloaded from 109.241.198.166 on Mon, 30 Jun 2014 11:30:11 AMAll use subject to JSTOR Terms and Conditions

TRANS. AM. MICROSC. SOC.
h prior to fixation. The ultrastructure of isolated zooxanthellae and zoochlo- rellae appeared to be unchanged from the algal profiles observed within the intact symbiotic association (O'Brien, 1978).
DISCUSSION
Three strains of lytic algal viruses are described from isolated symbiotic algae of fresh-water Hydra viridis and a fourth viral strain is described from the algal symbionts of fresh-water Paramecium bursaria (see Van Etten et al., 1982). While the origin of lytic algal viruses is unknown, it does appear that these viruses are important components of symbiotic hydras and paramecia. The results presented in this report demonstrate that these lytic algal viruses have no apparent counterpart in either of two symbiotic marine algae of An- thopleura xanthogrammica. Thus, a fundamental distinction, not previously recognized, may exist between marine and fresh-water algal-invertebrate sym- biosis.
The symbiotic algae of H. viridis behave as non-permissive hosts to viral replication while maintained within hydras as symbionts. Once the algae are removed from hydras, they become permissive to viral replication which caus- es their ultimate lysis. Apparently then, hydras regulate viral production within symbiotic algae although the mechanism is unknown. Therefore, one might assume that the selective forces acting upon an algal-invertebrate symbiosis, including the regulation of a lytic virus (as in symbiotic hydras), might be different than those selective forces encountered within algal-invertebrate symbiosis where lytic viruses apparently are absent (as in A. xanthogrammica). It would be of value of know at this point if any marine algal-invertebrate symbiosis includes a lytic algal virus analogous to those observed within fresh- water hydras and paramecia.
LITERATURE CITED
MACLEOD, R. 1967. The preparation of plant viruses for use as antigens. In Williams, C. A. & Chase, M. W., eds., Methods in Immunology and Immunochemistry, Academic Press, New York, pp. 102-115.
MEINTS, R. H., VAN ETTEN, J. L., KUCZMARSKI, D., LEE, K. & ANG, B. 1981. Viral infection of the symbiotic Chlorella-like alga present in Hydra viridis. Virology, 113: 698-703.
MUSCATINE, L. 1971. Experiments on green algae coexistent with zooxanthellae in sea anemones. Pac. Sci., 25: 13-21.
MUSCATINE, L. & LENHOFF, H. M. 1965. Symbiosis of hydra and algae. I. Effects of some environmental cations on growth of symbiotic and aposymbiotic hydra. Biol. Bull. (Woods Hole), 128: 415-424.
O'BRIEN, T. L. 1978. An ultrastructural study of zoochlorellae in a marine coelenterate. Trans. Am. Microsc. Soc., 97: 320-329.
TRENCH, R. K. 1979. The cell biology of plant-animal symbiosis. Ann. Rev. Plant Physiol., 30: 485-531.
VAN ETTEN, J. L., MEINTS, R. H., BURBANK, D. E., KUCZMARSKI, D., CUPPELS, D. A. & LANE, L. C. 1981. Isolation and characterization of a virus from the intracellular green alga sym- biotic with Hydra viridis. Virology, 113: 704-711.
VAN ETTEN, J. L., MEINTS, R. H., KUCZMARSKI, D., BURBANK, D. E. & LEE, K. 1982. Viruses of
symbiotic Chlorella-like algae isolated from Paramecium bursaria and Hydra viridis. Proc. Natl. Acad. Sci. U.S.A., 79: 3867-3871.
232
This content downloaded from 109.241.198.166 on Mon, 30 Jun 2014 11:30:11 AMAll use subject to JSTOR Terms and Conditions