a novel enzyme immunoassay of anti-insulin igg in guinea pig serum
TRANSCRIPT

Vol. 147, No. 2, 1987 September 15, 1987
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Pages 644-649
A NOVEL ENZYME IMMUNOASSAY OF ANTI-INSULIN IgG IN GUINEA PIG SERUM
Takeyuki Kohno and Eiji Ishikawa
Department of Biochemistry, Medical College of Miyazaki, Kiyotake, Miyazaki 889-16, Japan
Received July 29, 1987
A novel enzyme immunoassay of anti-insulin IgG in guinea pig serum is described. Guinea pig anti-insulin serum diluted with nonspecific guinea pig serum was incubated with dinitrophenyl biotinyl nonspecific rabbit IgG-insulin conjugate and a rabbit (anti-dinitrophenyl bovine serum albumin) IgG-coated polystyrene ball. After washing to eliminate nonspecific guinea pig IgG in the diluted serum, the polystyrene ball was incubated with dinitro- phenyl-L-lysine to elute the complex of anti-insulin IgG and the conjugate. The eluate was incubated with an avidin-coated polystyrene ball. Finally, the amount of guinea pig anti-insulin IgG in the complex trapped onto the avidin-coated polystyrene ball was measured by incubation with rabbit (anti-guinea pig IgG) Fab'-peroxidase conjugate. This enzyme immunoassay was lO,OOO-fold more sensitive than the conventional enzyme immunoassay using insulin-coated polystyrene ball and rabbit (anti-guinea pig IgG) Fab'-peroxi- dase conjugate. o 1987 Academic Press, Inc.
In the conventional enzyme immunoassay of antibodies in serum, an
antigen-coated solid phase is incubated with antiserum to trap specific
immunoglobulins and reacted with anti-immunoglobulin antibody-enzyme conjugate
to estimate the amount of specific immunoglobulins on the solid phase (1).
The sensitivity of this assay is seriously limited by the nonspecific binding
of nonspecific immunoglobulins in antiserum to the solid phase (2). In
the conventional type of enzyme immunoassay for anti-insulin IgG in guinea pig
serum, the nonspecific binding of nonspecific guinea pig IgG was reduced by
incubation of insulin-nonspecific rabbit IgG-coated solid phase (instead of
insulin-bovine serum albumin-coated solid phase) with guinea pig anti-insulin
serum in the presence of nonspecific rabbit IgG at 20°C (instead of 37°C) (3).
However, the sensitivity was improved only 10 to loo-fold.
This paper describes a novel enzyme immunoassay for anti-insulin IgG in
guinea pig serum using dinitrophenyl biotinyl nonspecific rabbit IgG-insulin
conjugate, in which the nonspecific binding of nonspecific IgG was remarkably
reduced and the sensitivity was improved approximately lO,OOO-fold as compared
with the conventional enzyme immunoassay.
0006-291X/87 $1.50 Copyright 0 1987 by Academic Press, Inc. AN rights of reproduction in any form reserved. 644

Vol. 147, No. 2, 1987 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
MATERIALS AND METHODS
Buffers Te regularly used buffers were 0.1 M sodium phosphate buffer, pH 6.0, containing 5 mM EDTA (buffer A), 10 mM sodium phosphate buffer, pH 7.0 containing 0.1 M NaCl, 6.1 % bovine serum albumin and 0.1 % NaN (buffer B) and 10 mM sodium phosphate buffer, pH 7.0 containing 0.1 M NaCl (btffer C). Antibodies
Guinea pig anti-insulin serum and rabbit (anti-guinea pig IgG) IgG were obtained from Medical and Biological Laboratories Co., Ltd., Nagoya, Japan. Rabbit (anti-dinitrophenyl-bovine serum albumin) serum was obtained from Miles Laboratories, Ltd., Elkhart, Indiana. IgG was prepared by fractionation of
cellulose (4f sera with Na SO4 followed by passage through a column of diethylaminoethyl-
F(ab') was prepared by digestion of IgG with pepsin, and Fab' was prepared by redu&ion of F(ab')2 (4). Assay of peroxidase activity
The activity of peroxidase was assayed by fluorimetry at 30°C using 3-(4-hydroxyphenyl)propionic acid as a substrate (5). The fluorescence intensity was measured relative to lvg/ml quinine in 50 mM H2S04 (5). Preparation of protein-coated polystyrene balls
Polystyrene balls (3.2 mm in diameter, Precision Plastic Ball Co., Chicago, Illinois) were coated with rabbit (anti-dinitrophenyl-bovine serum albumin) IgG, avidin (Avidin D, Vector Laboratories, Inc., Burlingame, California) or bovine serum albumin by physical adsorption (6). The amount of avidin was calculated from thf extinction coefficient to be 1.54 g
absorba_nfe at 282 nm by taking the .liter.cm (7). Insulin-bovine serum
albumin-coated polystyrene balls were prepared by treatment of bovine serum albumin-coated polystyrene balls with glutaraldehyde followed by reaction with porcine insulin (3). Preparation of dinitrophenyl biotinyl nonspecific rabbit IgG-insulin conjugate
1. Preparation of maleimide-nonspecific rabbit IgG. Maleimide groups were introduced into nonspecific rabbit IgG using N-succinimidyl-6-maleimido- hexanoate (4, 9). The average number of maleimide groups introduced per nonspecific rabbit IgG molecule was 16 (4).
2. Preparation of N-biotinyl-2-mercaptoethylamine. An aliquot (0.1 ml) of 44 mM biotin-N-hydroxysuccinimide (Zymed Laboratories, Inc., San Fran- cisco, California) was incubated with 1.0 ml of 4.4 mM 2-mercaptoethylamine in 0.1 M sodium phosphate buffer, pH 7.0, containing 5 mM EDTA at 30°C for 30 min. After incubation, 0.1 ml of 1 M Tris-HCl buffer, pH 7.0, was added to the reaction mixture to eliminate remaining biotin-N-hydroxysuccinimide.
3. Preparation of biotinyl nonspecific rabbit IgG. An aliquot (0.22 ml) of the N-biotinyl-2-mercaptoethylamine solution was incubated with maleimide-nonspecific rabbit IgG (10 mg) in 2.0 ml of buffer A at 30°C for 30 min. After incubation, 0.05 ml of 0.1 M E-mercaptoethylamine in buffer A was added to the reaction mixture to block remaining maleimide groups, and the mixture was subjected to gel filtration on a column (1.0 x 30 cm) of Sephadex G-25 using 0.1 M sodium phosphate buffer, pH 7.5. The average number of biotin residues introduced per nonspecific rabbit IgG molecule was 9.7, which was calculated from the decrease in the number of maleimide groups (4).
4. Preparation of mercaptosuccinylated biotinyl nonspecific rabbit IgG. Thiol groups were introduced into biotinyl nonspecific rabbit IgG using S-acetylmercaptosuccinic anhydride (4). The average number of thiol groups introduced per biotinyl nonspecific rabbit IgG molecule was 17 (4).
5. Preparation of maleimide-dinitrophenyl-L-lysine. An aliquot (1.0 ml) of 5.5 mM dinitrophenyl-L-lysine.HCl (Tokyo Kasei Kogyo, Co., Ltd., Tokyo, Japan) in 0-1 M sodium phosphate buffer, pH 7.0, was incubated with 0.1 ml of 5.5 mM N-succinimidyl-6-maleimidohexanoate at 30°C for 30 min.
6. Preparation of mercaptosuccinylated dinitrophenyl biotinyl nonspecific rabbit IgG. An aliquot (0.59 ml) of the maleimide-dinitrophenyl-L-lysine solution was incubated with mercaptosuccinylated biotinyl nonspecific rabbit IgG (4.4 mg) in 3.1 ml of buffer A at 30°C for 30 min. The reaction mixture was subjected to gel filtration on a column (1.0 x 30 cm) of Sephadex
645

Vol. 147, No. 2, 1987 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
G-25 using buffer A. The average number of dinitrophenyl groups introduced per mercaptosuccinylated biotinyl nonspecific rabbit IgG molecule was 7.3, which was calculated from the absorb_afce at 36-q nm by taking the molar extinction coefficient to be 17,400 mol .liter.cm (8).
7. Preparation of maleimide-insulin. Maleimide groups were introduced into porcine insulin (Actrapid MC, Novo Industri A/S, Copenhagen, Denmark) using N-succinimidyl-6-maleimidohexanoate (4, 9). The average number of maleimide groups introduced per porcine insulin molecule was 0.23 (3, 4).
8. Preparation of dinitrophenyl biotinyl nonspecific rabbit IgG-insulin conjugate. Maleimide-insulin (1.2 mg) in 0.3 ml of buffer A was incubated with mercaptosuccinylated dinitrophenyl biotinyl nonspecific rabbit IgG (0.73 mg) in 0.2 ml of buffer A at 4'C for 20 h. (The average number of thiol groups per mercaptosuccinylated dinitrophenyl biotinyl nonspecific rabbit IgG molecule was 7.) After incubation, the reaction mixture was subjected to gel filtration on a column (1.5 x 45 cm) of Ultrogel AcA 34 (LKB, Stockholm, Sweden) using 0.1 M sodium phosphate buffer, pH 6.5. The average number of insulin molecules conjugated per dinitrophenyl biotinyl nonspecific rabbit IgG molecule was 5.8. The amount of dinitrophenyl biotinyl nonspecific rabbit IgG was calculated from the absorbance at 360 nm, and the number of insulin molecules introduced was calculated from the absorbance at 280 nm. Preparation of rabbit (anti-guinea pig IgG) Fab'-peroxidase conjugates
Rabbit (anti-guinea pig IgG) Fab' was conjugated to horseradish peroxi- dase using N-succinimidyl-4-(N-maleimidomethyl)cyclohexane-l-carboxylate (4). The amount of the conjugate was calculated from peroxidase activity (4). Present enzyme immunoassay
A rabbit (anti-dinitrophenyl bovine serum albumin) IgG-coated polyst rene ball was treated with nonspecific rabbit IgG as described previously (3 f and subsequently incubated with guinea pig anti-insulin serum (0.01 ml), which had been diluted with nonspecific guinea pig serum, dinitrophenyl biotinyl non- specific rabbit IgG-insulin conjugate (30 fmol/tube) and nonspecific rabbit IgG (0.3 mg/tube) in 0.09 ml of buffer B and 0.05 ml of 10 mM sodium phosphate buffer, pH 7.0, containing 1.0 M NaCl and 0.1 % bovine serum albumin and 0.1 % NaN at 20°C for 4 h. After incubation, the polystyrene ball was washed twi 3 e by addition and aspiration of 2 ml of buffer C and incubated with 1 mM dinitrophenyl-L-lysine in 0.15 ml of buffer B at room temperature overnight in the presence of nonspecific rabbit IgG (0.3 mg/tube) to elute the complex of anti-insulin IgG and the conjugate. After removal of the polystyrene ball, the eluate was incubated with an avidin-coated polystyrene ball, which had been treated with nonspecific rabbit IgG (3), at 20°C for 3 h. After incu- bation, the polystyrene ball was washed twice with buffer C as described above, and incubated with rabbit (anti-guinea pig IgG) Fab'-peroxidase conju- gate (50 ng) in 0.15 ml of 10 mM sodium phosphate buffer, pH 7.0, containing 0.1 M NaCl and 0.1 % bovine serum albumin at 20°C for 3 h. Finally, the polystyrene ball was washed twice with buffer B as described above, and bound peroxidase activity was assayed for 10 min as described above. Conventional enzyme immunoassay
An insulin-bovine serum albumin-coated polystyrene ball was incubated with guinea pig anti-insulin serum, whi$ had been diluted to various extents with nonspecific guinea pig serum and 10 -fold with buffer B to a final volume of 0.15 ml, and then with rabbit (anti-guinea pig IgG) Fab'-peroxidase conju- gate (3). Expression of the detection limit of anti-insulin IgG in guinea pig serum
The detection limit of anti-insulin IoG in ouinea piq serum was eXPreSSed as the maximal dilution of guinea pig anti-insulin serum with nonspecific guinea pig serum or the minimal amount of anti-insulin IgG which gave a bound peroxidase activity significantly in excess of that nonspecifically bound in the absence of guinea pig anti-insulin serum or IgG (background). The existence of a significant difference from the background was confirmed by the t-test (0 < 0.001. n=51. xffinity-purificationof guinea pig anti-insulin IgG
Guinea pig anti-insulin IgG was affinity-purified by elution from a column of insulin-Sepharose 48 (10).
646

Vol. 147, No. 2, 1987 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
RESULTS AND DISCUSSION
The present enzyme immunoassay is schematically shown in Fig. 1.
Dinitrophenyl biotinyl nonspecific rabbit IgG-insulin conjugate was incubated
with guinea pig anti-insulin serum, which had been diluted to various extents
with nonspecific guinea pig serum, and a rabbit (anti-dinitrophenyl bovine
serum albumin) IgG-coated polystyrene ball in the presence of nonspecific
rabbit IgG. In this step, the complex formed between anti-insulin IgG and
dinitrophenyl biotinyl nonspecific rabbit IgG-insulin conjugate was trapped
onto the polystyrene ball. After washing to eliminate nonspecific guinea
pig IgG in the diluted serum, the polystyrene ball was incubated with dinitro-
phenyl-L-lysine to elute the complex. After removal of the polystyrene
ball, the eluate was incubated with an avidin-coated polystyrene ball in the
presence of nonspecific rabbit IgG to trap the complex. After washing, the
polystyrene ball was incubated with rabbit (anti-guinea pig IgG) Fab'-peroxi-
I Ftb tgG - DNP
I Dinitrophenyl biotinyl nonspecific rabbit IgG-insulin conjugate
Siotin
I
Incubate with guinea pig anti-insulin serum (Gp antitinsulin IgG), nonspecific rabbit IgG and rabbit (anti-dinitrophenyl bovine serum albumin) IgG-coated polystyrene ball (Anti-DNP-PS Ball)
Insulin - Gp anti-insulin IgG I
Rb IgG - DNP - - - Anti-DNP-PS Ball
Bidtin
I
After washing, incubate with dinitrophenyl-L-lysine and nonspecific rabbit IgG, and remove Anti-DNP-PS Ball
Insulin - - - GP anti-insulin IgG I
RblgGDNP I
Biotin
1 Incubate with avidin-coated polystyrene ball (Avidin-PS Ball)
Insulin - - - Gp anti-insulin IgG I
RbtgG-DNP I
Biotin - - - Avidin-PS Ball
I
After washing, incubate with rabbit (anti-guinea pig IgG) Fab-peroxidase conjugate
(Rb (anti-Gp IgG) Fab’-POD)
Insulin - - - Gp anti-insulin IgG - - - Rb (anti-Gp IgG) Fab-POD
&G - DNP
B&in - - - Avidin-PS Ball
Fig. 1. Procedure of the present enzyme itmnunoassay for anti-insulin IgG in guineapig serum.
647
Vol. 147, No. 2, 1987 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
4co lo6 lo4 18
Dilution of Anti-Insulin Serum with Nonspecific Serum f-fold)
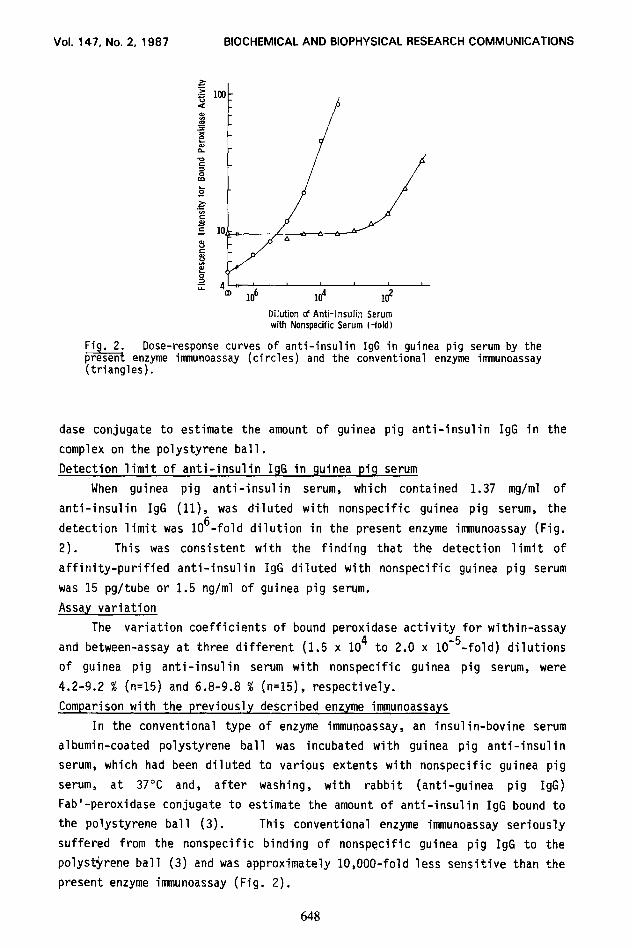
Fig. 2. Dose-response curves of anti-insulin IgG in guinea pig serum by the present enzyme immunoassay (circles) and the conventional enzyme innnunoassay (triangles).
dase conjugate to estimate the amount of guinea pig anti-insulin IgG in the
complex on the polystyrene ball.
Detection limit of anti-insulin IgG in guinea pig serum
When guinea pig anti-insulin serum, which contained 1.37 mg/ml of
anti-insulin IgG (ll), was diluted with nonspecific guinea pig serum, the
detection limit was 106-fold dilution in the present enzyme immunoassay (Fig.
2). This was consistent with the finding that the detection limit of
affinity-purified anti-insulin IgG diluted with nonspecific guinea pig serum
was 15 pg/tube or 1.5 rig/ml of guinea pig serum.
Assay variation
The variation coefficients of bound peroxidase activity for within-assay
and between-assay at three different (1.5 x lo4 to 2.0 x 10m5-fold) dilutions
of guinea pig anti-insulin serum with nonspecific guinea pig serum, were
4.2-9.2 % (n=15) and 6.8-9.8 % (n=15), respectively.
Comparison with the previously described enzyme immunoassays
In the conventional type of enzyme immunoassay, an insulin-bovine serum
albumin-coated polystyrene ball was incubated with guinea pig anti-insulin
serum, which had been diluted to various extents with nonspecific guinea pig
serum, at 37°C and, after washing, with rabbit (anti-guinea pig IgG)
Fab'-peroxidase conjugate to estimate the amount of anti-insulin IgG bound to
the polystyrene ball (3). This conventional enzyme immunoassay seriously
suffered from the nonspecific binding of nonspecific guinea pig IgG to the
polyst$rene ball (3) and was approximately lO,OOO-fold less sensitive than the
present enzyme immunoassay (Fig. 2).
648

Vol. 147,No. 2, 1987 8lOCHEMlCALAND8lOPHYSlCALRESEARCHCOMMUNlCATlONS
In the improved conventional enzyme immunoassay, an insulin-nonspecific
rabbit IgG-coated polystyrene ball was incubated with guinea pig anti-insulin
serum, which had been diluted to various extents with nonspecific guinea pig
serum, in the presence of nonspecific rabbit IgG at 20°C and, after washing,
with rabbit (anti-guinea pig IgG) Fab'-peroxidase conjugate (3). This
improved conventional enzyme immunoassay was approximately 30-fold more
sensitive than the conventional enzyme immunoassay but 300-fold less sensitive
than the present enzyme immunoassay.
Applicability of the present enzyme immunoassay
The principle of the present enzyme immunoassay makes it possible to
sensitively measure antibodies for most kinds of haptens and antigens with
identification of immunoglobulin classes.
1.
2.
::
5.
6. 7. a.
9.
:::
REFERENCES
Voller, A., Bidwell, D. E. and Grundy, S. (1983) In Immunoenzymatic Techniques (S. Avrameas, P. Druet, R. Masseyeff, and G. Feldmann, eds.) pp. 263-271 Elsevier Science Publishers B.V., Amsterdam, Netherlands. Kato, K., Haruyama, Y., Hamaguchi, Y. and Ishikawa, E. (1978) J. Biochem. 84, 93-102. Kohno, T., Hashida, S. and Ishikawa, E. (1985) J. Biochem. 98, 379-384. Ishikawa, E., Imagawa, M., Hashida, S., Yoshitake, S., Hamaguchi, Y. and Ueno, T. (1983) J. Immunoassay 4, 209-327. Imagawa, M., Hashida, S., Ishikawa, E., Mori, H., Nakai, C., Ichioka, Y. and Nakajima, K. (1983) Anal. Lett. 16, 1509-1523. Ishikawa, E. and Kato, K. (1978) Stand. J. Immunol. 8 (Suppl. 7), 43-55 Green, N. M. and Toms. E. J. (1970) Biochem. J. 118, 67-70. Eisen, H. N., Carsten, M. E. and Belman, S. (1954) J. Immunol. 73, 296-308. Hashida, S., Imagawa, M., Inoue, S., Ruan, K-h. and Ishikawa, E. (1984) J. Appl. Biochem. 6, 56-63. Kohno, T. and Ishikawa, E. (1986) J. Biochem. 100, 1247-1251. Kohno, T., Ruan, K-h. and Ishikawa, E. (1986) Anal. Lett. 19, 1083-1095.
649