a comprehensive microbial insight into two-stage anaerobic digestion of food waste-recycling...
TRANSCRIPT

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9
Avai lab le at www.sc iencedi rect .com
journa l homepage : www.e lsev ier . com/ loca te /wat res
A comprehensive microbial insight into two-stage anaerobicdigestion of food waste-recycling wastewater
Seung Gu Shin a, Gyuseong Han a, Juntaek Lim a, Changsoo Lee b, Seokhwan Hwang a,*aSchool of Environmental Science and Engineering, Pohang University of Science and Technology, Pohang, Gyeongbuk 790-784,
South KoreabDivision of Environmental and Water Resources Engineering, School of Civil and Environmental Engineering, Nanyang Technological
University, 50 Nanyang Avenue, Singapore 639798, Singapore
a r t i c l e i n f o
Article history:
Received 10 April 2010
Received in revised form
14 June 2010
Accepted 8 July 2010
Available online 16 July 2010
Keywords:
Anaerobic digestion
Denaturing gradient gel electropho-
resis
Food waste-recycling wastewater
Non-metric multidimensional
scaling
Two-stage process
* Corresponding author. Tel.: þ82 54 279 228E-mail address: [email protected] (S
0043-1354/$ e see front matter ª 2010 Elsevdoi:10.1016/j.watres.2010.07.019
a b s t r a c t
Microbial community structures were assessed in a two-stage anaerobic digestion system
treating food waste-recycling wastewater. The reactors were operated for 390 d at 10
different hydraulic retention times (HRTs) ranging from 25 to 4 d. Stable operation was
achieved with the overall chemical oxygen demand (COD) removal efficiency of 73.0e85.9%
at organic loading rate of up to 35.6 g COD/L$d. Performance of the acidogenic reactors,
however, changed significantly during operation. This change coincided with transition of
the bacterial community from one dominated by Aeriscardovia- and Lactobacillus amylovo-
rus-related species to one dominated by Lactobacillus acetotolerans- and Lactobacillus kefiri-
like organisms. In methanogenic reactors, the microbial community structures also
changed at this stage along with the shift from Methanoculleus- to Methanosarcina-like
organisms. This trend was confirmed by the non-metric multidimensional scaling joint
plot of microbial shifts along with performance parameters. These results indicated that
the overall process performance was relatively stable compared to the dynamic changes in
the microbial structures and the acidogenic performance.
ª 2010 Elsevier Ltd. All rights reserved.
1. Introduction Anaerobic digestion is an effective method of treating
Food waste constitutes one of the largest components of the
gross waste disposed in Korea (NIER, 2007). Korean food waste
is mostly collected separately from other wastes and recycled
as animal feed or compost. This recycling generates concen-
trated food waste-recycling wastewater (FRW) which causes
serious environmental concerns due to its quantity (3.3
million tons generated in 2008) and quality (up to 150 g
chemical oxygen demand (COD)/L) (Kim et al., 2010). Tradi-
tionally, most FRW has been discharged into the ocean, but
this adversely affects the ocean environment, so an alterna-
tive method of treating FRW is required.
2; fax: þ82 54 279 8299.. Hwang).ier Ltd. All rights reserve
different organic pollutants; it has the additional benefit of
generating combustible biogas which can be used to produce
energy. Previous reports have demonstrated that foodwastes
are desirable substrates for anaerobic digesters (Wang et al.,
2002; Heo et al., 2004; Zhang et al., 2007). In principle, FRW is
a diluted form of food waste and thus it is likely to be
manageable by anaerobic digestion (Kim et al., 2010).
Recently, methane production potential from FRW has been
successfully demonstrated in batch experiments (Lee et al.,
2009b; Behera et al., 2010). However, the application of
anaerobic processes for the treatment of FRW has not yet
been established in continuous-flow digesters; this type of
d.

Table 1 e Characteristics of the FRW.
Parameter Value (average � standarddeviation)
pH 3.68 � 0.23
COD (g/L) 136.8 � 12.1
Soluble COD (g/L) 86.5 � 4.7
TS (g/L) 79.5 � 8.7
VS (g/L) 63.4 � 8.3
VSS (g/L) 31.3 � 5.5
Carbohydrate (g/L) 17.5 � 4.9
Protein (g/L) 21.7 � 2.4
Lipid (g/L) 17.7 � 5.0
Total Kjeldahl nitrogen (g/L) 4.09 � 0.40
Total ammonia nitrogena (g/L) 0.62 � 0.07
NO2� (g/L) NDb
NO3� (g/L) ND
PO43- (g/L) 1.64 � 0.08
SO42- (g/L) 0.29 � 0.01
Acetate (g/L) 5.04 � 0.82
Propionate (g/L) 0.40 � 0.18
Butyrate (g/L) 0.19 � 0.08
Valerate (g/L) 0.06 � 0.02
Caproate (g/L) 0.05 � 0.02
Ethanol (g/L) 5.92 � 1.10
a The sum of N concentrations of free NH3 and NH4þ.
b Not detected.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9 4839
operation is dominant among field-scale digesters today
(Weiland, 2009).
Anaerobic digestion is amulti-stage biochemical process in
which complex organic materials undergo hydrolysis, acido-
genesis, acetogenesis, and methanogenesis in series. These
reactions are mediated by many species of microorganisms,
which are broadly classified into acidogenic bacteria and
methanogenic archaea (Chouari et al., 2005). These two groups
are present in a mixed anaerobic culture but they differ in
their nutritional and pH requirements, growth kinetics, and
their ability to tolerate environmental stresses (Demirer and
Chen, 2005). Therefore, phase separation into two digesters
has been claimed to enhance the overall process rate due to
the maintenance of favorable environments for, in series,
acidogens and methanogens (Yang et al., 2003). With the
proper enrichment of microbial communities, the two-stage
anaerobic process has been demonstrated to perform effi-
ciently (Liu et al., 2002; Stamatelatou et al., 2003; Yang et al.,
2003; Bouallagui et al., 2004).
Characterization of microbial community structure in
anaerobic digesters has attracted interest from engineers
because understanding of microbial behavior is essential to
improving the digestion process (Fernandez et al., 1999; Lee
et al., 2008). Recent advances in molecular techniques have
provided useful tools to analyze anaerobic communities in
different digester configurations (Leclerc et al., 2004;
Akarsubasi et al., 2005; Lee et al., 2009a; Riviere et al., 2009).
However, relatively little information is available on moni-
toring both acidogenic and methanogenic communities in
a two-stage system (Liu et al., 2002; Ueno et al., 2007). Moni-
toring microbial community structures in a two-stage anaer-
obic process can provide valuable information that can be
used to optimize conditions in series reactors to favor efficient
breakdown of wastes.
The purpose of this study was to assess the microbial
community structures through the molecular detection of
bacterial and archaeal species in a two-stage anaerobic
digestion for the treatment of FRW. The two-stage process
was operated at 10 different hydraulic retention times (HRTs)
ranging from 25 to 4 d. Themicrobial populationwas analyzed
using denaturing gradient gel electrophoresis (DGGE) of
bacterial and archaeal 16S rRNA genes. Community structures
were ordinated for further interpretation using non-metric
multidimensional scaling (NMS).
2. Material and methods
2.1. Preparation of substrate
FRW was collected from a food waste-recycling plant in
Kwangyang, South Korea. The FRWwas periodically collected,
mixed thoroughly, filtered through an 850-mm sieve, and
distributed in 20-L plastic bottles. The individual bottles were
stored at �20 �C until thawed and kept at 4 �C for use as
a feedstock. The pH of the FRWwas low (average value of 3.68;
Table 1), probably due to partial fermentation of residues
during food waste collection, processing, and storage (Heo
et al., 2004; Kim et al., 2008). The FRW was rich in
carbohydrate, protein, and lipid, the sum of which accounted
for 87.3e99.5% of volatile solids (VS).
2.2. Reactor setup and operations
Two identical sets of two-stage anaerobic digesters (D1, D2)
were operated in this study. Each set consisted of one acido-
genic reactor (working volume of 1 L, DA1 and DA2) and one
methanogenic reactor (working volume of 5 L, DM1 and DM2) in
series (D1, DA1eDM1, and D2, DA2eDM2). The reactors were
operated as a continuous stirred tank reactor (CSTR) at 35 �Cwithout pH control. At start-up, the four reactors were filled
with an anaerobic sludge (14.5 g VS/L) from a full-scale mes-
ophilic anaerobic digester receiving 95% v/v municipal
wastewater and 5% v/v foodwaste. Each set received substrate
stream semi-continuously through a peristaltic pump equip-
ped with a programmable mechanical timer. A steady state
was assumed when both the effluent COD concentration and
the methane production rate were constant (�5%) during
more than half of the HRT and over at least 7 d. HRT of the
two-stage digesters was decreased: 25, 15, 10, 8, 6.5, 5, and 4 d
for D1 (runs 1, 4, 6, 7, 8, 9, 10); 20, 17.5, and 12.5 d for D2 (runs 2,
3, 5) (Table 2).
2.3. Analytical methods
Samples were collected from all reactors 2e3 times per week.
Biogas production from DA (DA1 and DA2) was negligible; the
biogas production from DM (DM1 and DM2) was recorded daily
using an automated gas metering system (Korean patent
application number 10-2005-0025013), and analyzed using
a gas chromatograph (6890 plus, Agilent, Palo Alto, CA)
equipped with an HP-5 capillary column and a thermal

Table 2 e Operational and performance characteristics of the two-stage anaerobic digesters.
Run 1 2 3 4 5 6 7 8 9 10
Digester D1 D2 D2 D1 D2 D1 D1 D1 D1 D1
Operational days 0e166 0e166 166e229 166e229 229e278 229e272 272e304 304e332 332e356 356e390
HRT (d)
DA 4.2 3.3 2.9 2.5 2.1 1.7 1.3 1.1 0.8 0.7
DM 20.8 16.7 14.6 12.5 10.4 8.3 6.7 5.4 4.2 3.3
Total 25.0 20.0 17.5 15.0 12.5 10.0 8.0 6.5 5.0 4.0
OLRa (g COD/L$d) 5.7 7.2 8.2 9.5 10.5 13.1 16.4 20.2 26.2 35.6
MPb (L/L substrate) 42.2 � 2.5 43.6 � 2.0 44.3 � 1.2 39.5 � 1.6 40.5 � 1.7 36.2 � 1.8 34.4 � 1.5 32.4 � 1.5 30.7 � 1.8 27.1 � 2.2
COD removal (%) 84.0 � 0.9 85.9 � 1.0 84.0 � 1.3 81.1 � 1.6 80.9 � 1.6 75.5 � 2.3 78.2 � 1.2 77.8 � 1.2 75.8 � 2.5 73.0 � 1.9
a Organic loading rate; calculation based on the total (DA þ DM) reactor volume.
b Methane production.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 94840
conductivity detector. Volatile fatty acids (VFAs, C2eC6) and
ethanol were measured using an identical gas chromatograph
equipped with an Innowax capillary column and a flame
ionization detector. COD, total solids (TS), VS, and volatile
suspended solids (VSS) were determined according to the
procedures in Standard Methods (APHA, 2005). The carbohy-
drate concentration was measured using the phenolesulfuric
acidmethod (Dubois et al., 1956). Total Kjeldahl nitrogen, total
ammonia nitrogen, and protein concentrations were deter-
mined using the Kjeldahl method (APHA, 2005). Lipid
concentrations were measured using gravimetry after
extraction of lipids by solvent (chloroform: methanol 1:2 v/v)
(Bligh and Dyer, 1959). Nitrite, nitrate, phosphate, and sulfate
ion concentrations were measured using an ion chromato-
graph (790 Personal IC, Metrohm, Herisau, Switzerland)
equipped with a Metrosep A Supp 5 column.
2.4. Microbial community analysis
2.4.1. Nucleic acid extractionImmediately after sampling, 0.2 mL samples were centrifuged
twice and the supernatant was removed each time to mini-
mize concentrations of possible PCR inhibitors and DNA from
cell debris. The concentrated samples were stored frozen at
�20 �C before DNA extraction (Wells et al., 2009). Total DNA
was extracted from steady state samples using an automated
nucleic acid extractor (Magtration System 6 GC, Precision
System Science, Chiba, Japan). The purified DNA was eluted
with nuclease-free water and stored at �20 �C until use. All
extractions were carried out in duplicate.
2.4.2. DGGE and phylogenetic analysisBacterial and archaeal community structures were investi-
gated using DGGE, targeting the 16S rRNA gene. Conventional
PCR was performed with domain-level universal primers:
BAC338F (50-ACTCCTACGGGAGGCAG-30) and BAC805R (50-GACTACCAGGGTATCTAATCC-30) (Lee et al., 2008) for bacteria;
ARC787F (50-ATTAGATACCCSBGTAGTCC-30) and ARC1059R
(50-GCCATGCACCWCCTCT-30) for archaea. The 50 ends of the
forward primers were capped with 40-bp GC-clamps, 50-CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGG-30
for bacteria and 50-CGCCCGCCGCGCCCCGCGCCCGTCCCGC-CGCCCCCGCCCG-30 for archaea, to stabilize the melting
behavior of the PCR products (Muyzer et al., 1993). The PCR
mixture of 50 mL was prepared using the SolGent Taq DNA
Polymerase kit (Solgent, Daejon, South Korea): 5 mL of
10� reaction solution, 2 mL of each primer (final concentration
400 nM), 1 mL of dNTP mixture (final concentration 200 mM
each), 0.25 mL of Taq polymerase, 5 mL of template DNA, and
34.75 mL of PCR-grade water. A touch-down PCR was con-
ducted according to the following protocol: (1) initial dena-
turation at 94 �C for 10 min; (2) 20 cycles of denaturation at
94 �C for 30 s, annealing at 65e55 �C (reducing the temperature
by 0.5 �C per cycle) for 30 s, and extension at 72 �C for 1min; (3)
an additional 15 cycles of 94 �C for 30 s, 55 �C for 30 s, and 72 �Cfor 1 min; (4) final extension at 72 �C for 30 min. DGGE was
performed using a DCode system (Bio-Rad, Hercules, CA).
Duplicate DNA samples were used as template, and the
mixture (1:1, v/v) of the duplicate PCR products was loaded
onto an 8% (w/v) acrylamide gel containing a 30e60% dena-
turant gradient, where 100%was defined as 7 M urea with 40%
(v/v) formamide. Electrophoresis was run at 150 V for 7 h in
1 � TAE buffer. After staining with ethidium bromide, visible
bandswere excised and elutedwith distilledwater. The eluted
solution was further PCR-amplified using the corresponding
primers without GC-clamps. The PCR products were purified
from 1% agarose gel and cloned into the pGEM-T Easy vector
(Promega, Madison, WI). The cloned 16S rRNA gene fragments
were sequenced and the results were compared with refer-
ence sequences in the GenBank database using the BLAST
program (http://blast.ncbi.nlm.nih.gov/). Hierarchical classi-
fication of the 16S rRNA gene sequences was also assisted by
“Classifier” from the Ribosomal Database Project (Cole et al.,
2009). Neighbor-joining trees were constructed for
phylogenetic analysis using MEGA 4 software (Tamura et al.,
2007). The nucleotide sequences reported in this study
have been deposited under GenBank accession numbers
HM066947eHM066994.
2.4.3. OrdinationOrdination offers a view into high-dimensional spaces by plot-
ting the strongest structure into reduceddimensions (Falk et al.,
2009).NMSis themostgenerally effectiveordinationmethod for
ecological community data because NMS avoids distributional
assumptions commonly associated with other ordination
techniques (McCune and Grace, 2002). In this study, NMS ordi-
nation based on the Sorensen distancemeasurewas performed
in PC-ORD software (MjM Software Design, Gleneden Beach,

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9 4841
OR).ThepresenceorabsenceofeachDGGEbandwasscored1or
0, respectively, to generate three main matrices with rows
DAþDM,DA, andDM. Eachmainmatrixwasprocessed such that
the stress (<10) and the instability (<10�4) criteriaweremet.The
probability of a similar stress obtained by chance was deter-
mined for 40 real and 50 randomized Monte Carlo runs (Morris
et al., 2010). To evaluate the correlation between process
parameters and ordination axes in the joint plots, second
matrices were generated for joint plotting with pH, methane
production, COD and carbohydrate removal efficiency, and
concentrations of acetate, propionate, and ethanol.
3. Results
3.1. Reactor performance
Fig. 1 shows the temporal process performance of the two-
stage anaerobic digesters. Overall, stable operation of the
anaerobic system was achieved with constant production of
methane (Fig. 1a). D1 was operated for 390 d at 7 different
HRTs; D2 was operated for 278 d at 3 different HRTs (Table 2).
The performance of the anaerobic digesters was estimated at
steady states (Table 2). The concentrations of C4eC6 VFAs
were low (<0.2 g/L) in all digesters, the sum of which
accounted for < 10% of the total VFA (TVFA).
Two acidogenic reactors (DA) were operated at HRTs of
4.2e0.7 d (organic loading rate (OLR) of 34.3e213.9 g COD/L$d).
The pH in DA remained low (<4.5) after the start-up period of
15 d (Fig. 1b). The pH in DA at steady state was between 4.2 and
4.4 in runs 1e4 (at acidogenic HRTs of 4.2e2.5 d) and below 4.0
in runs 5e10 (2.1e0.7 d). Concomitant to the relatively high pH
values, a high rate of acidogenesis (i.e., net acetate
production > 13.4 g/L) was achieved in runs 1e4 (Fig. 1d). The
amount of carbohydrate degradation in DA was equivalent to
68e96% of acetate production in these runs. In contrast, net
acetate production in DA was<1.2 g/L in runs 5e10, whichwas
significantly lower (<9%) than that in runs 1e4. In all runs, net
propionate and ethanol productions in DA were estimated to
be <0.8 g/L and <2.0 g/L, respectively (Fig. 1e, f); ethanol
production was higher than TVFA production only in run 10.
The performance of the methanogenic reactors (DM) was
relatively stable throughout the experiment. Methane
production rate increased with respect to the increase in the
OLR (Fig. 1a). However, methane production per unit volume
of substrate decreased in general as HRT decreased (Table 2).
In DM, pH was maintained between 7.6 and 8.1 (Fig. 1b), which
is in the range favorable for methanogens (Speece, 1996). The
COD removal in DM was stable with efficiencies of 71.0e82.3%
(Fig. 1c). Estimated overall COD reduction efficiencies through
the two-stage system were 73.0e85.9% (Table 2). The meth-
anogenic effluent contained relatively low levels of acetate,
propionate, and ethanol (Fig. 1d, e, f). However, the propionate
concentration increased with decreasing HRT, up to
3.7 � 0.3 g/L in run 10. The methanogenic effluent also con-
tained 1.65e2.58 g/L of total ammonia nitrogen and
0.05e0.09 g/L of phosphate, indicating that nutrients (N, P)
were not limiting in this system. The sulfate concentration in
DM was measured to be 0.01e0.03 g/L.
3.2. Microbial community analysis
DGGE and subsequent phylogenetic analysis were conducted
to characterize the microbial community structures in the
two-stage anaerobic digesters. Both bacterial and archaeal
PCRs were performed with DNA samples collected when DA
andDMwere at steady state. Archaeal PCR ofDA samples failed
to produce amplification products, indicating the absence of
archaea in DA due to the suppression of methanogen growth
at low pH (Liu et al., 2002). Archaeal PCR of DM and bacterial
PCR of DA and DM samples successfully amplified 16S rRNA
gene fragments and DGGE band patterns were obtained
(Fig. 2). Phylogenetic affiliations of the 16S rRNA gene
sequences from DGGE bands were determined by comparing
against the GenBank database. Neighbor-joining trees
showing the phylogenetic identities of the 16S rRNA gene
fragments were also constructed (Figs. 3 and 4).
A total of 36bandswereexcisedandanalyzed frombacterial
DGGEs (Fig. 2a, b). Overall, bacterial band patterns in the seed
sludge, DA, and DM were distinct. The phylogenetic affiliations
of DA bacterial community converged within two phyla, Acti-
nobacteria and Firmicutes, and also within two genera, Aer-
iscardovia and Lactobacillus (Fig. 3), although the species
composition of the bacterial community shifted clearly
between runs 1 and 2e4, between runs 2e4 and 5e9, and
between runs 5e9 and 10 (Fig. 2a). B7 was one of the dominant
bands of DA in runs 1e4 (Fig. 2a). B7 was most closely (91%)
related to Aeriscardovia aeriphila, a Gram-positive, non-spore-
forming bacteria which utilizes various carbohydrates to form
acids at pH�4.2 (Simpson et al., 2004). Phylogenetic analysis in
theRibosomalDatabaseProject has classifiedB7 into the genus
Aeriscardoviawith 97%confidence, although this bandwas also
closely (up to 90%) related to Bifidobacterium spp. The band
intensity of B11 was also dominant in DA in runs 2e4. B11
showed 99% sequence similarity with Lactobacillus amylovorus,
a fermentative bacteria which produces lactic acid and small
amounts of acetic acid from sugars (Nakamura, 1981). B13 and
B15 were closely (99e100%) matched with Lactobacillus kefiri,
a heterofermentative microorganism which produces lactic
and other acids simultaneously (Marshall et al., 1984). B12
and B17 were closely related to Lactobacillus acetotolerans and
Lactobacillus suntoryeus with sequence similarity of 99% and
97%, respectively. These two species are homofermentative,
producing lactic acid as the sole fermentative product (Entani
et al., 1986; Cachat and Priest, 2005).
The bacterial community in DM included members affili-
ated within four different phyla, Firmicutes, Proteobacteria,
Spirochaetes, and Bacteroidetes, and one unaffiliated band
(B20) (Fig. 3); thus it was more diverse than the community in
DA. No band obtained from DM was related primarily to a 16S
rRNA gene sequence from an isolated microorganism.
Instead, most of the bands (B20e23, 26e36) were closely
(>97%) related to environmental clones derived from natural
and engineered environments.
The archaeal community structures in the seed sludge and
DM were visualized in Fig. 2c. The archaeal bands in DM were
affiliated within three orders, Methanomicrobiales, Meth-
anosarcinales, and Thermoplasmatales (Fig. 4). Bands similar
toMethanocorpusculumbavaricum (A1e3)were only found in the
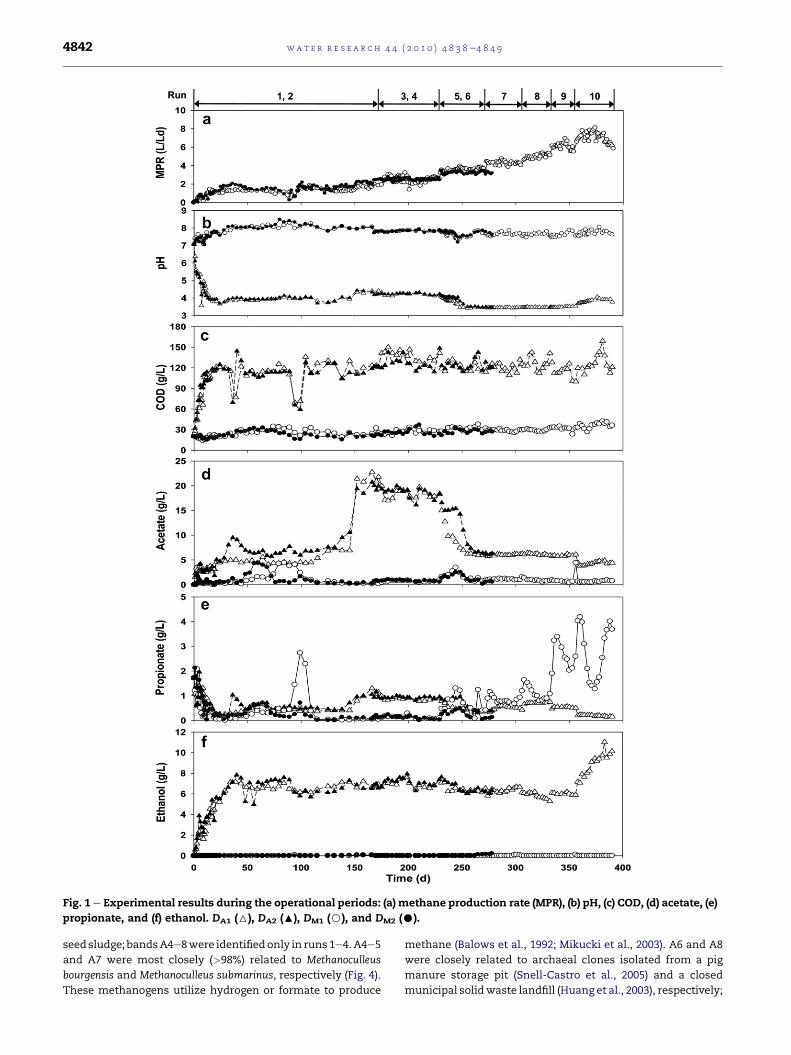
Fig. 1 e Experimental results during the operational periods: (a) methane production rate (MPR), (b) pH, (c) COD, (d) acetate, (e)
propionate, and (f) ethanol. DA1 (6), DA2 (:), DM1 (B), and DM2 (C).
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 94842
seedsludge; bandsA4e8were identifiedonly in runs1e4.A4e5
and A7 were most closely (>98%) related to Methanoculleus
bourgensis and Methanoculleus submarinus, respectively (Fig. 4).
These methanogens utilize hydrogen or formate to produce
methane (Balows et al., 1992; Mikucki et al., 2003). A6 and A8
were closely related to archaeal clones isolated from a pig
manure storage pit (Snell-Castro et al., 2005) and a closed
municipal solidwaste landfill (Huang et al., 2003), respectively;

Fig. 2 e DGGE band patterns of the 16S rRNA gene fragments: (a) bacterial profiles in DA, (b) bacterial profiles in DM, and
(c) archaealprofiles inDM.Each lane isequivalent toeitherseedsludge (seed ) oronesteadystate (indicatedbythe runnumbers).
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9 4843
they were affiliated within the order Thermoplasmatales
(Fig. 4). A9e12 showed high (>98%) sequence similarity with
the acetate-utilizingmethanogenMethanosarcina mazei; theM.
mazei-like sequence was retrieved from all experimental runs.
3.3. Ordination
NMS was used to visualize the patterns in microbial
community structure across the sample set (Fig. 5). The NMS
analysis was performed on three individual community
matrices (i.e., DA þ DM, DA, and DM). DM generated a 1-dimen-
sional resultant matrix with two distinct values: one in runs
1e4 and the other in runs 5e10 (figure not shown). Thereby,
NMS analysis on DM bands confirmed the existence of two
distinct microbial community structures, whereas a joint plot
could not be drawn.
Both NMS plots in Fig. 5 met the stress (<10) and the
instability (<10�4) criteria. The Monte Carlo test resulted in
p-values of 0.0196 for DA þ DM and 0.0392 for DA. Microbial
structures in DA and DM reactors (Fig. 5a) were clearly sepa-
rated; they were located at opposite ends of Axis 1A, which
explained 72.0% of the variability in community structures.
Community structures in each digester were also separated
along Axis 2A, which explained 21.8% of the variance.
Parameters such as pH, COD and carbohydrate reduction
efficiency, and acetate and ethanol concentrations were
highly (>0.77) correlated with Axis 1A, whereas these
parameters were less correlated with Axis 2A (<0.63).
The patterns in microbial structure in DA were illustrated
in an NMS joint plot with performance data (Fig. 5b). The
communities could be assigned to four groups corresponding
to runs 1, 2e4, 5e9, and 10. The community shift along Axis 1B
explained 63.8% of the variability in the DGGE band patterns,
and the shift along Axis 2B accounted for 29.8%. Parameters
such as pH, carbohydrate reduction efficiency, and acetate
and propionate concentrations were highly (>0.73) correlated
with longer HRT along ordination Axis 1B (Morris et al., 2010).
In contrast, ethanol concentration was slightly more corre-
lated with the sample in run 10 (at HRT 4.0 d) along Axis 2B
(0.56) than along Axis 2A (0.34).
4. Discussion
Acetate is often regarded as the most important (equivalent
to >70% of methane production) intermediate in anaerobic
digestion (Speece, 1996). Acetate can be directly utilized by the
aceticlastic methanogens such as Methanosarcina spp. There-
fore, conversion of organic materials into acetate in the first
stage of the two-stage reactors provides favorable substrate
for the methanogenic consortia in the second stage (Yang
et al., 2003). In this study, high (>13.4 g/L) production of
acetate was observed only in runs 1e4 (Fig. 1d). The difference
in the acidogenic efficiency in DA between runs 1e4 and runs
5e10 can be attributed to changes in operational factors such
as HRT. HRT is among the most crucial parameters that
determine the efficiency of an acidogenic reactor (Hwang and
Hansen, 1997; Yang et al., 2003). Reduction in HRT imposes
selective pressure on slow-growing microorganisms, which
may result in the microbial community shifts (Demirel and
Yenig, 2002). Accordingly, bacterial community structure
changed from species related to A. aeriphila and L. amylovorus,
acidogens that are capable of producing acetate, in runs 1e4
into species related to homofermentative Lactobacillus spp. in
runs 5e10 (Figs. 2a and 3). Although lactate production was
not determined in this report, the strong intensity of Lactoba-
cillus-like bands suggested that lactate may have been
involved in the degradation of the FRW. In DA, pH was not
externally controlled but was dependent upon the type and
proportion of fermentation products. The pH values in runs
1e4 were between 4.2 and 4.4; the pH values in runs 5e9 were
between 3.4 and 3.5 (Fig. 1b). Similarly, in a three-stage
anaerobic digestion of Korean food waste, the major acido-
genic product was acetate at pH 4.5e5.5 and lactate at pH

B5B7B6
Aeriscardovia aeriphila (AY174107)Bifidobacterium bifidum (U25952)
Bifidobacterium minimum (M58741)Streptomyces albus (AJ621602)
Chloroflexus aurantiacus (D38365)Thermaerobacter marianensis (AB011495)
Pseudomonas aeruginosa (X06684)Burkholderia cepacia (U96927)
Caulobacter vibrioides (AJ009957)Rhodobacter capsulatus (D16428)
B30B32
clone D23 (AM500730)Desulfovibrio cuneatus (X99501)Desulfovibrio desulfuricans (M34113)
Geobacter metallireducens (L07834)Desulforhopalus sp.lsv20 (AF099057)Desulforhopalus vacuolatus (L42613)
B3Syntrophobacter wolinii (X70905)
Myxococcus fulvus (DQ768117)Desulfobulbus propionicus (AY548789)
Syntrophus buswellii (X85131)Smithella propionica (AF126282)
Spirochaeta aurantia (AY599019)Treponema pallidum (M88726)
Treponema maltophilum (X87140)B22
clone BS12 (EU358687)B1
clone A543 (EU283591)B20
clone QEDS3CA02 (CU921669)Flammeovirga aprica (D12655)
B23clone QEEB1DF04 (CU918196)
B21Dysgonomonas gadei (Y18530)
Bacteroides fragilis (CR626927)Bacteroides nordii (AY608697)
Persicobacter diffluens (AB260929)Flavobacterium aquatile (AM230485)
B2clone 035g04.g (EU837886)
B26clone A35 D28_L_B_D03 (EF559197)
Pedobacter heparinus (AJ438172)Sphingobacterium spiritivorum (EF090267)
99
99
99
99
99
99
a
93
99
99
95
78
73
72
9899
99
99
96
98
74
99
99
73
96
98
88
0.05
Bacteroidetes
Actinobacteria
Firmicutes
Proteobacteria
Spirochaetes
Chloroflexi
unidentified
Fig. 3b
Fig. 3 e Neighbor-joining tree illustrating the phylogenetic identities of the bacterial 16S rRNA gene fragments obtained
from DGGE bands excised from seed sludge (:), DA (B), and DM (C). Numbers at nodes are bootstrap values derived from
100 analyses. Firmicutes are presented in a separate tree (b).
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 94844
3.3e3.4 (Kim et al., 2008). Ethanol is one of the important
intermediates of anaerobic digestion. Ethanol production in
the first stage of the series reactors efficiently delivers energy
to the anaerobic consortia in the second stage (Pipyn and
Verstraete, 1981). In DA, ethanol production of 1.3e1.9 g/L,
equivalent to only 1.9e2.7% of the influent COD concentration,
was observed in runs 1e4 and 10; in runs 5e9, ethanol
production was negligible. Therefore, ethanol appears to have
played a limited role in conveying organic material between
DA and DM.
The COD removal efficiencies in this process (73.0%e85.9%;
Table 2) are comparable to previous studies in which organic
removal efficiencies of two-stage CSTRs ranged from 65 to 85%
(Demirel and Yenig, 2002). This high overall performance can
be attributed to the relatively stable performance of DM
throughout the operational period (Fig. 1c, Table 2). The diver-
sity of anaerobicmicroorganisms inDMwas higher than that in
DA; 14 bacterial and 5 archaeal operational taxonomic units
were identified from DM. Relatives of the bacterial bands from
DM include different trophic groups such as sulfate-reducing
Desulfovibrio spp. (B30, B32; Fig. 3a), carbohydrate-utilizing
Dysgonomonas gadei (B21, B23), syntrophic Syntrophomonaswolfei
(B35; Fig. 3b), sugar- and peptide-utilizing Sporanaerobacter ace-
tigenes (B28, B29), and cellulolytic Clostridium phytofermentans
(B34) (Hofstad et al., 2000; Hernandez-Eugenio et al., 2002;
Warnick et al., 2002). This bacterial diversity appears to have

B10Lactobacillus amylovorus (AY944408)
B8B9B11
Lactobacillus suntoryeus (AY445815)Lactobacillus crispatus (AF257097)
B17B14
B12Lactobacillus acetotolerans (AB303841)
Lactobacillus delbrueckii (CR954253)B15
Lactobacillus kefiri (AY363303)B13B18
clone C12 (EU873789)B16
Lactobacillus pontis (AJ422032)Lactobacillus reuteri (X76328)
B19Lactobacillus panis (NR 026310)
Streptococcus pyogenes (AB002521)Bacillus subtilis (AJ276351)Desulfotomaculum thermobenzoicum (AJ276351)
Syntrophomonas wolfei (DQ666175)B35
clone 134_BE1_15 (FJ825457)B24
B25clone d18 (AB241589)
clone 318-173-HRB1 (FJ669011)Desulfitibacter alkalitolerans (AY538171)
Clostridium butyricum (AJ458420)Clostridium novyi (AB041866)Desulfotomaculum nigrificans (X62176)
Clostridium sticklandii (M26494)B28B29
Sporanaerobacter acetigenes (AF358114)B31
clone 30BF17 (AB330617)B33
clone B-85 (AY676486)B27
clone S5 (EU551089)Acetivibrio cellulolyticus (L35516)
B36clone SB27 (AB494287)
Ruminococcus flavefaciens (L76603)B34
clone 054F04.b (EU837948)Clostridium phytofermentans (AF020431)
Anaerosporobacter mobilis (AY534872)Clostridium celerecrescens (X71848)
Hespellia stercorisuis (AF445264)Ruminococcus gauvreauii (EF529620)B4
clone 24 (FJ534970)
84
b
7290
93
90
99
98
92
91
98
90
98
9999
7594
86
98
99
99
9999
98
99
99
99
77
98
0.02
Lactobacillales
Clostridiales
Bacillales
Fig. 3 e (continued).
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9 4845
resulted in themore versatile utilization of organicmaterials in
DM with lower levels of COD and VFAs. One exception to this
trend was the elevated propionate concentration in DM
compared to that in DA in runs 5e10 (Fig. 1e). Propionate is one
of the important intermediates of the anaerobic metabolism
and comprises 6e35% of the carbon balance (Scholten and
Conrad, 2000). The existence of the DGGE band (B35) poten-
tially related to the genus Desulfotomaculum, some members of
which are able to oxidize propionate (McInerney et al., 2008),
implies that propionate metabolism may have occurred in DM.
The slow-growing nature of propionate-utilizing microorgan-
isms (Gavala et al., 2003) can be responsible for the accumula-
tion of propionate at shorter HRTs. Although some bands (B30,
B32) were closely related to sulfate-reducing bacteria (Fig. 3a),
sulfate reduction in this system (0.26e0.27 g/L) accounted for
only 0.2% of the total electron flow calculated as described
previously (Isa et al., 1986).
ThearchaealDGGEresults suggested thatMethanoculleus- and
Methanosarcina-like organisms were likely to be mainly respon-
sible for the hydrogenotrophic and aceticlasticmethanogenesis,
respectively (Figs. 2c and 4). The stable methanogenic perfor-
mance in DM implies the maintenance of well-established
methanogenic consortia throughout the reaction period.
However, the archaeal community structure showed a dynamic
shift between runs 1e4 and runs 5e10 (Figs. 2c and 5a), coinci-
dental with the change of the acidogenic performance in DA.
The putative hydrogenotrophic methanogens related to Meth-
anoculleus appear to have participated in syntrophic fatty acid
mineralization by removing acetogenic products in runs 1e4
(Batstone, 2002; de Bok et al., 2004). The lack of DGGE bands

Methanomicrobiales
Methanosarcinales
Thermoplasmatales
HalobacterialesArchaeoglobales
Methanobacteriales
Methanococcales
Methanopyrales
A1A3A2
Methanocorpusculum bavaricum (AY196676)Methanocorpusculum parvum (M59147)
Methanospirillum hungatei (AY196683)A4
A5Methanoculleus bourgensis (AB065298)
A7Methanoculleus marisnigri (M59134)Methanoculleus submarinus (NR 028856)
Methanomicrobium mobile (M59142)Methanogenium cariaci (M59130)
Methanoplanus petrolearius (U76631)Methanofollis tationis (AF095272)
A9A11
A10A12
Methanosarcina acetivorans (M59137)Methanosarcina mazei (FJ155845)Methanosarcina thermophila (M59140)
Methanosarcina barkeri (AJ012094)Methanolobus tindarius (M59135)
Methanococcoides methylutens (M59127)Methanosaeta concilii (M59146)
Methanosaeta thermophila (AB071701)Methanosaeta harundinacea (AY817738)
Haloferax volcanii (AY425724)Archaeoglobus fulgidus (AE000782)
Methanobacterium formicicum (AF169245)Methanosphaera stadtmanae (CP000102)
Methanobrevibacter ruminantium (AY196666)Methanothermobacter thermautotrophicus (AY196660)
Methanopyrus kandleri (AE009439)Methanococcus voltae (M59290)Methanococcus vannielii (AY196675)
Methanocaldococcus jannaschii (L77117)A6
Uncultured archaeon clone DF86 (AY816986)A8
Uncultured archaeon clone GZK61 (AJ576235)Thermogymnomonas acidicola(AB269873)Thermoplasma acidophilum (M38637)
100
100
100
100
92
100
100
8997
84
82
84
83
100
99
78
77
95
99
99 86
89
99
88
0.05
Fig. 4 e Neighbor-joining tree illustrating the phylogenetic identities of the archaeal 16S rRNA gene fragments obtained
from DGGE bands excised from seed sludge (:) and DM (C). Numbers at nodes are bootstrap values derived from 100
analyses.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 94846
related to hydrogenotrophic methanogens in runs 5e10 was
beyond anticipation because hydrogen often accounts for about
30% of the methane production in anaerobiosis (Speece, 1996)
and the influent ethanol, the degradation of which produces
hydrogen,was completely utilized inDM (Fig. 1f). This shift in the
major DGGE bandsmay be attributed to the general limitation of
the PCR-based methods with which a numerically minor but
functionally important population could be omitted in the result
(Forney et al., 2004; Shin et al., 2008). Twoarchaeal bands (A6, A8)
were also identified in DM; they are only distantly related to the
thermoacidophilic non-methanogenic archaea Thermoplasma
acidophilum or Thermogymnomonas acidicola, so their metabolic
roles remain unclear (Itoh et al., 2007). However, many of the
bands (B20e23, B26e36, A6, A8) isolated from DM were closely
(>97%) related toenvironmental clonesderived fromnatural and
engineered environments. This suggests that these yet-to-be-
cultured organisms are ubiquitous in the environment and,
potentially,havebroadmetaboliccapabilities (Huangetal., 2003).
Operational conditions of the series digesters (DA and DM)
differed only in HRT; in both digesters, mesophilic tempera-
ture (35 �C) was maintained and pH was not controlled. At the
start-up, both DA and DM were seeded with the same meso-
philic sludge. After the long-term operation in runs 1 and 2,
however, completely different structures of the microbial
communities, also distinct from the structure in the seed
sludge, have evolved in the series reactors (Fig. 5a). As a result,
the performance of the two-stage reactors was totally
different in terms of biogas production, COD removal, and the
effluent quality. Interestingly, the performance was also
different between DA in run 1 (HRT of 4.2 d) and DM in run 9
(4.2 d) or 10 (3.3 d), where theHRT ofDMwas equal to or shorter
than that of DA. Therefore, it can be deduced that the main-
tenance of well-functioning methanogenic consortia through
a systematic acclimation to a new substrate, e.g. a stepwise
increase in OLR, is important for the successful operation of
anaerobic processes (McMahon et al., 2004; Song et al., 2010).
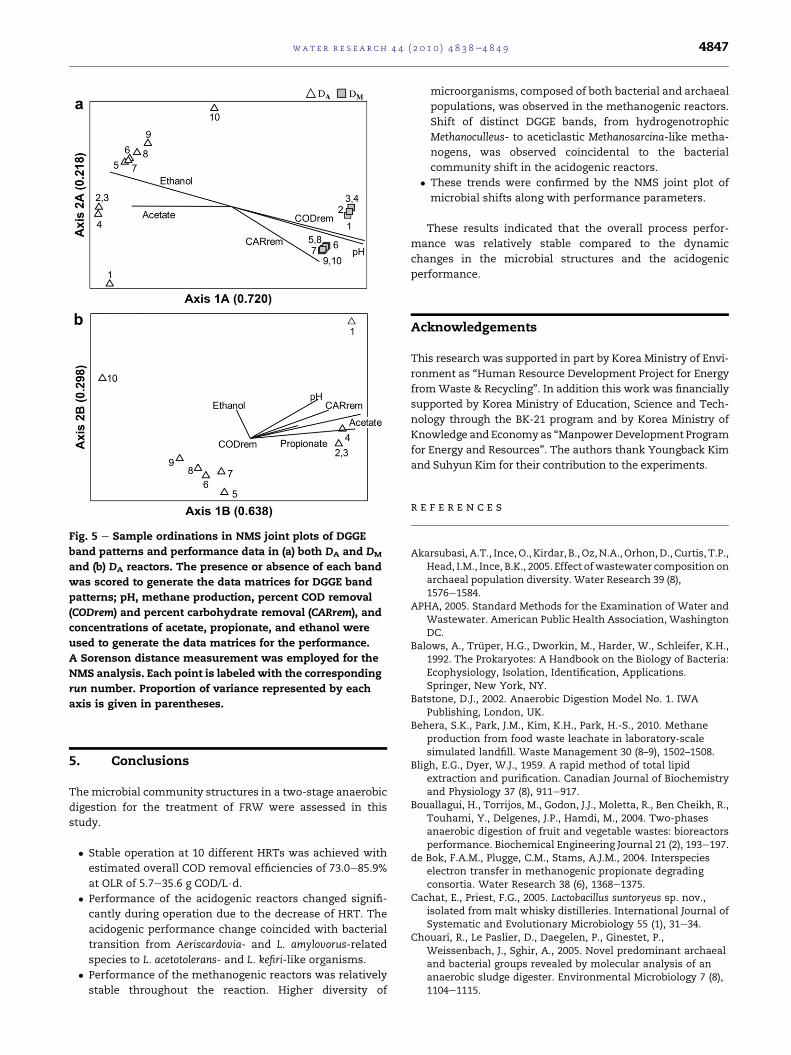
Fig. 5 e Sample ordinations in NMS joint plots of DGGE
band patterns and performance data in (a) both DA and DM
and (b) DA reactors. The presence or absence of each band
was scored to generate the data matrices for DGGE band
patterns; pH, methane production, percent COD removal
(CODrem) and percent carbohydrate removal (CARrem), and
concentrations of acetate, propionate, and ethanol were
used to generate the data matrices for the performance.
A Sorenson distance measurement was employed for the
NMS analysis. Each point is labeled with the corresponding
run number. Proportion of variance represented by each
axis is given in parentheses.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9 4847
5. Conclusions
Themicrobial community structures in a two-stage anaerobic
digestion for the treatment of FRW were assessed in this
study.
� Stable operation at 10 different HRTs was achieved with
estimated overall COD removal efficiencies of 73.0e85.9%
at OLR of 5.7e35.6 g COD/L$d.
� Performance of the acidogenic reactors changed signifi-
cantly during operation due to the decrease of HRT. The
acidogenic performance change coincided with bacterial
transition from Aeriscardovia- and L. amylovorus-related
species to L. acetotolerans- and L. kefiri-like organisms.
� Performance of the methanogenic reactors was relatively
stable throughout the reaction. Higher diversity of
microorganisms, composed of both bacterial and archaeal
populations, was observed in the methanogenic reactors.
Shift of distinct DGGE bands, from hydrogenotrophic
Methanoculleus- to aceticlastic Methanosarcina-like metha-
nogens, was observed coincidental to the bacterial
community shift in the acidogenic reactors.
� These trends were confirmed by the NMS joint plot of
microbial shifts along with performance parameters.
These results indicated that the overall process perfor-
mance was relatively stable compared to the dynamic
changes in the microbial structures and the acidogenic
performance.
Acknowledgements
This research was supported in part by Korea Ministry of Envi-
ronment as “Human Resource Development Project for Energy
fromWaste & Recycling”. In addition this work was financially
supported by Korea Ministry of Education, Science and Tech-
nology through the BK-21 program and by Korea Ministry of
Knowledge and Economyas “ManpowerDevelopment Program
for Energy and Resources”. The authors thank Youngback Kim
and Suhyun Kim for their contribution to the experiments.
r e f e r e n c e s
Akarsubasi, A.T., Ince,O.,Kirdar, B.,Oz,N.A.,Orhon,D., Curtis, T.P.,Head, I.M., Ince, B.K., 2005. Effect ofwastewater composition onarchaeal population diversity. Water Research 39 (8),1576e1584.
APHA, 2005. Standard Methods for the Examination of Water andWastewater. American Public Health Association, WashingtonDC.
Balows, A., Truper, H.G., Dworkin, M., Harder, W., Schleifer, K.H.,1992. The Prokaryotes: A Handbook on the Biology of Bacteria:Ecophysiology, Isolation, Identification, Applications.Springer, New York, NY.
Batstone, D.J., 2002. Anaerobic Digestion Model No. 1. IWAPublishing, London, UK.
Behera, S.K., Park, J.M., Kim, K.H., Park, H.-S., 2010. Methaneproduction from food waste leachate in laboratory-scalesimulated landfill. Waste Management 30 (8–9), 1502–1508.
Bligh, E.G., Dyer, W.J., 1959. A rapid method of total lipidextraction and purification. Canadian Journal of Biochemistryand Physiology 37 (8), 911e917.
Bouallagui, H., Torrijos, M., Godon, J.J., Moletta, R., Ben Cheikh, R.,Touhami, Y., Delgenes, J.P., Hamdi, M., 2004. Two-phasesanaerobic digestion of fruit and vegetable wastes: bioreactorsperformance. Biochemical Engineering Journal 21 (2), 193e197.
de Bok, F.A.M., Plugge, C.M., Stams, A.J.M., 2004. Interspecieselectron transfer in methanogenic propionate degradingconsortia. Water Research 38 (6), 1368e1375.
Cachat, E., Priest, F.G., 2005. Lactobacillus suntoryeus sp. nov.,isolated from malt whisky distilleries. International Journal ofSystematic and Evolutionary Microbiology 55 (1), 31e34.
Chouari, R., Le Paslier, D., Daegelen, P., Ginestet, P.,Weissenbach, J., Sghir, A., 2005. Novel predominant archaealand bacterial groups revealed by molecular analysis of ananaerobic sludge digester. Environmental Microbiology 7 (8),1104e1115.

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 94848
Cole, J.R., Wang, Q., Cardenas, E., Fish, J., Chai, B., Farris, R.J.,Kulam-Syed-Mohideen, A.S., McGarrell, D.M., Marsh, T.,Garrity, G.M., Tiedje, J.M., 2009. The Ribosomal DatabaseProject: improved alignments and new tools for rRNAanalysis. Nucleic Acids Research 37 (Suppl. 1), D141eD145.
Demirel, B., Yenig, O., 2002. Two-phase anaerobic digestionprocesses: a review. Journal of Chemical Technology andBiotechnology 77 (7), 743e755.
Demirer, G.N., Chen, S., 2005. Two-phase anaerobic digestion ofunscreened dairy manure. Process Biochemistry 40 (11),3542e3549.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, F.,1956. Colorimetric determination of sugars and relatedsubstances. Analytical Chemistry 28 (31), 350e356.
Entani, E., Masai, H., Suzuki, K.-I., 1986. Lactobacillus acetotolerans,a new species from fermented vinegar broth. InternationalJournal of Systematic and Evolutionary Microbiology 36 (4),544e549.
Falk, M.W., Song, K.-G., Matiasek, M.G., Wuertz, S., 2009. Microbialcommunity dynamics in replicate membrane bioreactors e
natural reproducible fluctuations. Water Research 43 (3),842e852.
Fernandez, A., Huang, S., Seston, S., Xing, J., Hickey, R., Criddle, C., Tiedje, J., 1999. How stable is stable? Function versuscommunity composition. Applied and EnvironmentalMicrobiology 65 (8), 3697e3704.
Forney, L.J., Zhou, X., Brown, C.J., 2004. Molecular microbialecology: land of the one-eyed king. Current Opinion inMicrobiology 7 (3), 210e220.
Gavala, H., Angelidaki, I., Ahring, B., 2003. Biomethanation I,pp. 57e93.
Heo, N.H., Park, S.C., Kang, H., 2004. Effects of mixture ratio andhydraulic retention time on single-stage anaerobic co-digestion of food waste and waste activated sludge. Journal ofEnvironmental Science and Health, Part A: Toxic/HazardousSubstances and Environmental Engineering 39 (7), 1739e1756.
Hernandez-Eugenio, G., Fardeau, M.L., Cayol, J.L., Patel, B.K.C.,Thomas, P., Macarie, H., Garcia, J.L., Ollivier, B., 2002.Sporanaerobacter acetigenes gen. nov., sp. nov., a novelacetogenic, facultatively sulfur-reducing bacterium.International Journal of Systematic and EvolutionaryMicrobiology 52 (4), 1217e1223.
Hofstad, T., Olsen, I., Eribe, E.R., Falsen, E., Collins, M.D.,Lawson, P.A., 2000. Dysgonomonas gen. nov. to accommodateDysgonomonas gadei sp. nov., an organism isolated froma human gall bladder, and Dysgonomonas capnocytophagoides(formerly CDC group DF-3). International Journal ofSystematic and Evolutionary Microbiology 50 (6), 2189e2195.
Huang, L.-N., Chen, Y.-Q., Zhou, H., Luo, S., Lan, C.-Y., Qu, L.-H.,2003. Characterization of methanogenic Archaea in theleachate of a closed municipal solid waste landfill. FEMSMicrobiology Ecology 46 (2), 171e177.
Hwang, S., Hansen, C.L., 1997. Modeling and optimization inanaerobic bioconversion of complex substrates to acetic andbutyric acids. Biotechnology and Bioengineering 54 (5).
Isa, Z., Grusenmeyer, S., Verstraete, W., 1986. Sulfate reductionrelative to methane production in high-rate anaerobicdigestion: microbiological aspects. Applied andEnvironmental Microbiology 51 (3), 580e587.
Itoh, T., Yoshikawa, N., Takashina, T., 2007. Thermogymnomonasacidicola gen. nov., sp. nov., a novel thermoacidophilic, cellwall-less archaeon in the order Thermoplasmatales, isolatedfrom a solfataric soil in Hakone, Japan. International Journal ofSystematic and Evolutionary Microbiology 57 (11), 2557e2561.
Kim, J.K., Han, G.H., Oh, B.R., Chun, Y.N., Eom, C.-Y., Kim, S.W.,2008. Volumetric scale-up of a three stage fermentationsystem for food waste treatment. Bioresource Technology 99(10), 4394e4399.
Kim, M.D., Song, M., Jo, M., Shin, S.G., Khim, J.H., Hwang, S., 2010.Growth condition and bacterial community for maximumhydrolysis of suspended organic materials in anaerobicdigestion of food-waste recycling wastewater. AppliedMicrobiology and Biotechnology 85 (5), 1611e1618.
Leclerc, M., Delgenes, J.-P., Godon, J.-J., 2004. Diversity of thearchaeal community in 44 anaerobic digesters as determinedby single strand conformation polymorphism analysis and16S rDNA sequencing. Environmental Microbiology 6 (8),809e819.
Lee, C., Kim, J., Hwang, K., O’Flaherty, V., Hwang, S., 2009a.Quantitative analysis of methanogenic community dynamicsin three anaerobic batch digesters treating differentwastewaters. Water Research 43 (1), 157e165.
Lee, C., Kim, J., Shin, S.G., Hwang, S., 2008. Monitoring bacterialand archaeal community shifts in a mesophilic anaerobicbatch reactor treating a high-strength organic wastewater.FEMS Microbiology Ecology 65 (3), 544e554.
Lee, D.H., Behera, S.K., Kim, J.W., Park, H.-S., 2009b. Methaneproduction potential of leachate generated from Korean foodwaste recycling facilities: a lab-scale study. WasteManagement 29 (2), 876e882.
Liu, W.T., Chan, O.C., Fang, H.H., 2002. Microbial communitydynamics during start-up of acidogenic anaerobic reactors.Water Research 36 (13), 3203e3210.
Marshall, V., Cole, W., Farrow, J., 1984. A note on theheterofermentative Lactobacillus isolated from kefir grains.Journal of Applied Bacteriology 56 (3), 503e505.
McCune, B., Grace, J.B., 2002. Analysis of Ecological Communities.MjM Software Design, Gleneden Beach, OR.
McInerney, M.J., Struchtemeyer, C.G., Sieber, J., Mouttaki, H.,Stams, A.J.M., Schink, B., Rohlin, L., Gunsalus, R.P., 2008.Physiology, ecology, phylogeny, and genomics ofmicroorganisms capable of syntrophic metabolism. Annals ofthe New York Academy of Sciences 1125 (1), 58e72. incredibleanaerobes from physiology to genomics to fuels.
McMahon, K.D., Alfons, D.Z., Stams, J.M., Mackie, R.I., Raskin, L.,2004. Microbial population dynamics during start-up andoverload conditions of anaerobic digesters treating municipalsolid waste and sewage sludge. Biotechnology andBioengineering 87 (7), 823e834.
Mikucki, J.A., Liu, Y., Delwiche, M., Colwell, F.S., Boone, D.R., 2003.Isolation of a methanogen from deep marine sediments thatcontain methane hydrates, and description of Methanoculleussubmarinus sp. nov. Applied and Environmental Microbiology69 (6), 3311e3316.
Morris, R.M., Nunn, B.L., Frazar, C., Goodlett, D.R., Ting, Y.S.,Rocap, G., 2010. Comparative metaproteomics reveals ocean-scale shifts in microbial nutrient utilization and energytransduction. The ISME Journal 4.
Muyzer, G., de Waal, E.C., Uitterlinden, A.G., 1993. Profiling ofcomplex microbial populations by denaturing gradient gelelectrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied andEnvironmental Microbiology 59 (3), 695e700.
Nakamura, L.K., 1981. Lactobacillus amylovorus, a new starch-hydrolyzing species from cattle waste-corn fermentations.International Journal of Systematic Bacteriology 31 (1),56e63.
NIER, 2007. The Third (2006e2007) National Waste Statistics Survey(제3차 (2006e2007년) 전국폐기물통계조사). National Institute ofEnvironmental Research, Incheon, Korea.
Pipyn, P., Verstraete, W., 1981. Lactate and ethanol asintermediates in two-phase anaerobic digestion.Biotechnology and Bioengineering 23 (5), 1145e1154.
Riviere, D., Desvignes, V., Pelletier, E., Chaussonnerie, S.,Guermazi, S., Weissenbach, J., Li, T., Camacho, P., Sghir, A.,2009. Towards the definition of a core of microorganisms

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 8 3 8e4 8 4 9 4849
involved in anaerobic digestion of sludge. The ISME Journal 3(6), 700e714.
Scholten, J.C.M., Conrad, R., 2000. Energetics of syntrophicpropionate oxidation in defined batch and chemostatcocultures. Applied and Environmental Microbiology 66 (7),2934e2942.
Shin, S.G., Lee, C., Hwang, K., Ahn, J.-H., Hwang, S., 2008. Use oforder-specific primers to investigate the methanogenicdiversity in acetate enrichment system. Journal of IndustrialMicrobiology and Biotechnology 35 (11), 1345e1352.
Simpson, P.J., Ross, R.P., Fitzgerald, G.F., Stanton, C., 2004.Bifidobacterium psychraerophilum sp. nov. and Aeriscardoviaaeriphila gen. nov., sp. nov., isolated from a porcine caecum.International Journal of Systematic and EvolutionaryMicrobiology 54 (2), 401e406.
Snell-Castro, R., Godon, J.-J., Delgen, J.-P., Dabert, P., 2005.Characterisation of the microbial diversity in a pig manurestorage pit using small subunit rDNA sequence analysis. FEMSMicrobiology Ecology 52 (2), 229e242.
Song, M., Shin, S.G., Hwang, S., 2010. Methanogenic populationdynamics assessed by real-time quantitative PCR in sludgegranule in upflow anaerobic sludge blanket treating swinewastewater. Bioresource Technology 101 (1, Suppl. 1), S23eS28.
Speece, R.E., 1996. Anaerobic Biotechnology for IndustrialWastewaters. Archae Press, Nashville, TN.
Stamatelatou, K., Dravillas, K., Lyberatos, G., 2003. Methaneproduction from sweet sorghum residues via a two-stageprocess. Water Science and Technology 48 (4), 235e238.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: molecularevolutionary genetics analysis (MEGA) software version 4.0.Molecular Biology and Evolution 24 (8), 1596e1599.
Ueno, Y., Fukui, H., Goto, M., 2007. Operation of a two-stagefermentation process producing hydrogen and methane fromorganic waste. Environmental Science & Technology 41 (4),1413e1419.
Wang, J.Y., Xu, H.L., Tay, J.H., 2002. A hybrid two-phase system foranaerobic digestion of food waste. Water Science andTechnology 45 (12), 159e165.
Warnick, T.A., Methe, B.A., Leschine, S.B., 2002. Clostridiumphytofermentans sp. nov., a cellulolytic mesophile from forestsoil. International Journal of Systematic and EvolutionaryMicrobiology 52 (4), 1155e1160.
Weiland, P., 2009. Biogas production: current state andperspectives. Applied Microbiology and Biotechnology.
Wells, G.F., Park, H.-D., Yeung, C.-H., Eggleston, B., Francis, C.A.,Criddle, C.S., 2009. Ammonia-oxidizing communities ina highly aerated full-scale activated sludge bioreactor:betaproteobacterial dynamics and low relative abundance ofCrenarchaea. Environmental Microbiology 11 (9), 2310e2328.
Yang, K., Yu, Y., Hwang, S., 2003. Selective optimization inthermophilic acidogenesis of cheese-whey wastewater toacetic and butyric acids: partial acidification andmethanation. Water Research 37 (10), 2467e2477.
Zhang,R., El-Mashad,H.M.,Hartman,K.,Wang,F., Liu,G.,Choate,C.,Gamble, P., 2007. Characterization of foodwaste as feedstock foranaerobic digestion. Bioresource Technology 98 (4), 929e935.