4. vorlesung rädler ws 2010 - lmu münchen · 4. vorlesung rädler ws 2010 28. oct. 2010 protein...
TRANSCRIPT

Biophysik der Moleküle
4. Vorlesung Rädler WS 2010
28. Oct. 2010
Protein folding- Afinsen hypothesis- hydrophobic interaction

Gaub/SS 2005 BPM §1.3 2
Protein Unfolding: Sushi Restaurant
When foods with proteins are exposed to heat and certain chemicals (such as vinegar), they turn white.
1. Distinguish salmon roe from imitation salmon
roe by dropping into hot tea.
2. Mackerel is pickled in vinegar for
preservation.

Gaub/SS 2005 BPM §1.3 3

Nobel Prize for Chemistry in 1972
C. Afinsen 1916-1995
The Thermodynamic Hypothesis (Afinsen 1973)
„the native state is thermodynamically stable“
=> the sequence alone
determines 3D structure!
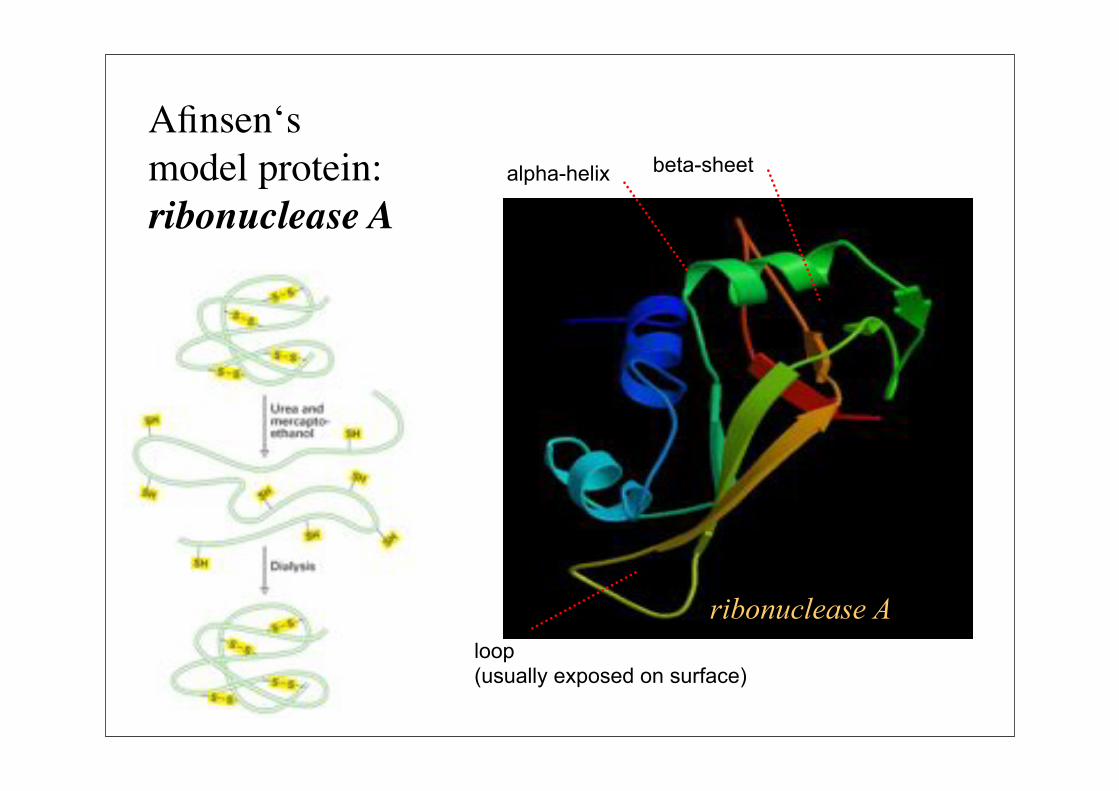
ribonuclease A
loop(usually exposed on surface)
alpha-helix beta-sheet
Afinsen‘smodel protein:ribonuclease A
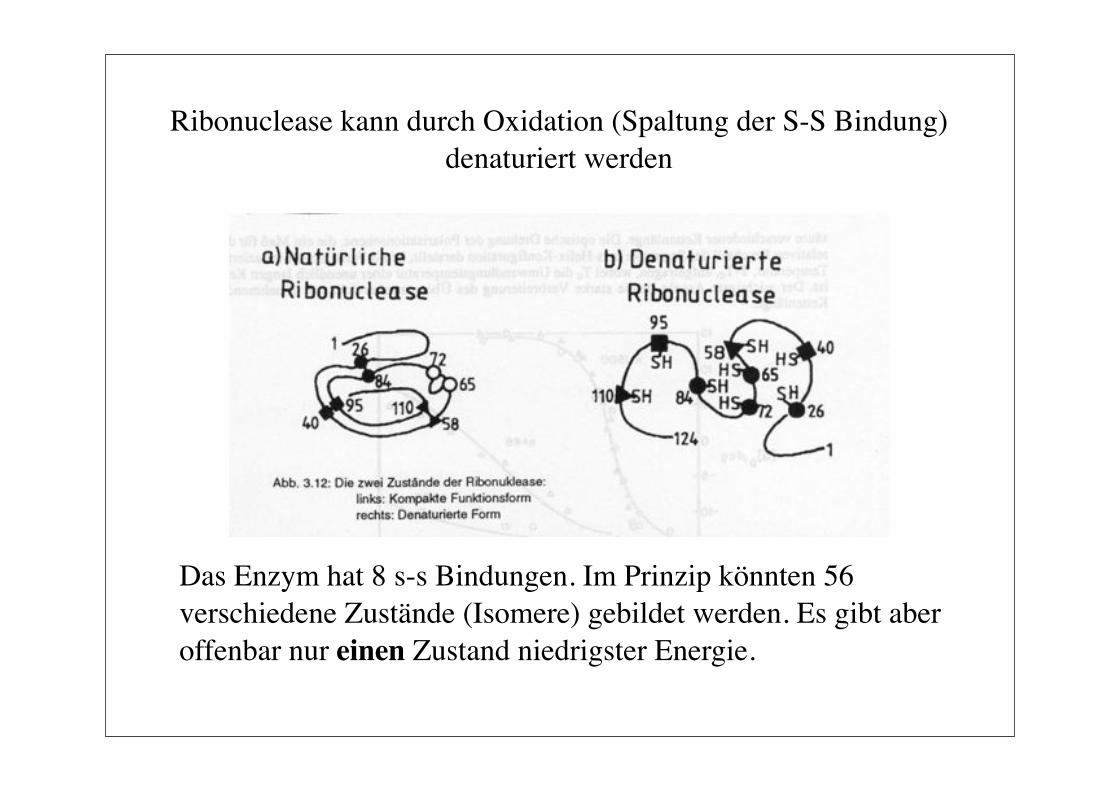
Ribonuclease kann durch Oxidation (Spaltung der S-S Bindung)denaturiert werden
o) Nofür t iche
Ribonuclea se
Abb. 3.12: Die zwei Zuslände der Ribonuklease:
links: KomDakt! Funktionsfom
rechts: Oenatudert! Form
b )0eno tu r ie r te
Ribonu cleose95
HH
65
Das Enzym hat 8 s-s Bindungen. Im Prinzip könnten 56 verschiedene Zustände (Isomere) gebildet werden. Es gibt aber offenbar nur einen Zustand niedrigster Energie.

7
Folding of RNAse A in the test tube
denaturation renaturation
Incubate proteinin guanidine
hydrochloride(GuHCl)or urea
100-folddilution of proteininto physiological
buffer
Anfinsen, CB (1973) Principles that govern the folding of protein chains.
Science 181, 223-230.
- the amino acid sequence of a polypeptide is sufficient to specify its three-dimensional conformation
Thus: “protein folding is a spontaneous process that does not require the assistance of extraneous factors”
(aggregation)
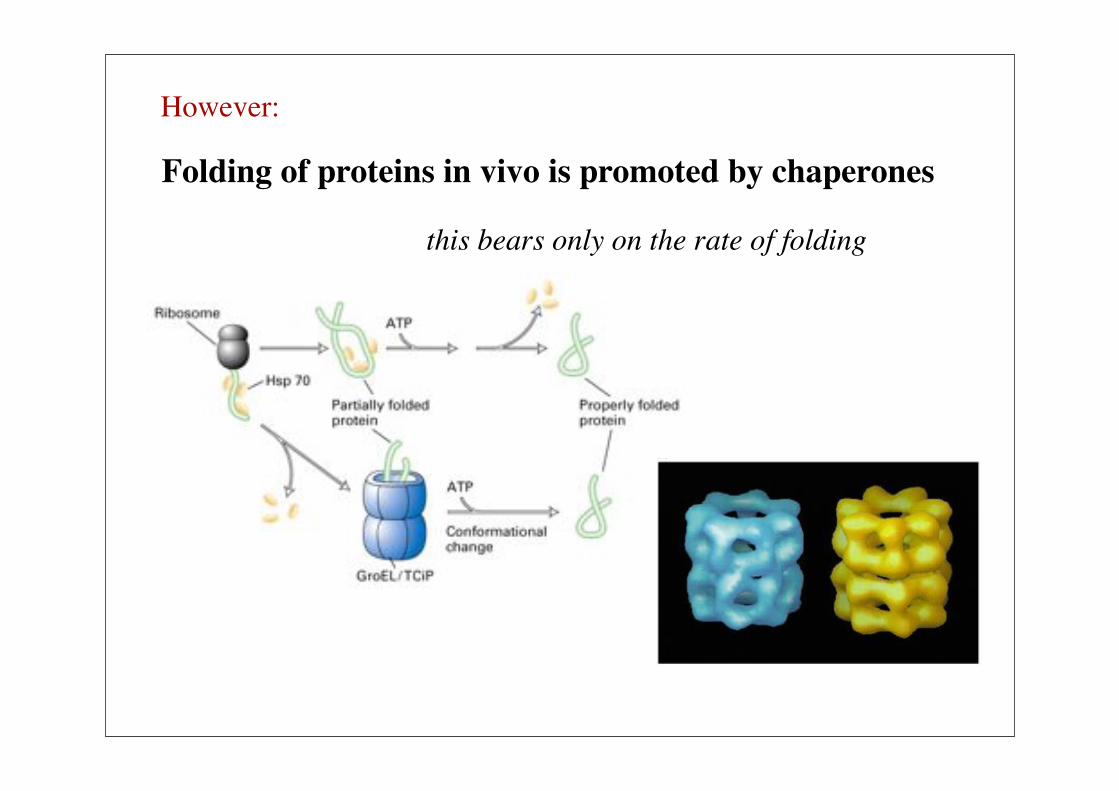
Folding of proteins in vivo is promoted by chaperones
this bears only on the rate of folding
However:
9
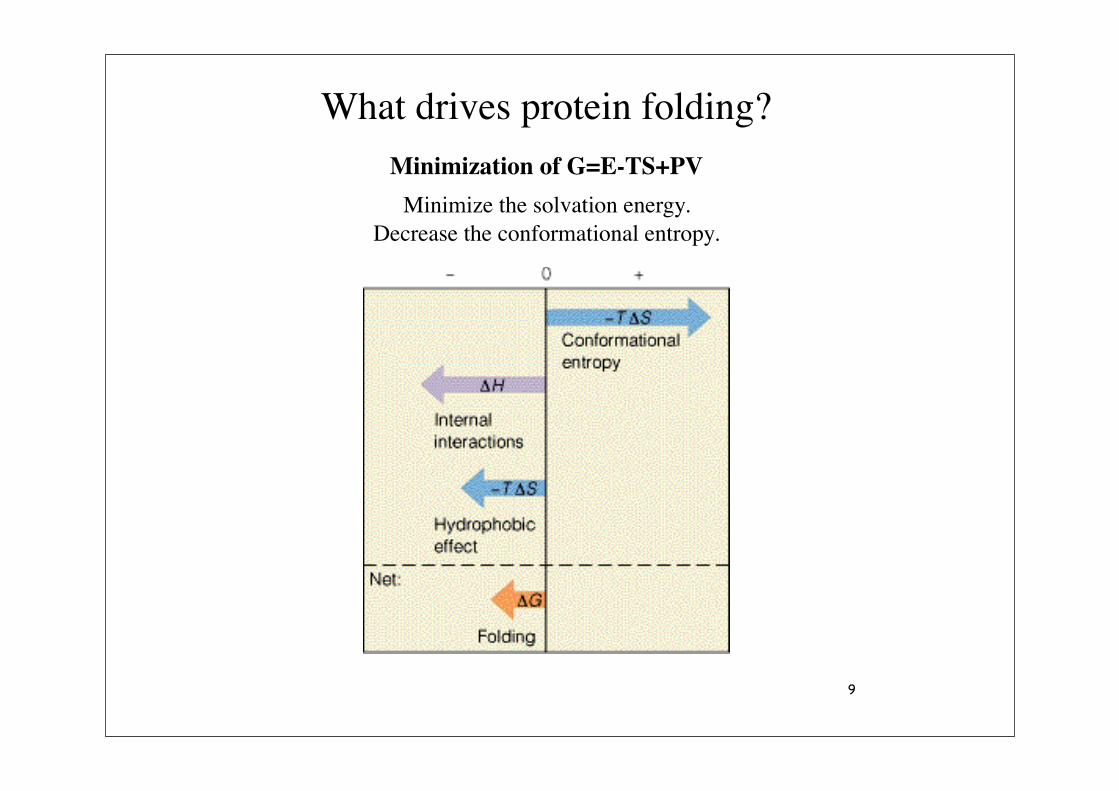
What drives protein folding?
Minimization of G=E-TS+PV
Minimize the solvation energy.Decrease the conformational entropy.

10
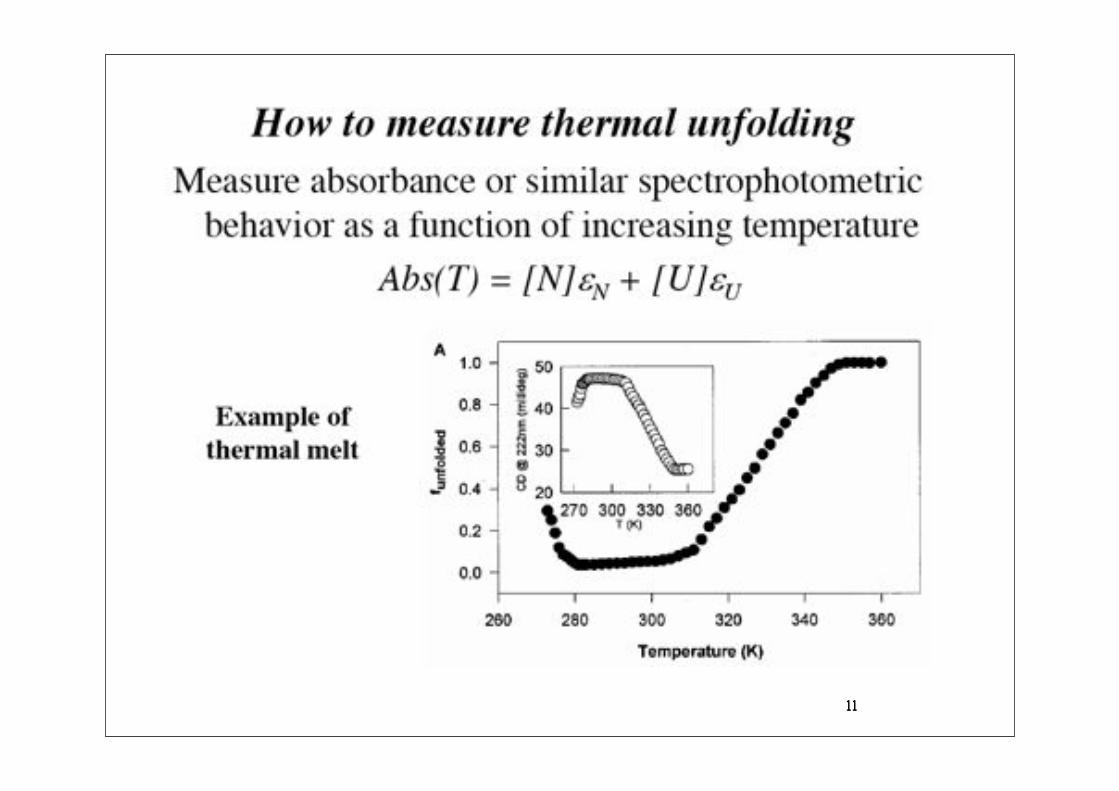
11
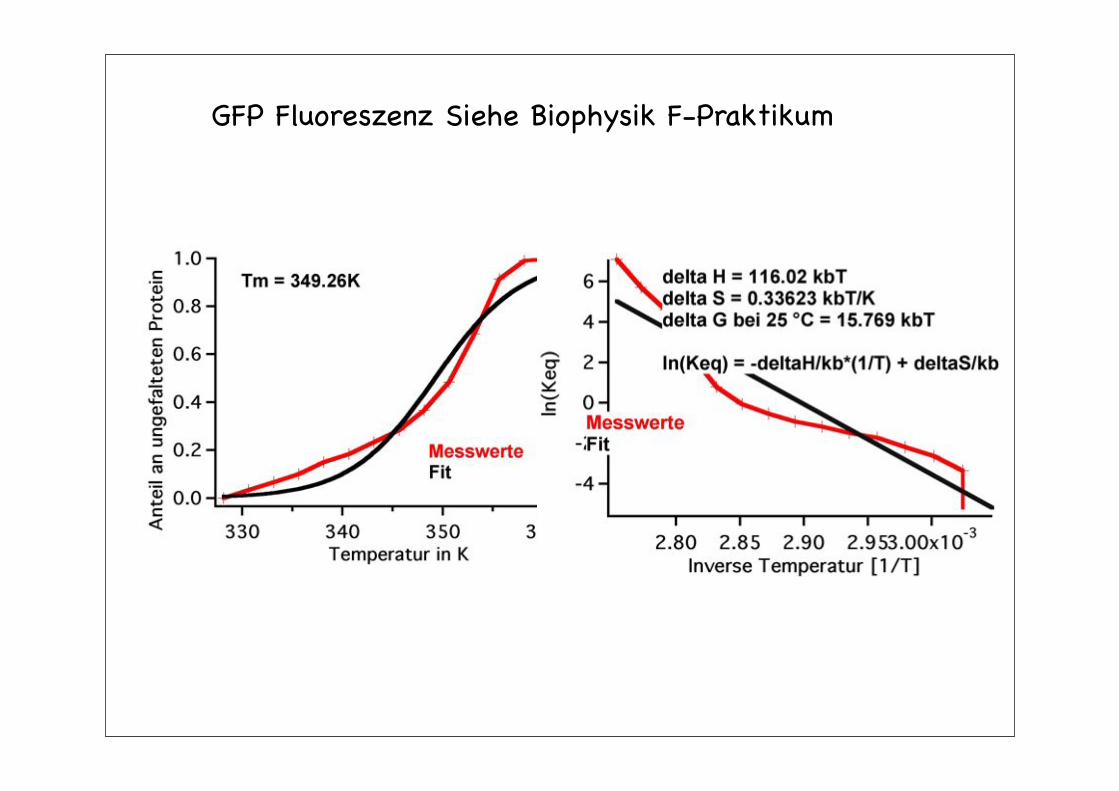
GFP Fluoreszenz Siehe Biophysik F-Praktikum

Other techniques to probe unfolding
High-resolution techniques (local):
• FTIR
• Fluorescence
• NMR
• UV absorption
Low-resolution techniques:
• SAXS
• DLS
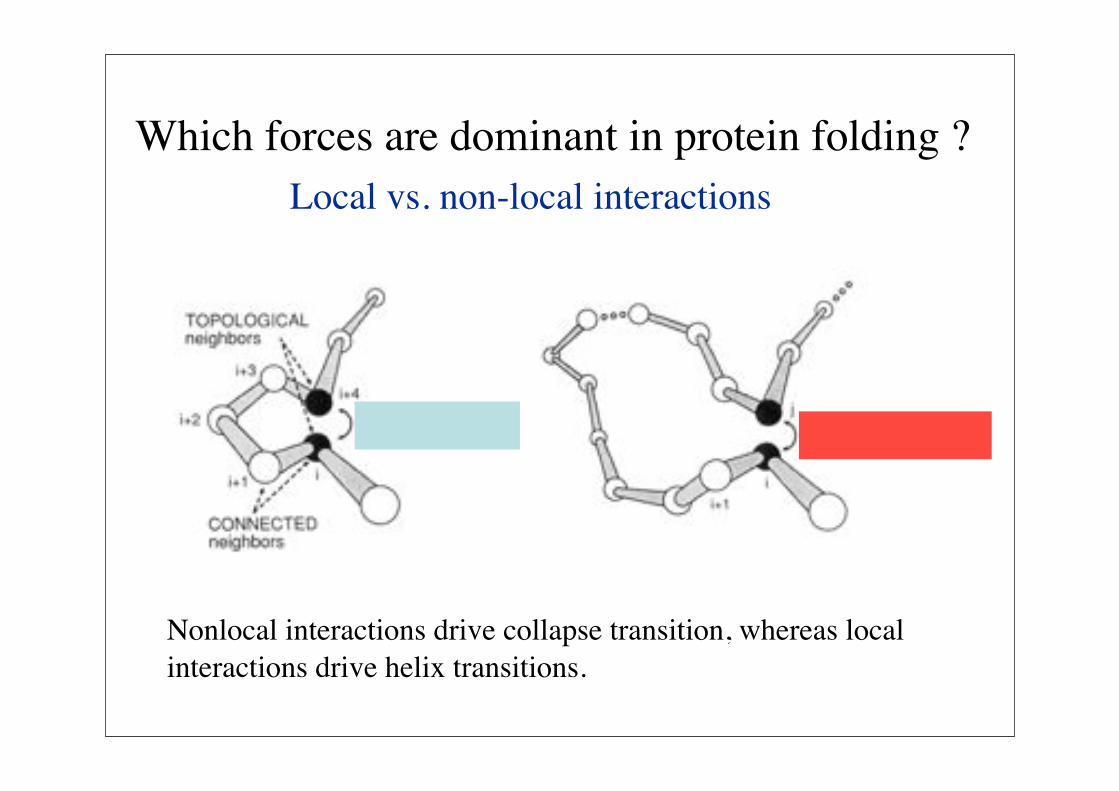
Which forces are dominant in protein folding ?Local vs. non-local interactions
Nonlocal interactions drive collapse transition, whereas local interactions drive helix transitions.


16

Early model in which protein folding was proposed to be
driven by ion-paired hydrogen bonding among side chains
(Mirsky& Pauling, 1936; Eyring & Stearn, 1939)
disproven by Jacobsen and Linderstrom-Lang

Electrostatic Contributions
• !i=(zie/4"#o)(1/r2) coulomb potential
• Sensitive to pH and ion concentrations
• pH determines total charge (pI)
• Ionic strength determines effective range of interactions
• Ion pairs contribute 1-3 kcal/mol (on surface)
• Ion pairs generally destabilizing if buried (cost up to 19 kcal/mol/ion to completely bury
• Ion pairs contribute ~5-15 kcal/mol per 150 aa’s

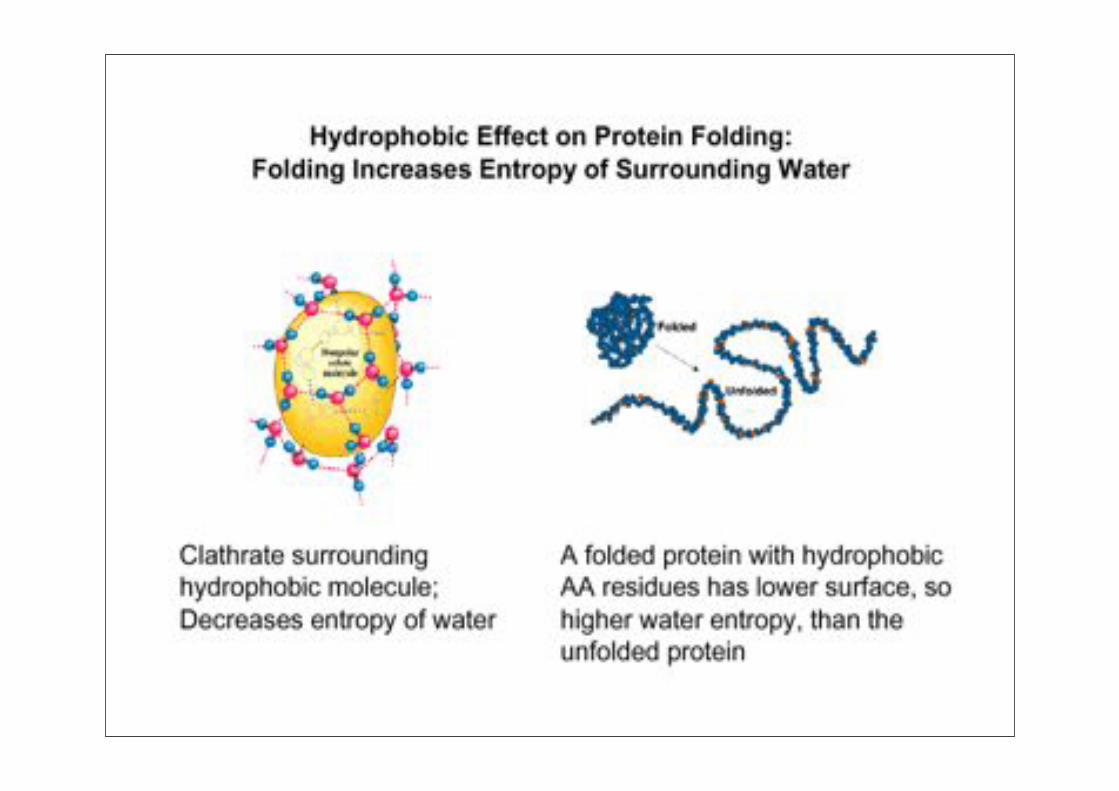
The Kauzmann Hypothesis
„hydrophobic interactions determine the thermal stability of the native state“
* non-polar solvents denature proteins* „unusual“ temperature dependence: (stability decreases at high AND low temperatures)* protein stability follows same salt dependence as lyotropic (Hofmeister) series
Key arguments
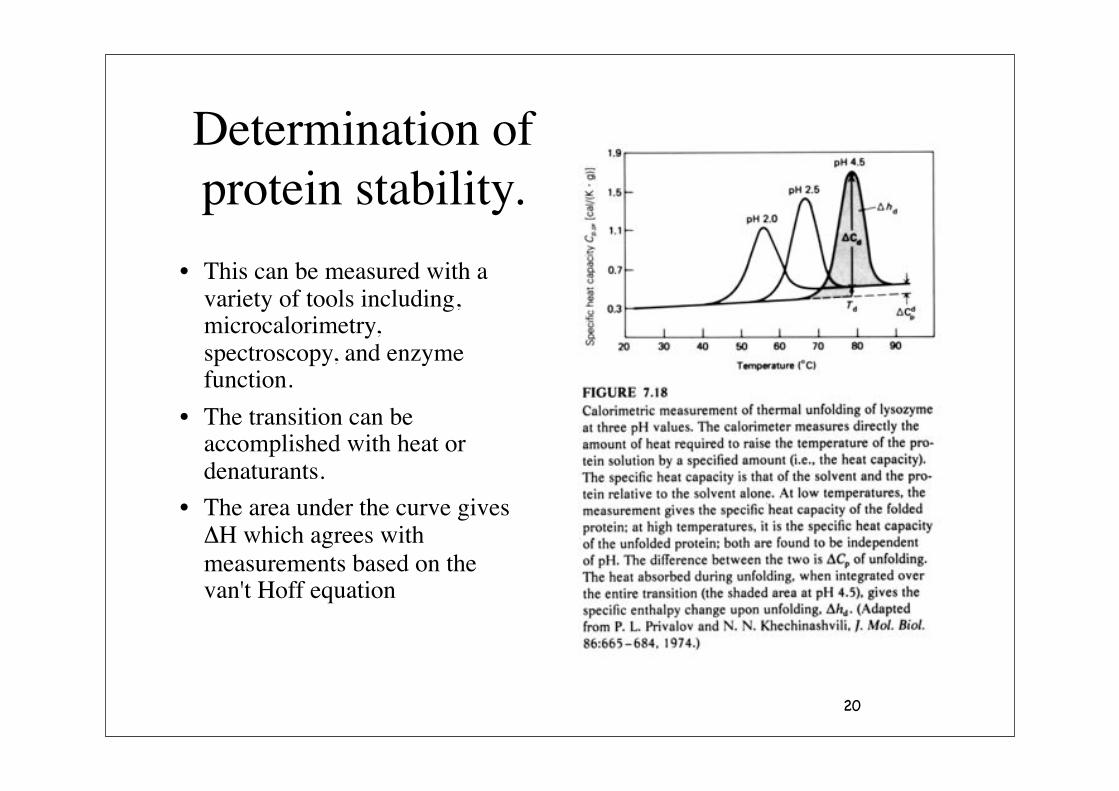
20
Determination of protein stability.
• This can be measured with a variety of tools including, microcalorimetry, spectroscopy, and enzyme function.
• The transition can be accomplished with heat or denaturants.
• The area under the curve gives $H which agrees with measurements based on the van't Hoff equation

21
Denaturants• Temperature
• pH– Change the ionization state of critical residues
• Detergents– Bind strongly to the unfolded protein
• High concentrations of water soluble organic substances– Aliphatic alcohols. These disrupt the water structure
• Ionic or polar denaturants including urea and guanidinium

22
Denaturants: The Hofmeister Series
• The ability of an ion to stabilize a protein follows the Hofmeister series
• Anions! SO4
2->H2PO4->CH3COO->Cl->Br->I'->ClO4
->SCN-
• Cations
NH4+,Cs+,K+,Na+>Li+>Mg2+>Ca2+>Ba2+
! ! ! >guanidinium>urea
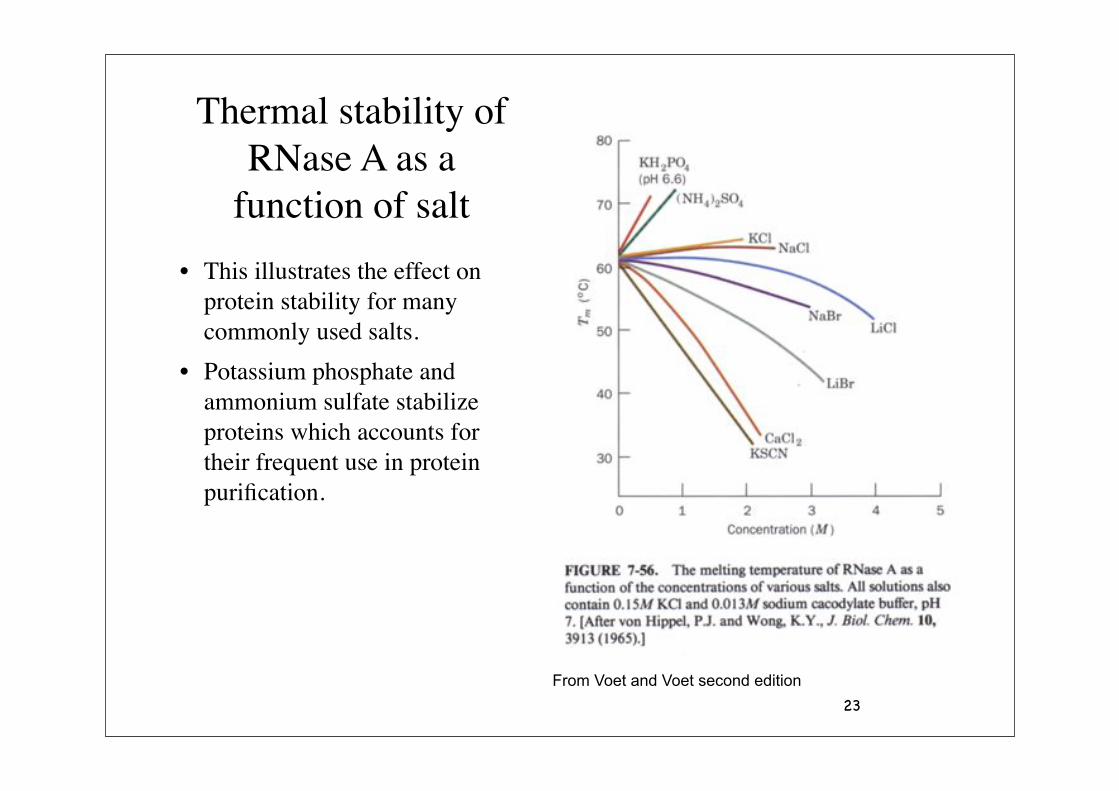
23
Thermal stability of RNase A as a
function of salt
• This illustrates the effect on protein stability for many commonly used salts.
• Potassium phosphate and ammonium sulfate stabilize proteins which accounts for their frequent use in protein purification.
From Voet and Voet second edition
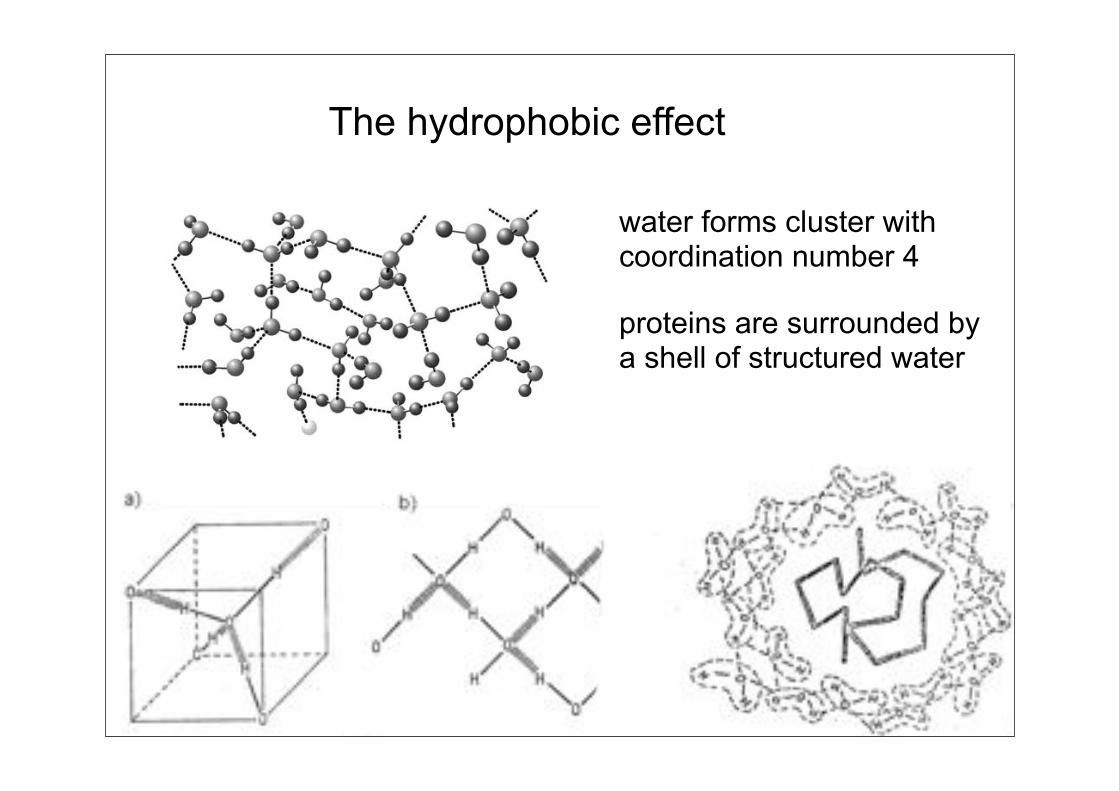
The hydrophobic effect
water forms cluster with coordination number 4
proteins are surrounded bya shell of structured water

!
µK ,W
= µK ,W
0 + RT ln xK ,W
Solubility and partition function
!
µK
0"µ
W
0 = HK
0"H
W
0( ) "T SK
0" S
W
0( )!
µW
= µK
!
ln xW
* =µK
0"µ
W
0( )RT
chemical potential,!, and partition coefficient,x of oil molecule in water (w) and oil (K)
at equilibrium:
!
"µ = µW
0#µ
K
0 = 2.44 + 0.88nC
enthalpic entropic
[kcal/Mol]
The entropic change (cost of inducing water order) dominates over the enthalpy change (gain in intermolecular interaction), which is also negative.
!
"µ = #4.2 + 0.825nC
for alcoholes
for alcanes

!"#$%&'()*%+,-"#./*0#).1&).2&"3#)4*,,#$,0&5#).+).6*,,#$.7#+.89:.;
StoffµW
0
! µK
0
in kcal/Mol
HW
0! H
K
0
in kcal/Mol
SW
0! S
K
0
in cal/Mol K
C2H
63.9 -2.5 -21
C3H
84.9 -1.7 -22
C4H
105.9 -0.8 -23
HW
0
! HK
0!"#$%&'$()*$+,-$.&%$/%&$.%-$0/%-+,1-2'3$%&'%"$(45"$.%"$67$84'$9$'):1$;$+-%&;%-.%'.%$7<-=%>$?)$.&%"%-$7%-#$'%3)@8$&"#A$&"#$.&%$0/%-+,1-2'3$%&'$%B4#1%-=%-$C-4D%*>$?&%$E)'F.%-F7))5"FG'D&%12'3$D;&":1%'$.%=$67F(45%9,5$2'.$7)""%-$&"#$/%#-<:1#5&:1>$
$!"#$'%3)@8A$.)"$/%.%2#%#A$%"$H'.%#$%&'%$G/')1=%$.%-$I)-@%55%'$J'#-4I&%$/%&$.%-
0/%-+,1-2'3$.%"$67$84'$9$'):1$;$"#)K>$LJ&"/%-3"#-29#2-$.%"$7)""%-"MSW
0
! SK
0
!µ = µW
0" µ
K
0= 2.44 + 0.88n
C%=I&-&":1N ':NO)15$.%-$6415%'"#4P)#4=%

Hydrophobic Effect• At normal temp’s the “hydrophobic effect” is entropic
water molecules form ordered structures around nonpolar
compounds
• Hydrophobic residues collapse in to exclude water
• Additional forces can then stabilize (vdw, h-bond,intrinsic properties)
• Hydrophobic effect is dependent on temperature (unstable at high AND low temp).

Thermodynamic considerations• Protein stability is composed of two components.
% % $G = $H-T$S
• There is a complex temperature dependence for $H and T$S which means that the contribution of the enthalpic and entropic terms changes with temperature.
• This temperature dependence arises from the anomalously high change in heat capacity on transferring hydrophobic compounds into water. This is the hall-mark of the hydrophobic effect and arises from the water-ordering.

Heat Capacity
• The heat capacity influences both the temperature dependence of the enthalpy and entropy
• It is proportional to the buried non-polar surface area as are all of the thermodynamic parameters.
• The large heat capacity is indicative of a well ordered water structure around non-polar molecules in water as is evident from their partial specific volumes when dissolved in water
!
Cp ="H
"T=T"S
"T
31
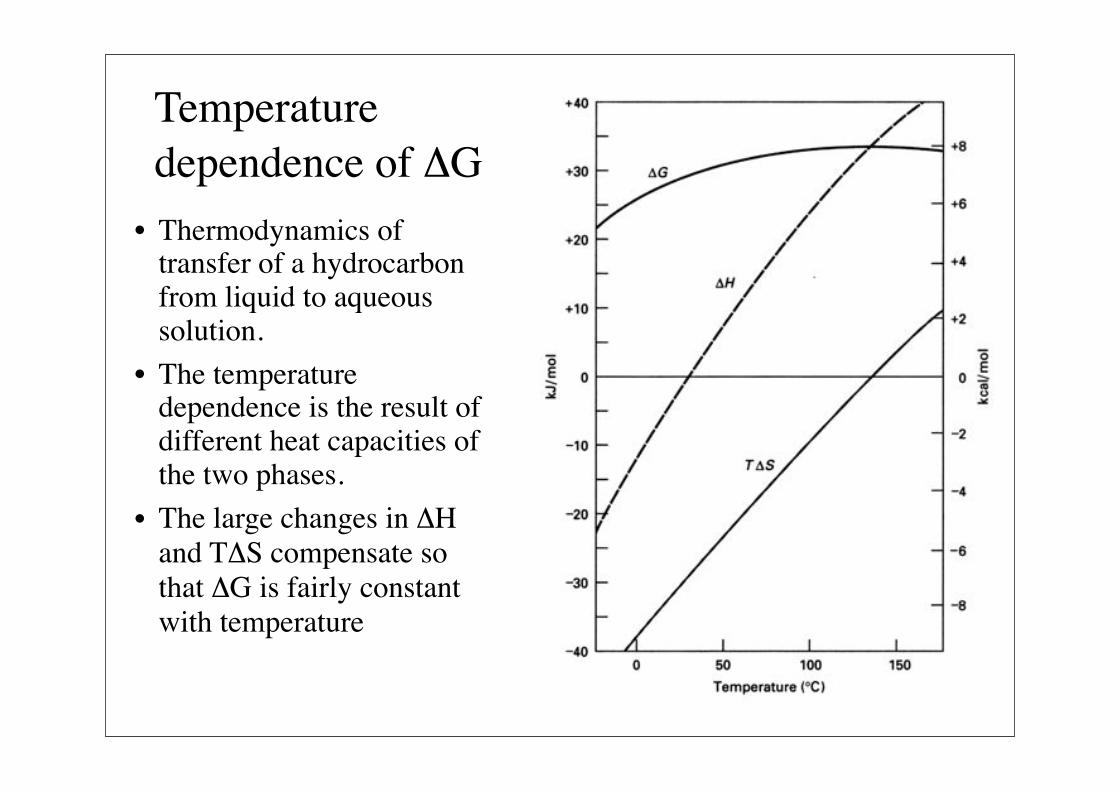
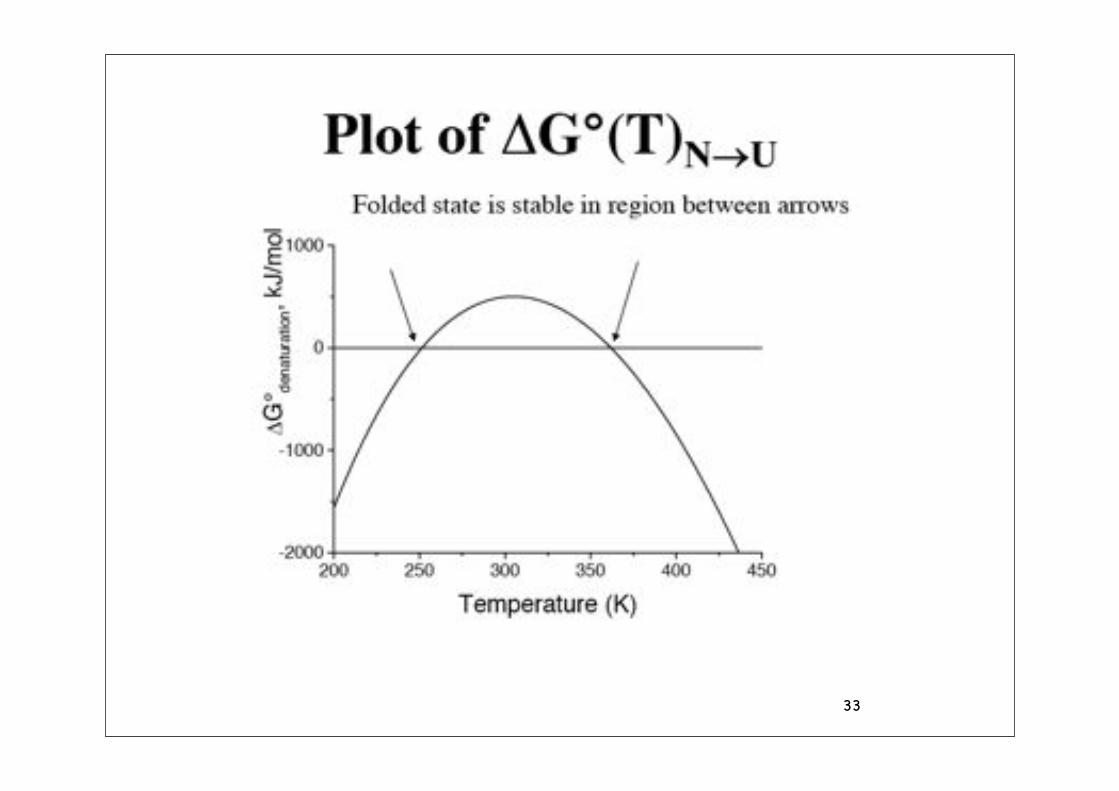
Temperature dependence of $G
• Thermodynamics of transfer of a hydrocarbon from liquid to aqueous solution.
• The temperature dependence is the result of different heat capacities of the two phases.
• The large changes in $H and T$S compensate so that $G is fairly constant with temperature
32
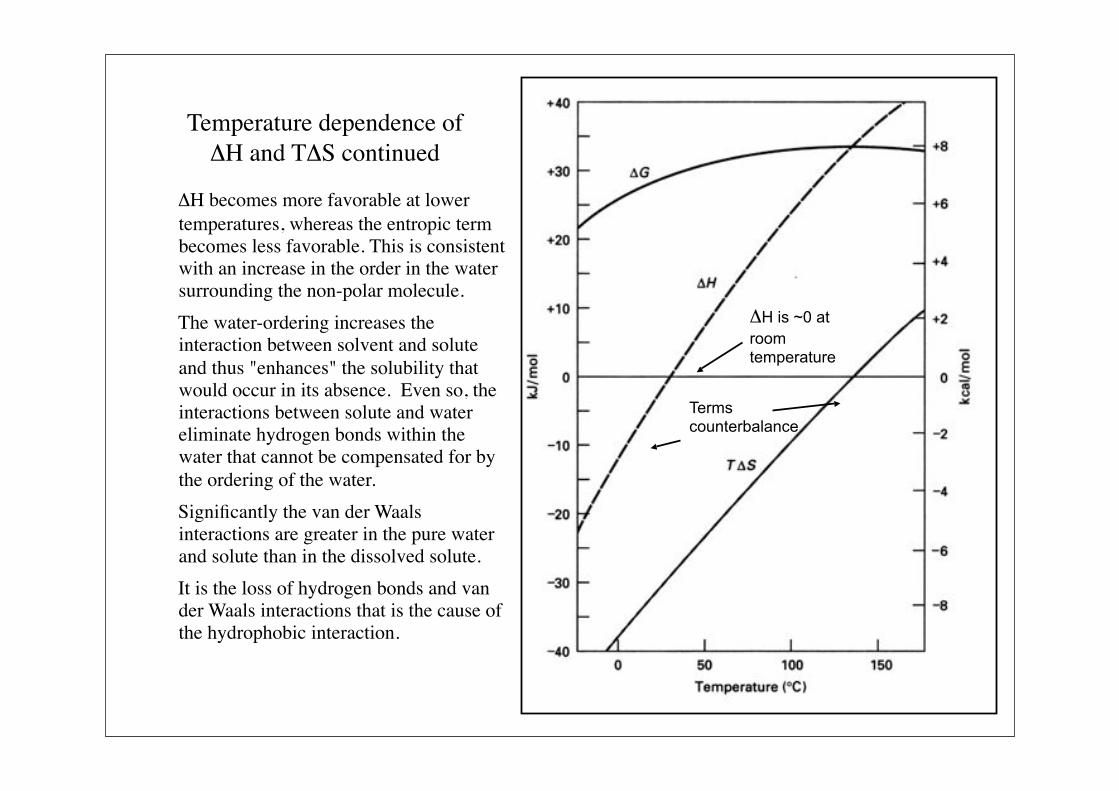
Temperature dependence of $H and T$S continued
$H becomes more favorable at lower temperatures, whereas the entropic term becomes less favorable. This is consistent with an increase in the order in the water surrounding the non-polar molecule.
The water-ordering increases the interaction between solvent and solute and thus "enhances" the solubility that would occur in its absence. Even so, the interactions between solute and water eliminate hydrogen bonds within the water that cannot be compensated for by the ordering of the water.
Significantly the van der Waals interactions are greater in the pure water and solute than in the dissolved solute.
It is the loss of hydrogen bonds and van der Waals interactions that is the cause of the hydrophobic interaction.
$H is ~0 at
room temperature
Terms counterbalance
33

34
Buried hydrophobic surface area• The buried hydrophobic surface area for a protein
correlates with the protein stability.
• Although it is difficult to predict the overall stability of a protein, it is possible to predict the worst case scenario that a mutation might produce based on changes in buried surface area.
• Occlusion of 1Å2 of hydrophobic surface area provides ~25 cal mol-1 of stability.
35
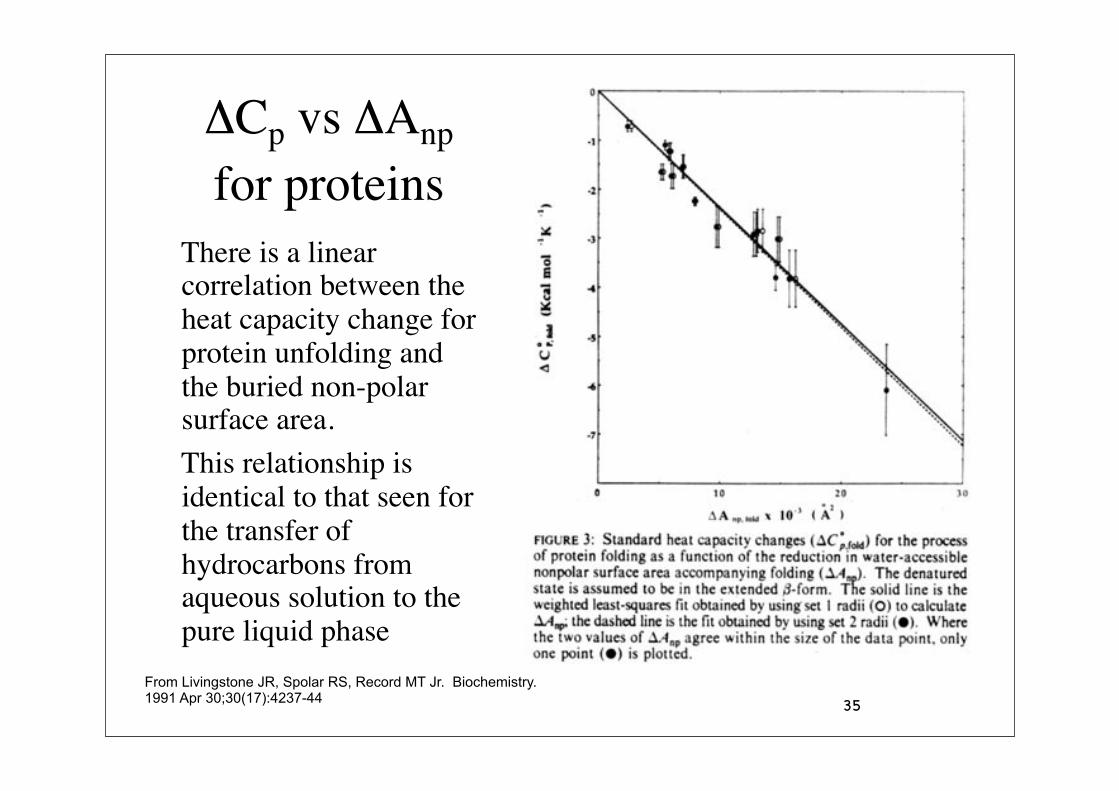
$Cp vs $Anp for proteins
There is a linear correlation between the heat capacity change for protein unfolding and the buried non-polar surface area.
This relationship is identical to that seen for the transfer of hydrocarbons from aqueous solution to the pure liquid phase
From Livingstone JR, Spolar RS, Record MT Jr. Biochemistry. 1991 Apr 30;30(17):4237-44

36
Protein Unfolding: Pressure?• 1895 Royer discovered that high hydrostatic pressure kills
bacteria.
• 1899 Hite uses pressure for milk preservation.
• 1914 Bridgman notices that egg white looks ‘cooked’ after pressure treatment.
Though it isn’t intuitive, proteins also unfold with pressure.
37
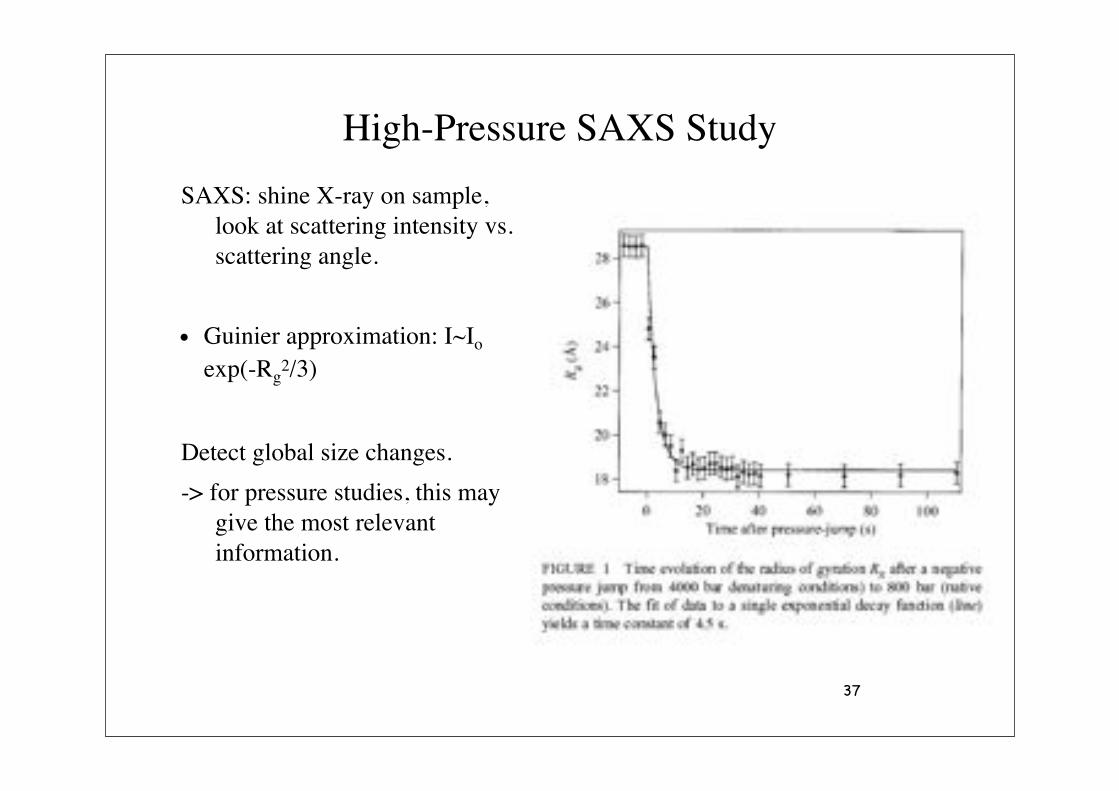
High-Pressure SAXS Study
SAXS: shine X-ray on sample, look at scattering intensity vs. scattering angle.
• Guinier approximation: I~Io exp(-Rg
2/3)
Detect global size changes.
-> for pressure studies, this may give the most relevant information.
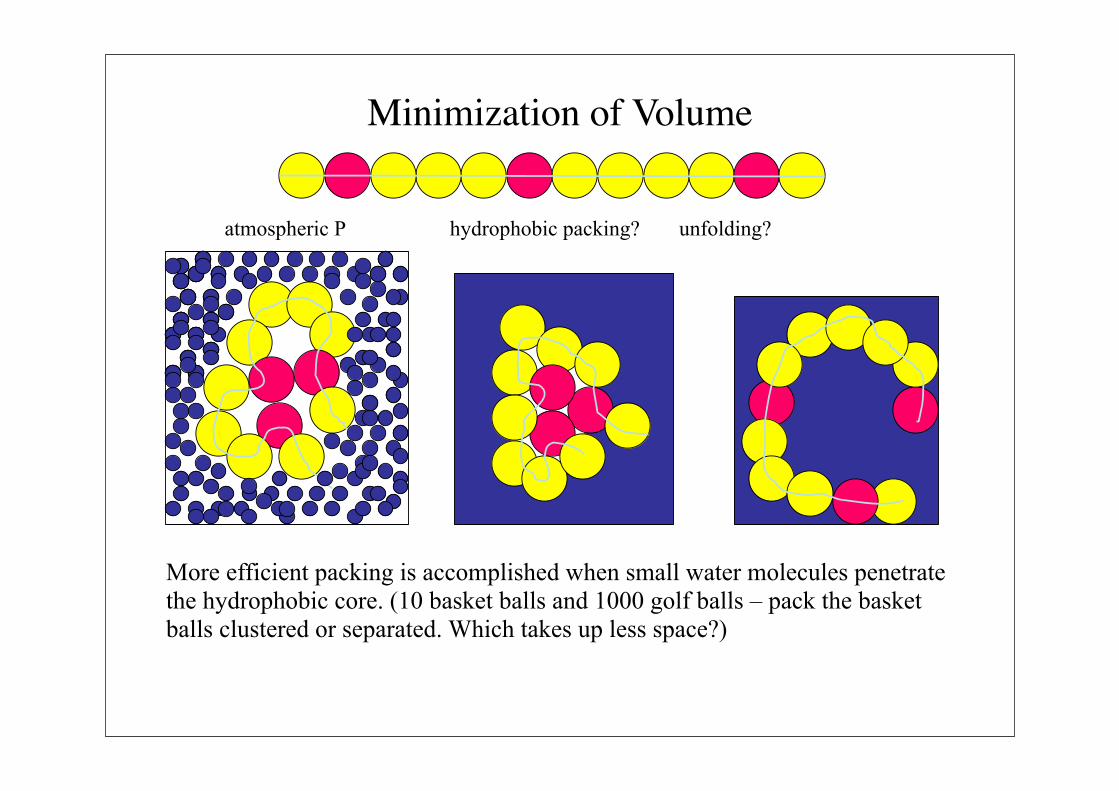
Minimization of Volume
More efficient packing is accomplished when small water molecules penetrate
the hydrophobic core. (10 basket balls and 1000 golf balls – pack the basket
balls clustered or separated. Which takes up less space?)
atmospheric P hydrophobic packing? unfolding?

Faltungsproblem
Konformation eines Proteins als Random Walk:
Gitter-Modell: Kleines Protein mit 100 Aminosäuren
=> Mögliche Konformationen: 3100" 1030
Interne Dynamik typ ns
&Zeit, um alle möglichen
Kombinationen durchzuspielen " 1021 s
Vergleiche: Alter des Universums " 1020 s !
Mother nature has no folding problem,but we do!

How many conformations are there in the Native state?
the number of sequences that have N native states decreases strongly
the HP model
molten globuleshave many configurations
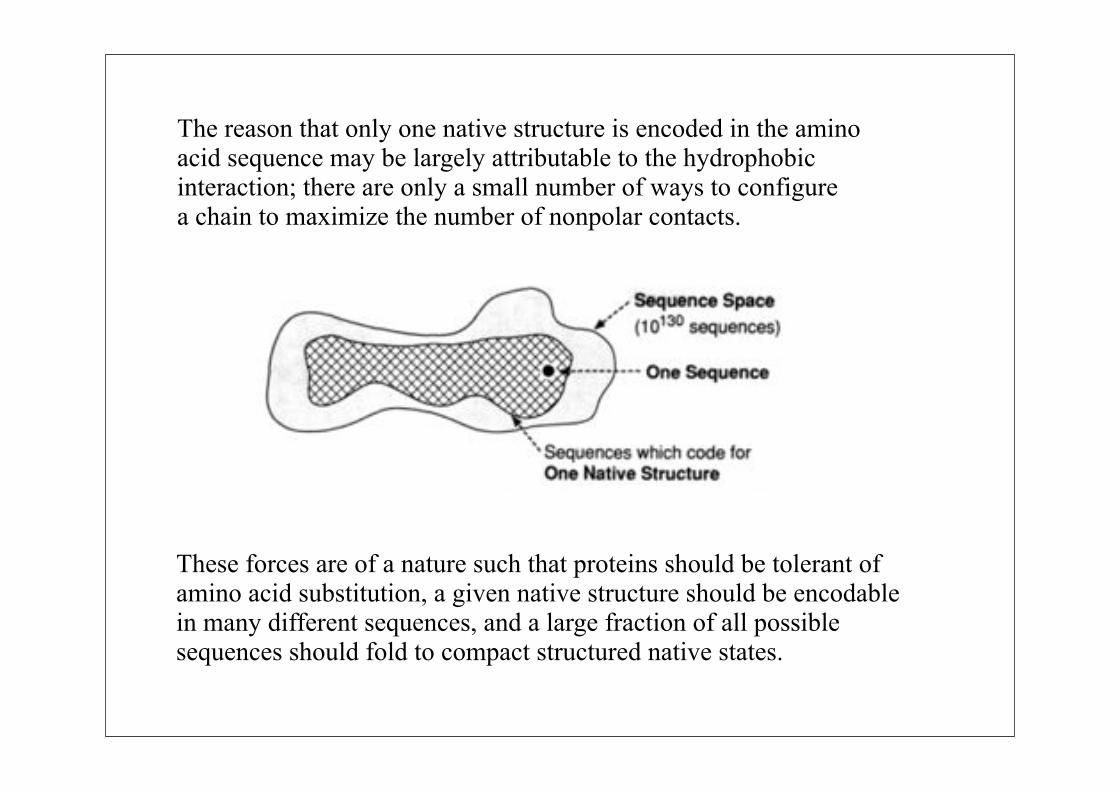
The reason that only one native structure is encoded in the aminoacid sequence may be largely attributable to the hydrophobicinteraction; there are only a small number of ways to configurea chain to maximize the number of nonpolar contacts.
These forces are of a nature such that proteins should be tolerant of amino acid substitution, a given native structure should be encodable in many different sequences, and a large fraction of all possible sequences should fold to compact structured native states.
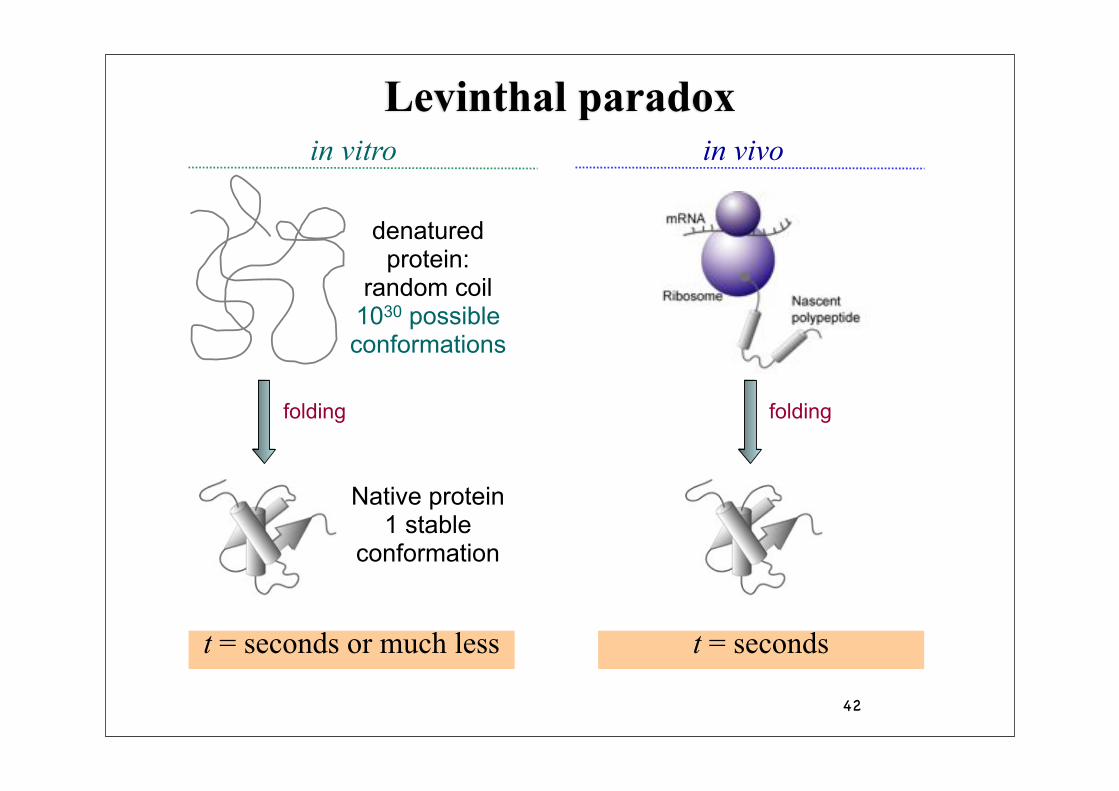
42
Levinthal paradox
folding
denaturedprotein:
random coil1030 possibleconformations
Native protein1 stable
conformation
in vitro in vivo
folding
t = secondst = seconds or much less
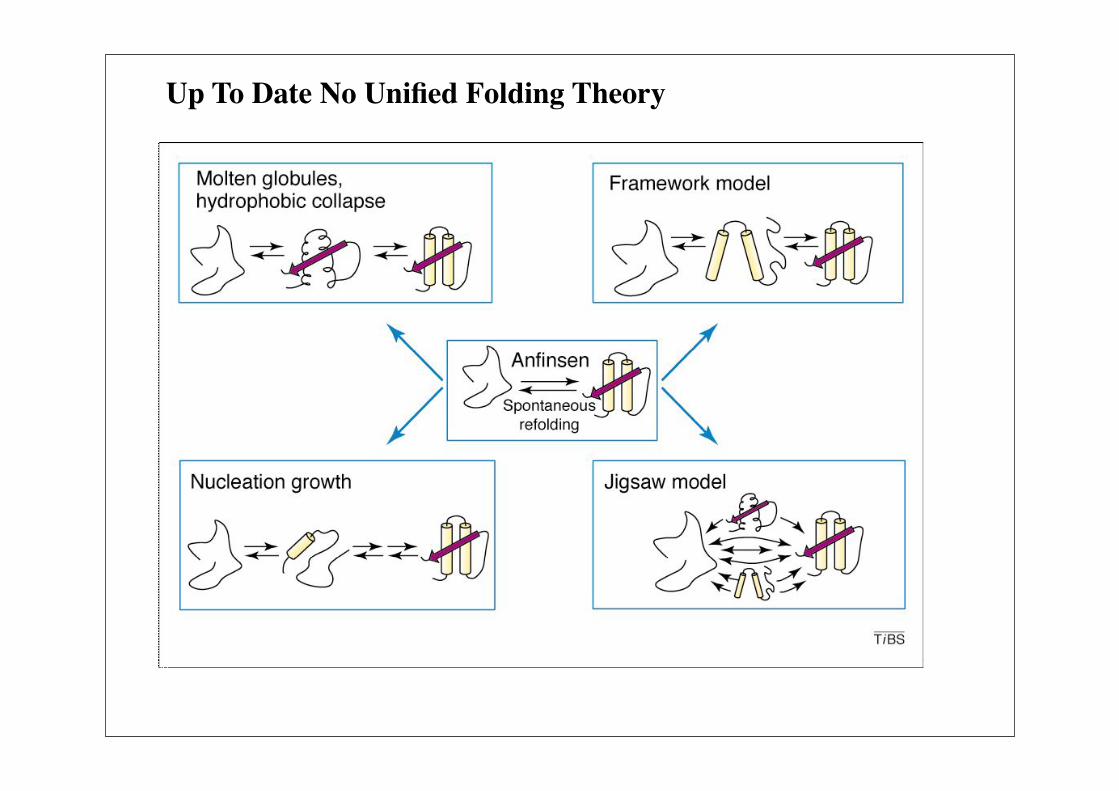
Up To Date No Unified Folding Theory