4. seleksi dan karakterisasi bakteri asam laktat … · pada proses fermentasi daging spontan, bal...
TRANSCRIPT

43
4. SELEKSI DAN KARAKTERISASI BAKTERI ASAM
LAKTAT INDIGENUS ASAL DAGING SAPI SEBAGAI
KANDIDAT PROBIOTIK SECARA IN VITRO
ABSTRAK
Sebanyak 28 isolat BAL indigenus asal daging sapi lokal Indonesia diseleksi dan dievaluasi sifat probiotiknya secara in vitro. Isolat BAL indigenus mempunyai ketahanan yang berbeda terhadap pH 2.0, 2.5, 3.2 dan 7.2 sesuai pH lambung dan usus. Hanya 10 isolat (2B4, 1B1, 1A5, 2C2, 2D1, 2B2, 1C4, 2B1, 1A32 dan 2C12) yang mampu bertahan dengan baik pada pH 2.0 dengan ketahanan hidup lebih dari 50%. Kesepuluh isolat tersebut juga menunjukkan ketahanan terhadap garam empedu 0.5% yang tinggi yaitu diatas 70%. Isolat 2B4 mempunyai ketahanan hidup terhadap garam empedu tertinggi yaitu sebanyak 90.93%. Selain itu, kesepuluh isolat mempunyai aktivitas antimikroba yang tinggi dan mampu melakukan koagregasi terhadap bakteri patogen E. coli ATCC 25922, S. Typhimurium ATCC 14028, EPEC serta S. aureus ATCC 25923 (diameter penghambatan lebih dari 6 mm, kecuali isolat 2B1 terhadap EPEC; koagregasi lebih dari 20% kecuali isolat 1A5 terhadap EPEC), serta mampu menempel pada permukaan usus sebesar 15.90-31.57%. Isolat 2C12 memiliki kemampuan penempelan tertinggi yaitu sebesar 31.57%. Berdasarkan hasil tersebut, maka disimpulkan bahwa sebanyak 10 isolat BAL indigenus asal daging sapi Indonesia memiliki sifat sebagai kandidat probiotik.
PENDAHULUAN
Pengembangan bakteri asam laktat (BAL) sebagai salah satu bahan pangan
fungsional yaitu probiotik, menjadi tren teknologi pengolahan pangan akhir-akhir
ini. BAL sering ditemukan secara alamiah dalam bahan pangan. Bakteri ini
hidup pada susu, daging segar, dan sayur-sayuran. Pada proses fermentasi daging
spontan, BAL yang berasal dari bahan mentah atau lingkungan menyebabkan
terbentuknya asam laktat dari penggunaan karbohidrat, dan menurunkan nilai pH
(5.9 – 4.6). Lactobacillus spp. merupakan genus terbesar dari kelompok BAL
(Axelsson 1993). Genus Lactobacillus bersifat Gram-positif dan tidak membentuk
spora, bersifat anaerob fakultatif, tumbuh optimum pada kisaran suhu 30-40°C
tetapi dapat tumbuh pada kisaran 5-35°C. Lactobacillus tumbuh optimum pada
pH 5.5-5.8, namun secara umum dapat tumbuh pada pH kurang dari 5.
Lactobacillus banyak terdapat pada produk makanan fermentasi seperti produk-
produk susu fermentasi (yogurt, keju, yakult) produk fermentasi daging seperti

44
sosis fermentasi, serta produk fermentasi sayuran seperti pikel dan sauerkraut.
Lactobacillus berkontribusi untuk pengawetan, ketersediaan nutrisi, dan flavor
pada produk fermentasi tersebut (Salminen & Wright 2004).
BAL mendapat perhatian besar karena banyak galur yang bermanfaat bagi
kesehatan yang disebut sebagai probiotik. Probiotik didefinisikan sebagai
mikroorganisme hidup yang apabila dikonsumsi oleh manusia atau hewan dalam
jumlah cukup, mampu memberikan manfaat kesehatan bagi inangnya (FAO/WHO
2002). FAO/WHO (2002) telah mengeluarkan panduan untuk mengevaluasi
probiotik dalam makanan. Working Group yang dibentuk oleh FAO/WHO
menetapkan secara detil panduan dan kriteria rekomendasi serta metodologi yang
digunakan untuk evaluasi probiotik, mengidentifikasi serta menentukan data-data
yang dibutuhkan untuk mengklaim kesehatan probiotik. Kriteria pertama yang
harus dipenuhi adalah bahwa isolat yang diperoleh harus diketahui
identifikasinya, baik secara fenotipik maupun genotipik, mulai dari genus sampai
spesies bahkan sampai tingkatan galur. Kriteria selanjutnya adalah karakterisasi
sifat probiotik, baik secara in vitro maupun studi hewan, kemudian dilanjutkan
dengan pengujian keamanan secara in vitro dan in vivo, serta studi fase satu di
manusia untuk produk pangan probiotik (FAO/WHO 2002).
Beberapa peneliti mengemukakan jaminan kriteria untuk bakteri
probiotik. Probiotik harus dapat bertahan melewati lambung dan usus halus,
sehingga probiotik harus toleran terhadap suasana asam dan adanya asam empedu
(Tuomola et al. 2001, Bourlioux et al. 2003, Roberfroid 2001, Sunny-Roberts &
Knoor 2008). Probiotik harus mempunyai kemampuan dalam melakukan
penempelan ke usus (Nitisinprasert et al. 2006, Tuomola et al. 2001, Bourlioux et
al. 2003), karena sangat berkaitan dengan beberapa efek kesehatan antara lain
mempersingkat durasi diare, efek imunologik dan eksklusi kompetitif dengan
mikroba patogen (Tuomola et al. 2001; Herick & Levkut 2002; Bourlioux et al.
2003).
Sampai saat ini, penelitian eksplorasi BAL yang mempunyai potensi
sebagai probiotik terus dilakukan oleh peneliti di berbagai negara, seiring dengan
peningkatan konsumsi pangan probiotik. Di Indonesia, pangan probiotik juga
semakin berkembang dan diminati masyarakat. Namun demikian, sebagian besar

45
probiotik yang digunakan masih berasal dari luar negeri. Hal ini mengakibatkan
produk pangan probiotik di Indonesia sangat tergantung pada ketersediaan dan
suplai probiotik dari luar negeri, yang akhirnya berdampak pada tingginya harga
produk tersebut. Oleh karenanya, sangat diperlukan probiotik indigenus
Indonesia agar ketersediaan probiotik sebagai bahan baku berbagai produk
pangan probiotik dapat semakin berkembang dikonsumsi oleh masyarakat
Indonesia. Sebanyak 28 isolat indigenus BAL telah diisolasi dari daging sapi
lokal dari berbagai pasar tradisional di daerah Bogor oleh Arief et al (2007).
Penelitian ini bertujuan untuk menseleksi dan mengkarakterisasi isolat
indigenus BAL asal daging lokal sebagai kandidat probiotik secara in vitro untuk
memenuhi syarat studi awal probiotik isolat bakteri baru menurut FAO/WHO
(2002). Karakteristik sifat probiotik meliputi ketahanan terhadap pH sesuai
kondisi saluran pencernaan (lambung dan usus), garam empedu, aktivitas
antimikroba terhadap bakteri patogen, koagregasi dengan bakteri patogen serta
penempelan pada permukaan usus.
BAHAN DAN METODE
Bahan
Bahan utama yang digunakan dalam penelitian ini adalah 28 buah isolat
BAL yang telah diisolasi dari daging sapi segar bangsa Peranakan Ongole di pasar
tradisional wilayah Bogor (Arief et al. 2007). Isolat diawetkan dalam bentuk liofil
dan disimpan pada suhu -30°C. Peremajaan kultur dilakukan dengan
menumbuhkannya pada media MRS broth dan kemudian disegarkan dan
disimpan pada media MRS agar sebagai kultur stok. Asal isolasi BAL tersebut
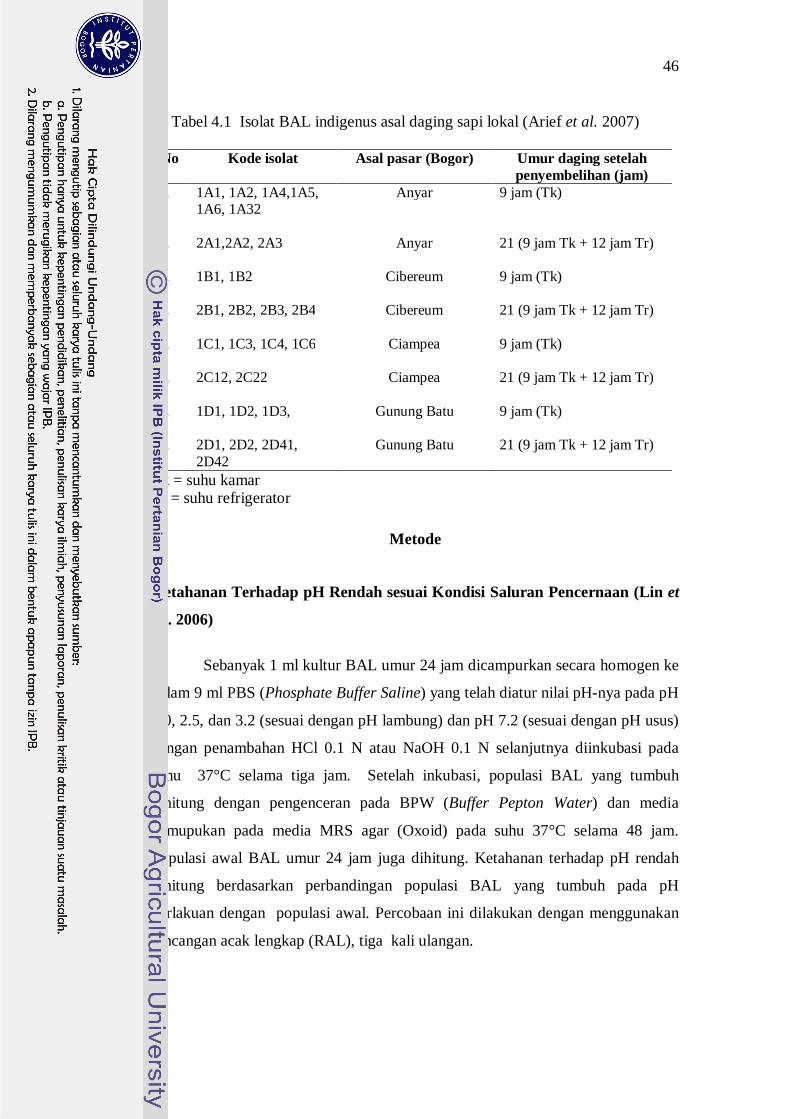
ditunjukkan pada Tabel 4.1 dan semuanya bersifat Gram positif, katalase negatif
dan non motil.
46
Tabel 4.1 Isolat BAL indigenus asal daging sapi lokal (Arief et al. 2007)
No Kode isolat Asal pasar (Bogor) Umur daging setelah penyembelihan (jam)
1. 1A1, 1A2, 1A4,1A5, 1A6, 1A32
Anyar 9 jam (Tk)
2. 2A1,2A2, 2A3
Anyar 21 (9 jam Tk + 12 jam Tr)
3. 1B1, 1B2 Cibereum
9 jam (Tk)
4. 2B1, 2B2, 2B3, 2B4
Cibereum 21 (9 jam Tk + 12 jam Tr)
5. 1C1, 1C3, 1C4, 1C6
Ciampea 9 jam (Tk)
6. 2C12, 2C22 Ciampea
21 (9 jam Tk + 12 jam Tr)
7. 1D1, 1D2, 1D3, Gunung Batu
9 jam (Tk)
8. 2D1, 2D2, 2D41, 2D42
Gunung Batu
21 (9 jam Tk + 12 jam Tr)
Tk = suhu kamar Tr = suhu refrigerator
Metode
Ketahanan Terhadap pH Rendah sesuai Kondisi Saluran Pencernaan (Lin et
al . 2006)
Sebanyak 1 ml kultur BAL umur 24 jam dicampurkan secara homogen ke
dalam 9 ml PBS (Phosphate Buffer Saline) yang telah diatur nilai pH-nya pada pH
2.0, 2.5, dan 3.2 (sesuai dengan pH lambung) dan pH 7.2 (sesuai dengan pH usus)
dengan penambahan HCl 0.1 N atau NaOH 0.1 N selanjutnya diinkubasi pada
suhu 37°C selama tiga jam. Setelah inkubasi, populasi BAL yang tumbuh
dihitung dengan pengenceran pada BPW (Buffer Pepton Water) dan media
pemupukan pada media MRS agar (Oxoid) pada suhu 37°C selama 48 jam.
Populasi awal BAL umur 24 jam juga dihitung. Ketahanan terhadap pH rendah
dihitung berdasarkan perbandingan populasi BAL yang tumbuh pada pH
perlakuan dengan populasi awal. Percobaan ini dilakukan dengan menggunakan
rancangan acak lengkap (RAL), tiga kali ulangan.

47
Ketahanan terhadap Garam Empedu (Lin et al. 2006)
Uji ketahanan terhadap garam empedu dilakukan dengan menumbuhkan
BAL yang tahan hidup pada pH 2.0 selama tiga jam inkubasi (ketahanan hidup ≥
50%) pada media MRS broth yang mengandung garam empedu 0.5%. Sebanyak
satu ml kultur BAL umur 24 jam dimasukkan ke dalam sembilan ml MRS broth
yang mengandung 0.5% garam empedu (Bile salt, Pronadisa) lalu diinkubasikan
pada suhu 37°C selama enam jam sesuai dengan lamanya waktu transit makanan
di usus halus sebelum ke usus besar (Bourlioux et al. 2003). Populasi awal BAL
yang berumur 24 jam sebelum diinokulasikan ke media MRS yang mengandung
garam empedu dihitung. Jumlah BAL dihitung pada media MRSA dengan metode
tuang dengan inkubasi suhu 37°C selama 48 jam. Nilai ketahanan hidup
ditunjukkan dengan persentase populasi yang tumbuh pada media garam empedu
0.5% dibandingkan dengan populasi awal. Percobaan ini dilakukan dengan RAL,
tiga kali ulangan.
Uji Aktivitas Antimikroba BAL (Savadogo et al. 2004)
Isolat indigenus BAL diinokulasikan ke dalam MRS broth dan
diinkubasikan pada suhu 37°C selama 20 jam. Supernatan bebas sel dipanen
melalui sentrifugasi 10.000 rpm pada suhu 4 °C selama 20 menit, lalu disaring
dengan menggunakan 0.22 µm membran filter (Sartorius). Selanjutnya
supernatan bebas sel yang merupakan substrat antimikroba tersebut siap untuk
diuji aktivitas antimikrobanya dengan menggunakan metode difusi agar.
Bakteri uji yang digunakan adalah beberapa bakteri patogen yaitu
Escherichia coli ATCC 25922, Salmonella Typhimurium ATCC 14028
Staphylococcus aureus ATCC 25923 serta Escherichia coli enteropatogen
(EPEC). Bakteri uji dibiakkan pada media Nutrien Agar (Difco) selama 24 jam
pada suhu 37°C. Setelah itu diambil sebanyak satu ose kultur kerja tersebut lalu
dibiakkan ke dalam tabung berisi media Nutrien Broth (Difco). Setelah 24 jam
inkubasi, kultur bakteri uji diambil dua ose untuk diinokulasikan ke larutan
pengencer NaCl 0.85% lalu disetarakan kekeruhannya dengan standar Mc.
Farland no 0.5, yang memiliki kesamaan dengan jumlah populasi bakteri sebesar

48
8x108 cfu/ml. Suspensi bakteri uji yang terbentuk kemudian diencerkan dengan
NaCl fisiologis 0.85% sampai diperoleh konsentrasi 106 cfu/ml.
Suspensi bakteri uji yang telah diencerkan kemudian diambil sebanyak
satu ml dengan pipet steril kemudian dituangkan ke dalam cawan Petri steril,
setelah itu dilanjutkan dengan menuangkan media Mueller Hinton Agar (MHA)
bersuhu + 50°C. Setelah mengeras dibuat lubang sumur berdiameter lima mm
dengan menggunakan ujung pipet Pasteur steril. Sebanyak 50 µl supernatan bebas
sel dituangkan ke dalam setiap lubang sumur. Seluruh cawan diinkubasi pada
suhu 37°C selama 24 jam. Zona hambat yang terbentuk di sekitar sumur diamati
dan diukur diameternya dengan memakai jangka sorong. Percobaan ini dilakukan
sebanyak tiga kali ulangan secara duplo dengan menggunakan RAL.
Pengukuran pH dilakukan dengan menggunakan pH meter Hanna.
Terlebih dahulu pH meter dikalibrasi dengan larutan standar (ber-pH 4 dan 7),
kemudian elektrodanya dimasukkan ke dalam sampel 10 ml supernatan bebas sel
dan diamati nilai pH-nya. Penentuan total asam laktat dilakukan dengan metode
titrasi. Oleh karena pada pengujian awal diketahui bahwa semua isolat merupakan
bakteri homofermentatif, maka total asam tertitrasi yang diperoleh dinyatakan
sebagai asam laktat dengan adanya faktor koreksi asam laktat. Supernatan bebas
sel sebanyak 10 ml dimasukkan ke dalam labu Erlenmeyer, kemudian
ditambahkan tiga tetes larutan indikator phenophtalein 1%. Selanjutnya
supernatan dititrasi dengan larutan NaOH 0.1 N sampai terbentuk warna merah
muda. Total asam tertitrasi dihitung sebagai persen asam laktat dengan rumus:
NaOH (ml) x N.NaOH x BM x FK % asam laktat = x 100% sampel (ml)
Keterangan : N = normalitas BM = berat molekul asam laktat (90), 1 ml NaOH 0.1 N = 0.009 g asam laktat FK = faktor pengencer

49
Uji Koagregasi BAL dengan Bakteri Patogen
Uji koagregasi BAL dengan bakteri patogen dilakukan sesuai metode El-
Naggar (2004). Isolat BAL ditumbuhkan pada suhu 37 °C selama 24 jam di MRS
broth, sedangkan bakteri patogen ditumbuhkan pada media NB diinkubasikan
pada suhu 37°C selama 24 jam. Suspensi isolat BAL dan bakteri uji diukur OD-
nya pada panjang gelombang 660 nm. Setiap volume 0.5 ml dari setiap suspensi
bakteri dicampur dengan vortex termasuk kontrol dan OD diukur setelah 4 jam.
Tabung kontrol berisi 1 ml suspensi setiap bakteri. Percobaan dilakukan dengan
RAL, tiga kali ulangan secara duplo. Persentase koagregasi dikalkulasikan dengan
menggunakan persamaan sebagai berikut :
(A + B)/2 - C x 100 (A + B)/2 Keterangan :
A = OD (660 nm) dari tabung kontrol isolat BAL yang diukur pada waktu
inkubasi 4 jam
B = OD (660 nm) dari tabung kontrol bakteri patogen yang diukur pada
waktu inkubasi 4 jam
C = OD (660 nm) yang diukur dari suspensi campuran bakteri patogen
dengan isolat BAL pada waktu inkubasi 4 jam
Uji Penempelan BAL pada Permukaan Usus secara in vitro
Pengujian sifat penempelan BAL pada permukaan usus dilakukan sesuai
dengan metode Kos et al. (2003) dan Nitisinprasert et al.,(2006) yang
dimodifikasi. Sampel permukaan usus bagian ileum yang diambil dari tikus yang
berusia enam minggu dipotong dengan ukuran 1 cm x 1 cm. Terlebih dahulu,
usus dicuci dengan PBS sebanyak tiga kali, lalu dilakukan perendaman dengan
PBS selama 30 menit pada suhu refrigerator untuk menghilangkan mukosa usus.
Pengujian penempelan dilakukan dengan menginkubasikan permukaan usus yang
telah dihilangkan mukosanya pada suspensi BAL (populasi 108 cfu/ml) pada
larutan PBS pada suhu 37°C selama 30 menit, kemudian dilakukan pencucian
dengan PBS kembali sebanyak tiga kali. Selain itu dipersiapkan kontrol yaitu usus

50
yang tidak diinkubasi dengan suspensi BAL. Setelah itu, dilakukan penghitungan
populasi BAL yang menempel pada permukaan usus dengan metode tuang
menggunakan media MRS agar ditambah CaCO3 0.5%. Sifat penempelan BAL
pada permukaan usus dihitung dengan cara menghitung selisih populasi BAL
yang menempel pada permukaan usus pada perlakuan inkubasi dengan suspensi
BAL indigenus dibandingkan populasi BAL pada kontrol. Percobaan ini
dilakukan dengan RAL, tiga kali ulangan.
Analisis Data
Data yang diperoleh dianalisis dengan menggunakan Analysis of Variance
(ANOVA) (Steel & Torrie 1995). Data populasi BAL terlebih dahulu
ditransformasikan ke nilai log10. Apabila terjadi perbedaan antar perlakuan, maka
dilanjutkan dengan uji Tukey menggunakan bantuan program komputer Minitab
14.
HASIL DAN PEMBAHASAN
Ketahanan BAL terhadap pH Rendah sesuai Kondisi Saluran Pencernaan Untuk dapat bertahan dalam saluran pencernaan isolat probiotik harus
dapat melewati kondisi ekstrim keasaman yang tinggi di lambung serta mampu
bertahan pada kondisi garam empedu di saluran pencernaan. Ketahanan terhadap
tingkat keasaman yang tinggi merupakan sifat yang pertama yang harus dipenuhi
sebagai probiotik pada saat akan melakukan seleksi isolat probiotik (Tuomola et
al. 2001). Isolat indigenus BAL sebanyak 28 isolat diseleksi ketahanan hidupnya
pada kondisi pH 2.0, 2.5, 3.2 dan 7.2. Nilai pH 2.0 pada larutan PBS mewakili
nilai pH lambung saat lambung kosong, tidak terisi makanan, sedangkan nilai pH
2.5 dan 3.2 dipilih mewakili kondisi pH lambung saat lambung terisi makanan.
Nilai pH 7.2 ditentukan dengan berdasarkan pertimbangan kondisi pH di usus
halus. Populasi awal isolat indigenus BAL serta pertumbuhannya pada kondisi pH
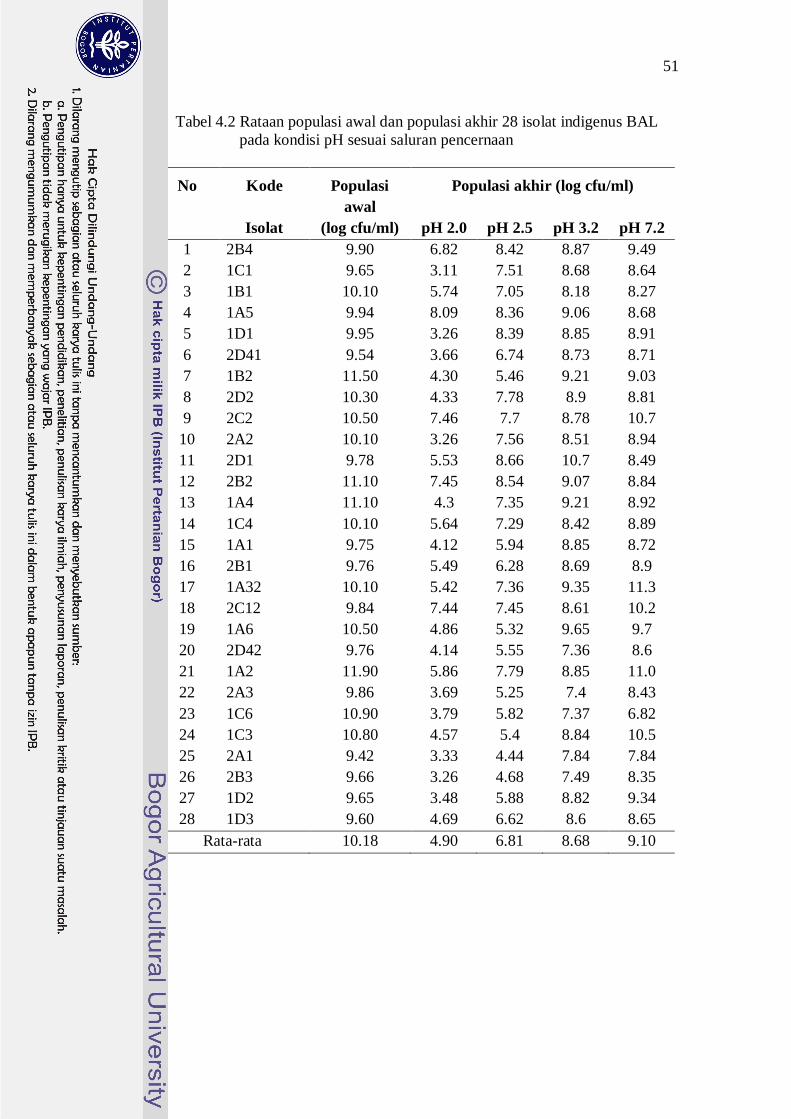
yang berbeda dapat dilihat pada Tabel 4.2.
51
Tabel 4.2 Rataan populasi awal dan populasi akhir 28 isolat indigenus BAL pada kondisi pH sesuai saluran pencernaan
No Kode Populasi Populasi akhir (log cfu/ml)
Isolat awal
(log cfu/ml) pH 2.0 pH 2.5 pH 3.2 pH 7.2 1 2B4 9.90 6.82 8.42 8.87 9.49 2 1C1 9.65 3.11 7.51 8.68 8.64 3 1B1 10.10 5.74 7.05 8.18 8.27 4 1A5 9.94 8.09 8.36 9.06 8.68 5 1D1 9.95 3.26 8.39 8.85 8.91 6 2D41 9.54 3.66 6.74 8.73 8.71 7 1B2 11.50 4.30 5.46 9.21 9.03 8 2D2 10.30 4.33 7.78 8.9 8.81 9 2C2 10.50 7.46 7.7 8.78 10.7
10 2A2 10.10 3.26 7.56 8.51 8.94 11 2D1 9.78 5.53 8.66 10.7 8.49 12 2B2 11.10 7.45 8.54 9.07 8.84 13 1A4 11.10 4.3 7.35 9.21 8.92 14 1C4 10.10 5.64 7.29 8.42 8.89 15 1A1 9.75 4.12 5.94 8.85 8.72 16 2B1 9.76 5.49 6.28 8.69 8.9 17 1A32 10.10 5.42 7.36 9.35 11.3 18 2C12 9.84 7.44 7.45 8.61 10.2 19 1A6 10.50 4.86 5.32 9.65 9.7 20 2D42 9.76 4.14 5.55 7.36 8.6 21 1A2 11.90 5.86 7.79 8.85 11.0 22 2A3 9.86 3.69 5.25 7.4 8.43 23 1C6 10.90 3.79 5.82 7.37 6.82 24 1C3 10.80 4.57 5.4 8.84 10.5 25 2A1 9.42 3.33 4.44 7.84 7.84 26 2B3 9.66 3.26 4.68 7.49 8.35 27 1D2 9.65 3.48 5.88 8.82 9.34 28 1D3 9.60 4.69 6.62 8.6 8.65
Rata-rata 10.18 4.90 6.81 8.68 9.10
52
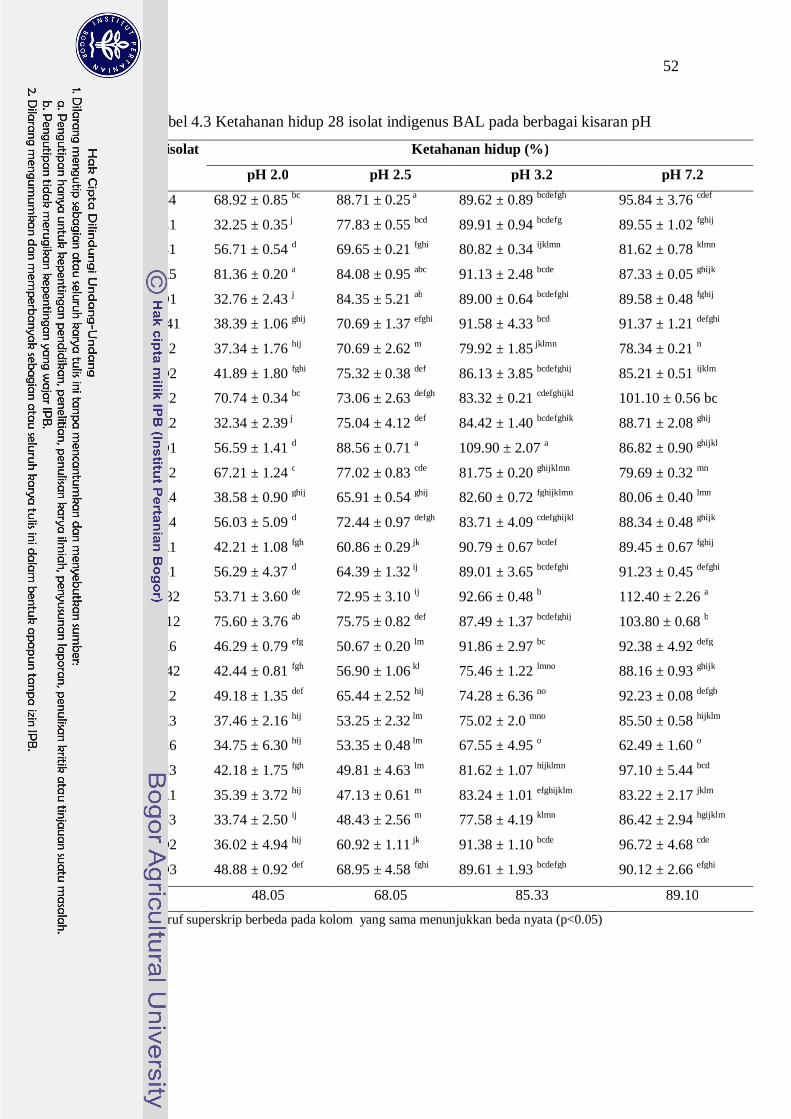
Tabel 4.3 Ketahanan hidup 28 isolat indigenus BAL pada berbagai kisaran pH
No Kode isolat Ketahanan hidup (%)
pH 2.0 pH 2.5 pH 3.2 pH 7.2
1. 2B4 68.92 ± 0.85 bc 88.71 ± 0.25 a 89.62 ± 0.89 bcdefgh 95.84 ± 3.76 cdef
2. 1C1 32.25 ± 0.35 j 77.83 ± 0.55 bcd 89.91 ± 0.94 bcdefg 89.55 ± 1.02 fghij
3. 1B1 56.71 ± 0.54 d 69.65 ± 0.21 fghi 80.82 ± 0.34 ijklmn 81.62 ± 0.78 klmn
4. 1A5 81.36 ± 0.20 a 84.08 ± 0.95 abc 91.13 ± 2.48 bcde 87.33 ± 0.05 ghijk
5. 1D1 32.76 ± 2.43 j 84.35 ± 5.21 ab 89.00 ± 0.64 bcdefghi 89.58 ± 0.48 fghij
6. 2D41 38.39 ± 1.06 ghij 70.69 ± 1.37 efghi 91.58 ± 4.33 bcd 91.37 ± 1.21 defghi
7. 1B2 37.34 ± 1.76 hij 70.69 ± 2.62 m 79.92 ± 1.85 jklmn 78.34 ± 0.21 n
8. 2D2 41.89 ± 1.80 fghi 75.32 ± 0.38 def 86.13 ± 3.85 bcdefghij 85.21 ± 0.51 ijklm
9. 2C2 70.74 ± 0.34 bc 73.06 ± 2.63 defgh 83.32 ± 0.21 cdefghijkl 101.10 ± 0.56 bc
10. 2A2 32.34 ± 2.39 j 75.04 ± 4.12 def 84.42 ± 1.40 bcdefghik 88.71 ± 2.08 ghij
11. 2D1 56.59 ± 1.41 d 88.56 ± 0.71 a 109.90 ± 2.07 a 86.82 ± 0.90 ghijkl
12. 2B2 67.21 ± 1.24 c 77.02 ± 0.83 cde 81.75 ± 0.20 ghijklmn 79.69 ± 0.32 mn
13. 1A4 38.58 ± 0.90 ghij 65.91 ± 0.54 ghij 82.60 ± 0.72 fghijklmn 80.06 ± 0.40 lmn
14. 1C4 56.03 ± 5.09 d 72.44 ± 0.97 defgh 83.71 ± 4.09 cdefghijkl 88.34 ± 0.48 ghijk
15. 1A1 42.21 ± 1.08 fgh 60.86 ± 0.29 jk 90.79 ± 0.67 bcdef 89.45 ± 0.67 fghij
16. 2B1 56.29 ± 4.37 d 64.39 ± 1.32 ij 89.01 ± 3.65 bcdefghi 91.23 ± 0.45 defghi
17. 1A32 53.71 ± 3.60 de 72.95 ± 3.10 ij 92.66 ± 0.48 b 112.40 ± 2.26 a
18. 2C12 75.60 ± 3.76 ab 75.75 ± 0.82 def 87.49 ± 1.37 bcdefghij 103.80 ± 0.68 b
19. 1A6 46.29 ± 0.79 efg 50.67 ± 0.20 lm 91.86 ± 2.97 bc 92.38 ± 4.92 defg
20. 2D42 42.44 ± 0.81 fgh 56.90 ± 1.06 kl 75.46 ± 1.22 lmno 88.16 ± 0.93 ghijk
21. 1A2 49.18 ± 1.35 def 65.44 ± 2.52 hij 74.28 ± 6.36 no 92.23 ± 0.08 defgh
22. 2A3 37.46 ± 2.16 hij 53.25 ± 2.32 lm 75.02 ± 2.0 mno 85.50 ± 0.58 hijklm
23. 1C6 34.75 ± 6.30 hij 53.35 ± 0.48 lm 67.55 ± 4.95 o 62.49 ± 1.60 o
24. 1C3 42.18 ± 1.75 fgh 49.81 ± 4.63 lm 81.62 ± 1.07 hijklmn 97.10 ± 5.44 bcd
25. 2A1 35.39 ± 3.72 hij 47.13 ± 0.61 m 83.24 ± 1.01 efghijklm 83.22 ± 2.17 jklm
26. 2B3 33.74 ± 2.50 ij 48.43 ± 2.56 m 77.58 ± 4.19 klmn 86.42 ± 2.94 hgijklm
27. 1D2 36.02 ± 4.94 hij 60.92 ± 1.11 jk 91.38 ± 1.10 bcde 96.72 ± 4.68 cde
28. 1D3 48.88 ± 0.92 def 68.95 ± 4.58 fghi 89.61 ± 1.93 bcdefgh 90.12 ± 2.66 efghi
Rata-rata 48.05 68.05 85.33 89.10 Huruf superskrip berbeda pada kolom yang sama menunjukkan beda nyata (p<0.05)

53
Semua isolat indigenus BAL mempunyai ketahanan hidup yang berbeda
nyata (p<0.05) terhadap setiap kondisi pH 2.0, 2.5, 3.2, dan 7.2 (Tabel 4.2). Nilai
persentase ketahanan hidup sangat bervariasi pada semua kondisi pH. Semua
isolat indigenus BAL mampu bertahan hidup lebih baik pada kondisi pH yang
lebih tinggi. Hal ini ditunjukkan dengan nilai rataan persentase ketahanan hidup
semua isolat BAL pada kondisi pH 7.2 (sesuai kondisi pH di usus halus), adalah
paling tinggi (89.10%), diikuti dengan ketahanan hidup pada pH 3.2 (85.33%),
selanjutnya pada pH 2.5 (68.05%) dan ketahanan hidup terendah adalah pada pH
2.0 (48.05%).
Populasi awal semua isolat BAL yaitu populasi setelah ditumbuhkan pada
media MRS broth pada suhu 37°C selama 24 jam, yang juga digunakan pada
pengujian ketahanan pada pH rendah, berkisar pada 9 – 11 log cfu/ml, dengan
rataan populasi 10.18 log cfu/ml. Pada pH rendah, yaitu pH 2.0, 2.5, dan 3.2,
isolat BAL mengalami penurunan populasi, dengan rataan populasi yang tahan
pada pH 2.0 adalah 4.90 log cfu/ml; pada pH 3.2 sebesar 6.81 log cfu/ml, rataan
populasi pada pH 3.2 sebesar 8.68 log cfu/ml serta populasi yang mampu bertahan
hidup paling tinggi adalah pada pH 7.2 sebesar 9.10 log cfu/ml. Pada kondisi pH
2.0 selama tiga jam, ketahanan hidup isolat indigenus BAL berkisar dari 32.34% -
75.60% dengan rataan 48.05%. Sebanyak 10 isolat mempunyai nilai persentase
ketahanan hidup pada kondisi pH 2.0 lebih besar dari 50%. Kesepuluh isolat
tersebut juga mampu mempertahankan populasinya minimal sebesar 105 cfu/ml.
Lin et al (2006) menyatakan bahwa ketahanan hidup BAL ≥ 50% pada kondisi pH
2.0 mampu menunjukkan bahwa BAL tersebut mempunyai ketahanan hidup yang
tinggi. Isolat tersebut adalah isolat 2B4, 1B1, 1A5, 2C2, 2D1, 2B2, 1C4, 2B1,
1A32 dan 2C12. Isolat BAL lainnya sebanyak 18 isolat tidak dapat hidup dengan
baik pada pH 2.0. Hal ini ditunjukkan dengan nilai persentase ketahanan hidup di
bawah 50% dan juga populasi akhir yang tidak mencapai 105 cfu/ml, walaupun
populasi awalnya cukup tinggi (9-10 log cfu/ml).
Secara umum, nilai persentase ketahanan hidup semua isolat BAL pada
pH 2.5 lebih baik dibandingkan dengan kondisi pada pH 2.0, yaitu di atas 40%,
dengan kisaran 47.13-88.71% dan rataan 68.05% Sebanyak 14 isolat BAL
mempunyai nilai ketahanan hidup pada pH 2.5 di atas 70%, termasuk diantaranya

54
adalah kesepuluh isolat BAL yang mampu bertahan hidup pada pH 2.0. Demikian
juga halnya dengan ketahanan hidup isolat BAL pada pH 3.2 yang lebih baik
dibandingkan dengan kondisi pH 2.5. Semua isolat BAL mampu bertahan hidup
dengan nilai persentase ketahanan hidup di atas 60%, dengan kisaran 67.55-
109.90%, dengan rataan 85.33%. Kesepuluh isolat yang mampu bertahan hidup
pada pH 2.0 dan 2.5 menunjukkan nilai ketahanan hidup pada pH 3.2 lebih baik
dengan nilai di atas 80%. Namun demikian, tidak semua isolat BAL mempunyai
ketahanan hidup pada pH 7.2 yang lebih tinggi dibandingkan dengan kondisi pH
3.2. Sebanyak 17 isolat BAL menunjukkan adanya populasi yang lebih rendah
dibandingkan dengan populasi pada pH 3.2. Namun, kesemuanya tetap berada
pada nilai persentase ketahanan hidup di atas 78%, kecuali satu isolat yang hanya
mampu bertahan hidup sebesar 62%. Kesepuluh isolat yang mempunyai nilai
kemampuan bertahan hidup pada pH 2.0 cukup tinggi, mampu menunjukkan
ketahanannya pada pH 7.2 yaitu di atas 79%, bahkan ada yang melampaui
populasi awalnya yaitu isolat 1A32, 2C12 dan 2C2 dengan ketahanan hidup diatas
100%. Hal ini menunjukkan bahwa isolat yang mampu bertahan pada pH 2.0 juga
mampu bertahan dengan baik pada pH yang lebih tinggi sampai pH 7.2. Kondisi
ini sangat penting untuk menseleksi isolat BAL yang akan digunakan pada
tahapan karakterisasi sifat probiotik selanjutnya. Secara umum, isolat 2C12 dan
2B4 mempunyai ketahanan hidup pada pH rendah dan pH 7.2 (pH usus) yang
lebih baik dibandingkan dengan isolat lainnya.
Sebagian besar mikroorganisme akan mati dan rusak dengan adanya
pengaruh pH yang rendah dan kondisi asam klorida di dalam lambung. Pada
manusia, waktu transit dari makanan masuk ke mulut sampai lambung minimal
sekitar 90 menit, dan efek bakterisidal asam akan terjadi pada pH asam (Kimoto-
Nira et al. 2007). Bila bakteri terpapar oleh asam kuat, maka membran sel akan
rusak sehingga beberapa komponen intraseluler akan keluar dari sel, di antaranya
ion Mg, Ca, K, asam nukleat dan protein. Akibatnya, sel bakteri akan mengalami
kematian. Hal inilah yang menyebabkan terjadinya penurunan populasi isolat
BAL yang ditumbuhkan pada kondisi pH 2.0, 2.5 serta 3.2. Walaupun demikian,
isolat BAL mempunyai sistem regulasi sel yang mampu mengatur kondisi pH
intraselulernya sehingga mampu bertahan pada kondisi pH rendah.

55
BAL mempunyai mekanisme homeostatis instrinsik yang
menyebabkannya mampu bertahan pada kondisi pH rendah atau keasaman yang
tinggi. Mekanisme yang terjadi pada BAL adalah sistem glutamat-dekarboksilase
(GAD), sistem arginin deiminasi (ADI) dan pompa proton H+ - ATP ase. Melalui
salah satu dari ketiga mekanisme tersebut, BAL mampu meningkatkan pH
intaseluler dengan memproduksi γ-aminobutirat dari dekarboksilasi glutamat di
dalam sel, ataupun memproduksi amonia dari katabolisme arginin (Cotter & Hill
2003).
Hasil penelitian ini memperkuat beberapa penelitian lainnya yang
menunjukkan bahwa tidak semua galur BAL yang diisolasi dari pangan
mempunyai ketahanan hidup yang baik pada kondisi pH yang rendah di antaranya
penelitian Lin et al. (2006); Moyano et al. (2008); Mourad dan Eddine (2006)
serta Mourad dan Meriem (2008). Lin et al (2006) melaporkan pada yogurt yang
mengandung BAL L. acidophilus dan Bifidobacteria mengalami penurunan
jumlah populasi bakteri yang mampu bertahan hidup pada pH 2.0 selama tiga jam.
Populasi total awal BAL pada produk yogurt cair yang mengandung L.
acidophilus dan Bifidobacteria sebesar 108 cfu/ml, dan mampu bertahan sekitar
105 cfu/ml pada pH 2.0.
Sumber pangan seperti sosis fermentasi (Moyano et al. 2008), buah zaitun
fermentasi (Mourad & Eddine 2006), mentega susu onta (Mourad & Meriem
2008), tanaman horseradish (Gbassi 2011) berpotensi probiotik karena
mengandung L. plantarum yang mampu tahan pada pH rendah sesuai pH
lambung. Moyano et al. (2008) menseleksi BAL yang diisolasi dari sosis
fermentasi Iberian sebagai probiotik. Sebanyak 15 isolat dari 173 isolat mampu
bertahan pada kondisi pH 2.5 selama 1.5 jam dengan penurunan populasi akhir
maksimal 2 log cfu/ml, dengan populasi awal sebesar 108 cfu/ml. Isolat yang
mampu bertahan tersebut dinyatakan sebagai kandidat probiotik untuk digunakan
pada kultur starter produk sosis fermentasi.
Peneliti lainnya yaitu Mourad dan Eddine (2006) melakukan seleksi in
vitro probiotik untuk isolat L. plantarum yang diisolasi dari buah zaitun
terfermentasi. Sebanyak 11 isolat L. plantarum tidak mampu hidup pada pH 1.0
selama dua jam, persentase ketahanan hidupnya pada pH 2.0 selama dua jam

56
berkisar 33-65% dan pada pH 2.0 selama empat jam berkisar pada 18-53%,
selanjutnya menurun pada pH 2.0 selama enam jam dengan persentase ketahanan
hidup berkisar pada 11-28%. Mourad dan Meriem (2008) melaporkan bahwa dua
galur L. plantarum yang diisolasi dari mentega susu onta daerah Sahara Algeria
mampu bertahan hidup pada pH 2.0 selama dua jam inkubasi dengan tingkat
ketahanan hidup untuk L. plantarum SH12 sebesar 54% dan L. plantarum SH24
sebesar 55%. Gbassi (2011) juga melaporkan bahwa tiga galur L. plantarum,
yang salah satunya diisolasi dari tanaman horseradish, mampu bertahan hidup
pada kompartemen lambung dan jejunum secara in vitro.
Sepuluh isolat BAL yang berdasarkan seleksi awal mampu bertahan
dengan baik pada pH 2.0, 2.5, 3.2, dan 7.2, dengan minimal ketahanan hidup pada
pH 2.0 ≥ 50% dipilih untuk dilanjutkan pengujian sifat probiotik. Pengujian
tersebut meliputi ketahanan hidup pada garam empedu, aktivitas antimikroba
terhadap bakteri patogen, koagregasi terhadap bakteri patogen, serta sifat
penempelannya pada sel epitel usus secara in vitro. Kesepuluh isolat tersebut
adalah 2B4, 1B1, 1A5, 2C2, 2D1, 2B2, 1C4, 2B1, 1A32 dan 2C12.
Ketahanan Hidup BAL terhadap Garam Empedu
Konsentrasi garam empedu di usus halus adalah sekitar 0.3-0.5%. Pada
manusia normal, waktu transit makanan dari mulut sampai usus halus adalah
antara 4-6 jam akhirnya di usus besar selama 24-48 jam (Bourlioux et al. 2003).
Oleh karenanya pada penelitian ini digunakan konsentrasi garam empedu sebesar
0.5% selama enam jam inkubasi berdasarkan konsentrasi terbesar garam empedu
dan kisaran tertinggi lama waktu transit makanan sampai sekum. Ketahanan
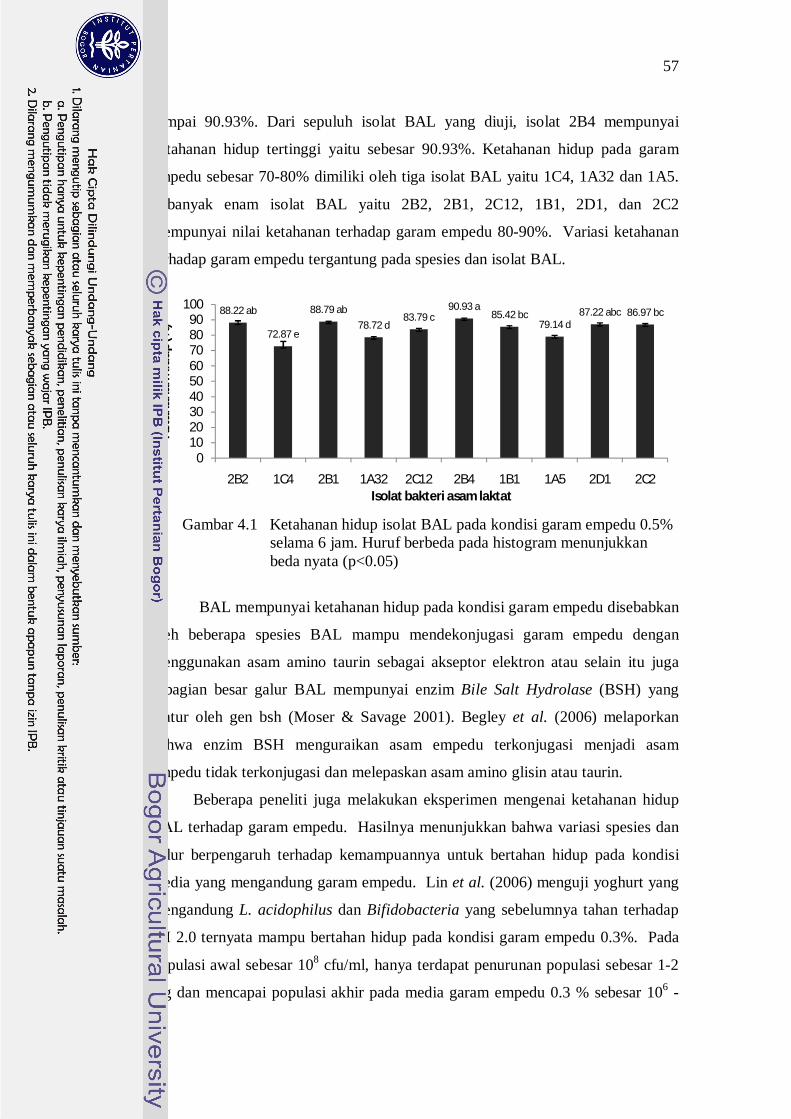
hidup isolat BAL pada kondisi garam empedu ditunjukkan pada Gambar 4.1.
Populasi awal kesepuluh isolat BAL yang tahan pada pH rendah adalah
berkisar pada 108 -109 cfu/ml. Setelah diinokulasikan ke dalam media MRS yang
diberikan garam empedu 0.5% maka terjadi penurunan sebesar 1-3 log dengan
populasi berkisar pada 106 - 108 cfu/ml. Nilai ketahanan terhadap garam empedu
berbeda nyata (p<0.05) pada semua isolat BAL (Gambar 4.1). Secara umum,
kesepuluh isolat BAL yang tahan pada pH 2.0 juga mempunyai ketahanan hidup
pada garam empedu 0.5% selama 6 jam yang baik yaitu berkisar antara 72.87
57
sampai 90.93%. Dari sepuluh isolat BAL yang diuji, isolat 2B4 mempunyai
ketahanan hidup tertinggi yaitu sebesar 90.93%. Ketahanan hidup pada garam
empedu sebesar 70-80% dimiliki oleh tiga isolat BAL yaitu 1C4, 1A32 dan 1A5.
Sebanyak enam isolat BAL yaitu 2B2, 2B1, 2C12, 1B1, 2D1, dan 2C2
mempunyai nilai ketahanan terhadap garam empedu 80-90%. Variasi ketahanan
terhadap garam empedu tergantung pada spesies dan isolat BAL.
Gambar 4.1 Ketahanan hidup isolat BAL pada kondisi garam empedu 0.5% selama 6 jam. Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
BAL mempunyai ketahanan hidup pada kondisi garam empedu disebabkan
oleh beberapa spesies BAL mampu mendekonjugasi garam empedu dengan
menggunakan asam amino taurin sebagai akseptor elektron atau selain itu juga
sebagian besar galur BAL mempunyai enzim Bile Salt Hydrolase (BSH) yang
diatur oleh gen bsh (Moser & Savage 2001). Begley et al. (2006) melaporkan
bahwa enzim BSH menguraikan asam empedu terkonjugasi menjadi asam
empedu tidak terkonjugasi dan melepaskan asam amino glisin atau taurin.
Beberapa peneliti juga melakukan eksperimen mengenai ketahanan hidup
BAL terhadap garam empedu. Hasilnya menunjukkan bahwa variasi spesies dan
galur berpengaruh terhadap kemampuannya untuk bertahan hidup pada kondisi
media yang mengandung garam empedu. Lin et al. (2006) menguji yoghurt yang
mengandung L. acidophilus dan Bifidobacteria yang sebelumnya tahan terhadap
pH 2.0 ternyata mampu bertahan hidup pada kondisi garam empedu 0.3%. Pada
populasi awal sebesar 108 cfu/ml, hanya terdapat penurunan populasi sebesar 1-2
log dan mencapai populasi akhir pada media garam empedu 0.3 % sebesar 106 -
88.22 ab
72.87 e
88.79 ab78.72 d
83.79 c90.93 a
85.42 bc79.14 d
87.22 abc 86.97 bc
0102030405060708090
100
2B2 1C4 2B1 1A32 2C12 2B4 1B1 1A5 2D1 2C2
Keta
hana
n hi
dup
(%)
Isolat bakteri asam laktat

58
107 cfu/ml. Pereira et al. (2003) menyatakan galur L. fermentum KC5b
mempunyai toleransi yang tinggi terhadap kondisi asam lambung dan garam
empedu dan juga aktivitas BSH-nya tinggi. Mourad dan Meriem (2008)
melaporkan bahwa galur L. plantarum SH 12 mempunyai ketahanan terhadap
garam empedu 2% sebesar 75% yang lebih tinggi dibandingkan dengan galur L.
plantarum SH 21 yaitu sebesar 65%.
Aktivitas Antimikroba BAL terhadap Bakteri Patogen
Bakteri patogen yang digunakan pada eksperimen pengujian aktivitas
antimikroba isolat BAL terlebih dahulu dihitung populasi awalnya. Jumlah bakteri
ini merupakan jumlah yang diinokulasikan pada cawan untuk prosedur difusi
agar. Kesemua bakteri patogen telah memenuhi jumlah yang diinginkan untuk
pengujian yaitu sebesar 106 cfu/ml (Tabel 4.4).
Tabel 4.4 Populasi bakteri patogen yang digunakan pada pengujian aktivitas antimikroba BAL
Jenis Bakteri Uji Populasi (cfu/ml)
E. coli ATCC 25922 3.2 x 106
S. aureus ATCC 25923 5.6 x 106
S. Typhimurium ATCC 14028 4.4 x 106
E. coli enteropatogen (EPEC) 4.0 x 106
Supernatan bebas sel yang digunakan pada pengujian aktivitas antimikroba
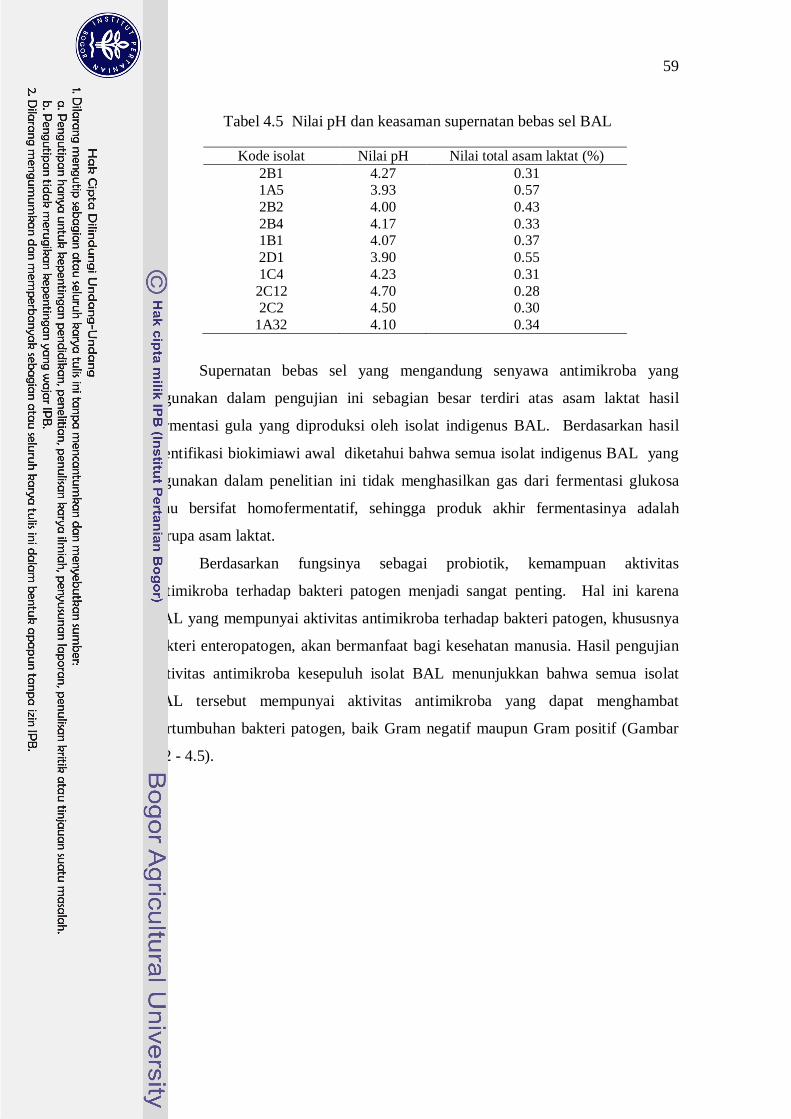
juga diukur pH dan total asam tertitrasinya. Nilai pH semua supernatan bebas sel
dari seluruh isolat BAL berkisar 3.90-4.70, sedangkan nilai total asam laktatnya
berkisar 0.28-0.57% (Tabel 4.5).
59
Tabel 4.5 Nilai pH dan keasaman supernatan bebas sel BAL
Kode isolat Nilai pH Nilai total asam laktat (%) 2B1 4.27 0.31 1A5 3.93 0.57 2B2 4.00 0.43 2B4 4.17 0.33 1B1 4.07 0.37 2D1 3.90 0.55 1C4 4.23 0.31 2C12 4.70 0.28 2C2 4.50 0.30
1A32 4.10 0.34
Supernatan bebas sel yang mengandung senyawa antimikroba yang
digunakan dalam pengujian ini sebagian besar terdiri atas asam laktat hasil
fermentasi gula yang diproduksi oleh isolat indigenus BAL. Berdasarkan hasil
identifikasi biokimiawi awal diketahui bahwa semua isolat indigenus BAL yang
digunakan dalam penelitian ini tidak menghasilkan gas dari fermentasi glukosa
atau bersifat homofermentatif, sehingga produk akhir fermentasinya adalah
berupa asam laktat.
Berdasarkan fungsinya sebagai probiotik, kemampuan aktivitas
antimikroba terhadap bakteri patogen menjadi sangat penting. Hal ini karena
BAL yang mempunyai aktivitas antimikroba terhadap bakteri patogen, khususnya
bakteri enteropatogen, akan bermanfaat bagi kesehatan manusia. Hasil pengujian
aktivitas antimikroba kesepuluh isolat BAL menunjukkan bahwa semua isolat
BAL tersebut mempunyai aktivitas antimikroba yang dapat menghambat
pertumbuhan bakteri patogen, baik Gram negatif maupun Gram positif (Gambar
4.2 - 4.5).
60
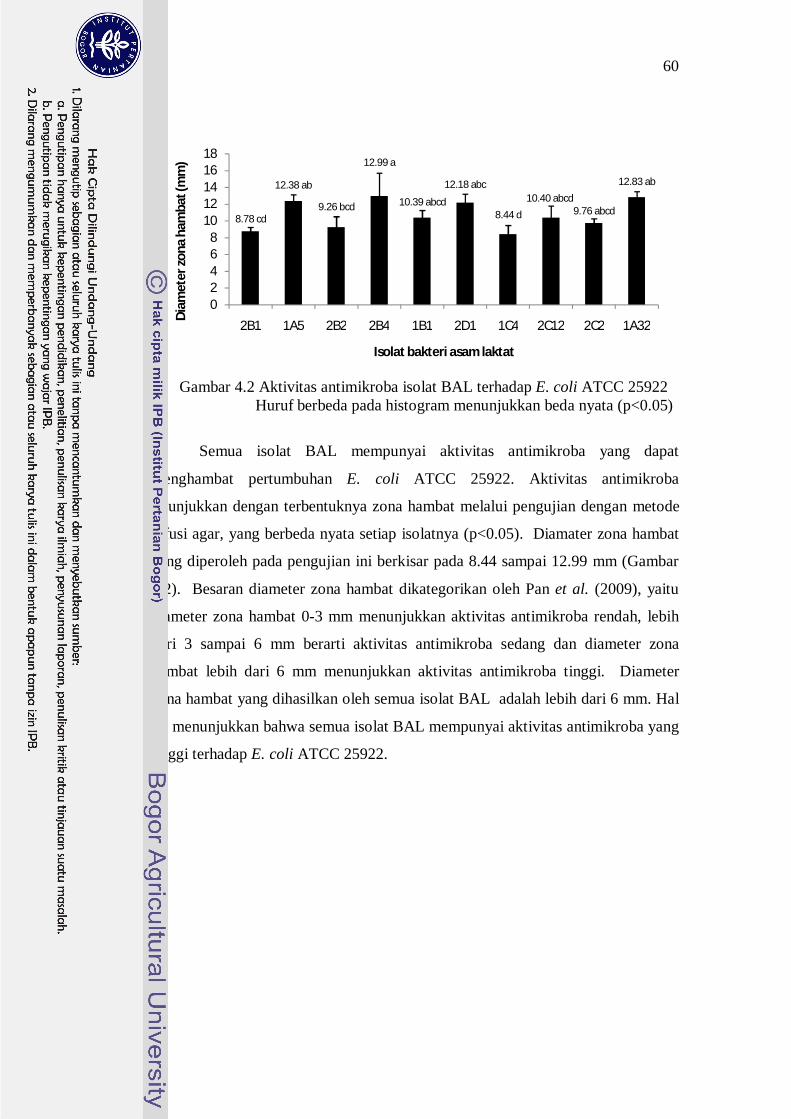
Gambar 4.2 Aktivitas antimikroba isolat BAL terhadap E. coli ATCC 25922 Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
Semua isolat BAL mempunyai aktivitas antimikroba yang dapat
menghambat pertumbuhan E. coli ATCC 25922. Aktivitas antimikroba
ditunjukkan dengan terbentuknya zona hambat melalui pengujian dengan metode
difusi agar, yang berbeda nyata setiap isolatnya (p<0.05). Diamater zona hambat
yang diperoleh pada pengujian ini berkisar pada 8.44 sampai 12.99 mm (Gambar
4.2). Besaran diameter zona hambat dikategorikan oleh Pan et al. (2009), yaitu
diameter zona hambat 0-3 mm menunjukkan aktivitas antimikroba rendah, lebih
dari 3 sampai 6 mm berarti aktivitas antimikroba sedang dan diameter zona
hambat lebih dari 6 mm menunjukkan aktivitas antimikroba tinggi. Diameter
zona hambat yang dihasilkan oleh semua isolat BAL adalah lebih dari 6 mm. Hal
ini menunjukkan bahwa semua isolat BAL mempunyai aktivitas antimikroba yang
tinggi terhadap E. coli ATCC 25922.
8.78 cd
12.38 ab
9.26 bcd
12.99 a
10.39 abcd
12.18 abc
8.44 d10.40 abcd
9.76 abcd
12.83 ab
02468
1012141618
2B1 1A5 2B2 2B4 1B1 2D1 1C4 2C12 2C2 1A32
Diam
eter
zona
ham
bat (
mm
)
Isolat bakteri asam laktat
61
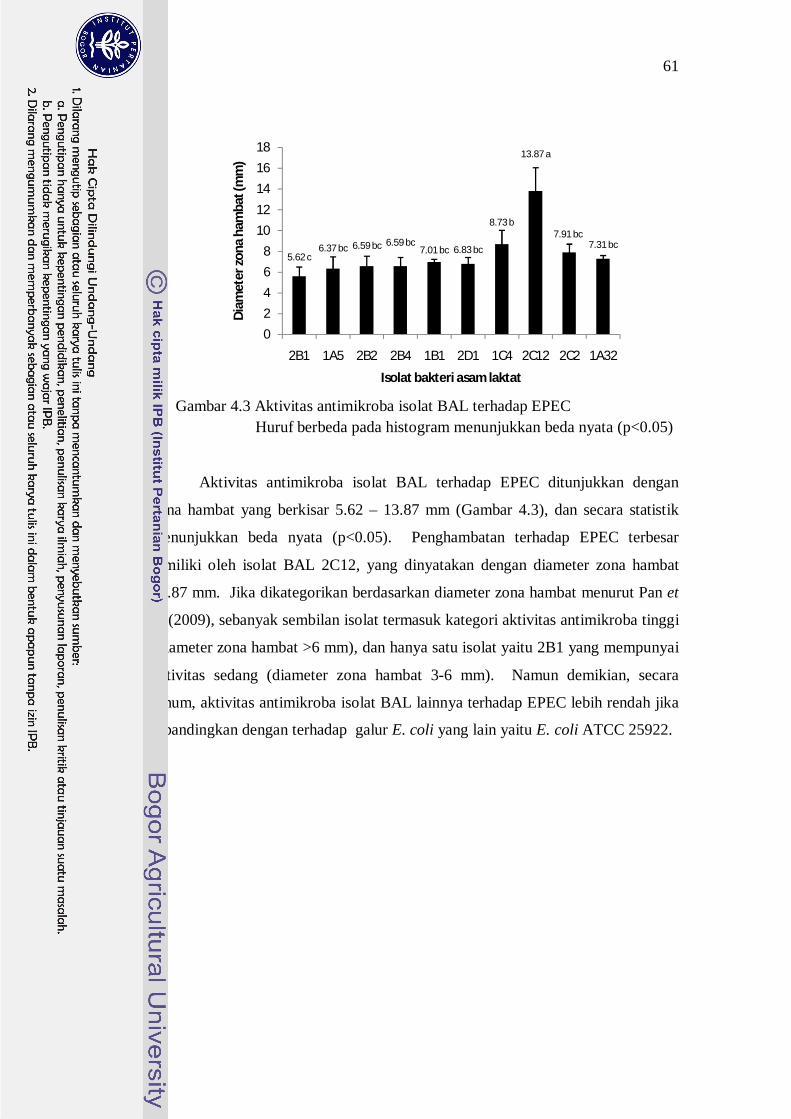
Gambar 4.3 Aktivitas antimikroba isolat BAL terhadap EPEC Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
Aktivitas antimikroba isolat BAL terhadap EPEC ditunjukkan dengan
zona hambat yang berkisar 5.62 – 13.87 mm (Gambar 4.3), dan secara statistik
menunjukkan beda nyata (p<0.05). Penghambatan terhadap EPEC terbesar
dimiliki oleh isolat BAL 2C12, yang dinyatakan dengan diameter zona hambat
13.87 mm. Jika dikategorikan berdasarkan diameter zona hambat menurut Pan et
al (2009), sebanyak sembilan isolat termasuk kategori aktivitas antimikroba tinggi
(diameter zona hambat >6 mm), dan hanya satu isolat yaitu 2B1 yang mempunyai
aktivitas sedang (diameter zona hambat 3-6 mm). Namun demikian, secara
umum, aktivitas antimikroba isolat BAL lainnya terhadap EPEC lebih rendah jika
dibandingkan dengan terhadap galur E. coli yang lain yaitu E. coli ATCC 25922.
5.62 c6.37 bc 6.59 bc 6.59 bc
7.01 bc 6.83 bc
8.73 b
13.87 a
7.91 bc7.31 bc
02468
1012141618
2B1 1A5 2B2 2B4 1B1 2D1 1C4 2C12 2C2 1A32
Diam
eter
zona
ham
bat (
mm
)
Isolat bakteri asam laktat
62
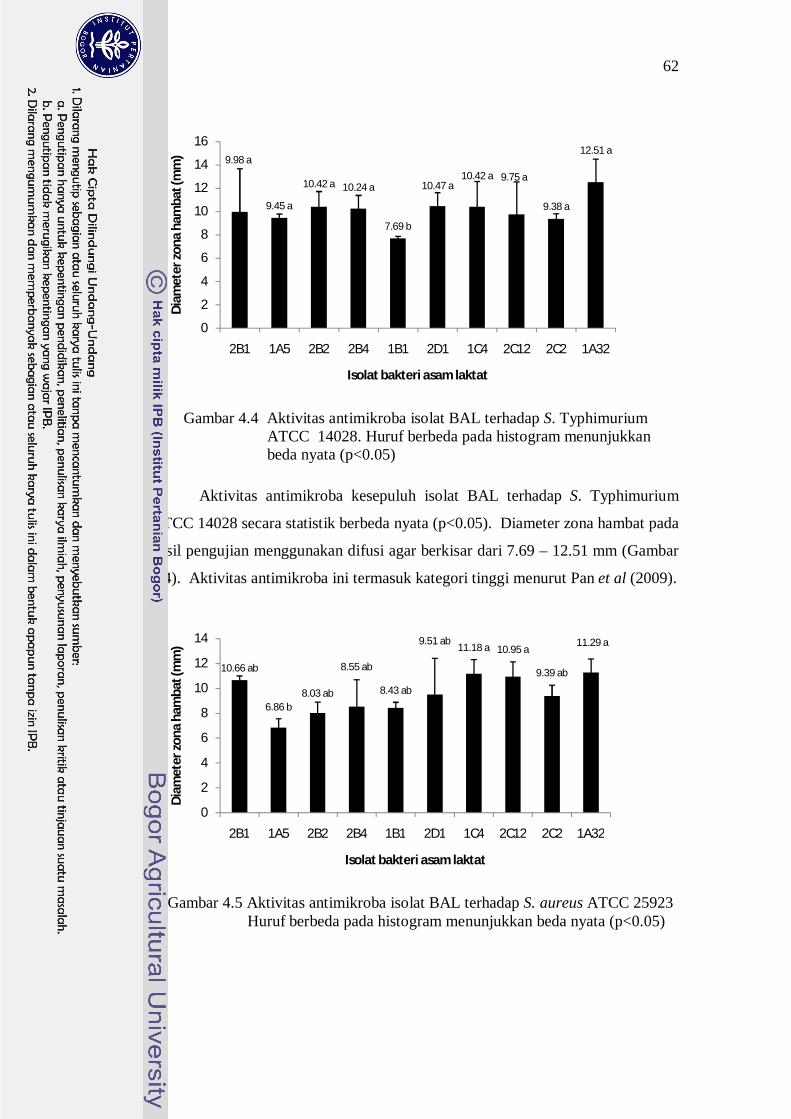
Gambar 4.4 Aktivitas antimikroba isolat BAL terhadap S. Typhimurium ATCC 14028. Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
Aktivitas antimikroba kesepuluh isolat BAL terhadap S. Typhimurium
ATCC 14028 secara statistik berbeda nyata (p<0.05). Diameter zona hambat pada
hasil pengujian menggunakan difusi agar berkisar dari 7.69 – 12.51 mm (Gambar
4.4). Aktivitas antimikroba ini termasuk kategori tinggi menurut Pan et al (2009).
Gambar 4.5 Aktivitas antimikroba isolat BAL terhadap S. aureus ATCC 25923 Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
9.98 a
9.45 a
10.42 a 10.24 a
7.69 b
10.47 a10.42 a 9.75 a
9.38 a
12.51 a
0
2
4
6
8
10
12
14
16
2B1 1A5 2B2 2B4 1B1 2D1 1C4 2C12 2C2 1A32
Diam
eter
zona
ham
bat (
mm
)
Isolat bakteri asam laktat
10.66 ab
6.86 b8.03 ab
8.55 ab
8.43 ab
9.51 ab11.18 a 10.95 a
9.39 ab
11.29 a
0
2
4
6
8
10
12
14
2B1 1A5 2B2 2B4 1B1 2D1 1C4 2C12 2C2 1A32
Diam
eter
zona
ham
bat (
mm
)
Isolat bakteri asam laktat

63
Penghambatan terhadap S. aureus oleh senyawa antimikroba yang
diproduksi oleh isolat BAL dipengaruhi oleh spesies bahkan galur dari isolat
BAL dan berbeda nyata (p<0.05) (Gambar 4.5). Aktivitas antimikroba yang
ditunjukkan dengan diameter zona hambat terhadap S. aureus ATCC 25923
berkisar 6.86-11.29 mm. Kesembilan BAL selain 1A5 mempunyai aktivitas daya
hambat yang tidak berbeda, namun 1A5 mempunyai aktivitas antimikroba
terendah. Walaupun demikian, dengan nilai diameter zona hambat di atas 6 mm,
jika dikategorikan sesuai dengan pendapat Pan et al. (2009) maka aktivitas
antimikroba dari kesepuluh isolat BAL termasuk kategori tinggi.
Dibandingkan dengan jenis patogen yang digunakan, aktivitas antimikroba
semua isolat BAL dimulai dari aktivitas antimikroba tertinggi berturut-turut
adalah terhadap E. coli ATCC 25922, S. Typhimurium ATCC 14028, S.aureus
ATCC 25923 serta aktivitas antimikroba terendah terhadap EPEC. Isolat BAL
lebih mampu menghambat bakteri Gram negatif (E coli ATCC 25922 dan S
Typhimurium ATCC 14028) kecuali terhadap EPEC, dibandingkan dengan Gram
positif S. aureus ATCC 25923. Dilaporkan oleh Fitrial (2009) dan Miksusanti
(2009), EPEC merupakan bakteri yang resisten terhadap beberapa jenis
antibiotika. Pada dosis antibiotik 2% b/v, EPEC K11 tidak dapat dihambat
pertumbuhannya oleh antibiotik amoksilin dan ampisilin (diameter zona hambat 0
mm), sedangkan penghambatan yang sangat rendah (diameter zona hambat 1.72
mm) ditunjukkan oleh antibiotik kloramfenikol (Fitrial 2009; Miksusanti 2009).
Tingkat resistensi EPEC terhadap senyawa antibiotik yang cukup tinggi ini
selaras dengan resistensi EPEC terhadap senyawa antimikroba yang diproduksi
oleh isolat BAL. Jika dilihat dari zona hambat yang dihasilkan oleh isolat BAL
terhadap EPEC, isolat 2C12 mampu melakukan penghambatan yang signifikan
lebih tinggi dibandingkan dengan isolat lainnya. Secara umum, isolat 2C12 juga
mempunyai aktvitas penghambatan yang tinggi terhadap E. coli 25922, S.
Typhimurium ATCC 14028 serta S. aureus ATCC 25922. Diameter zona hambat
isolat 2C12 terhadap bakteri patogen ditunjukkan pada Gambar 4.6.
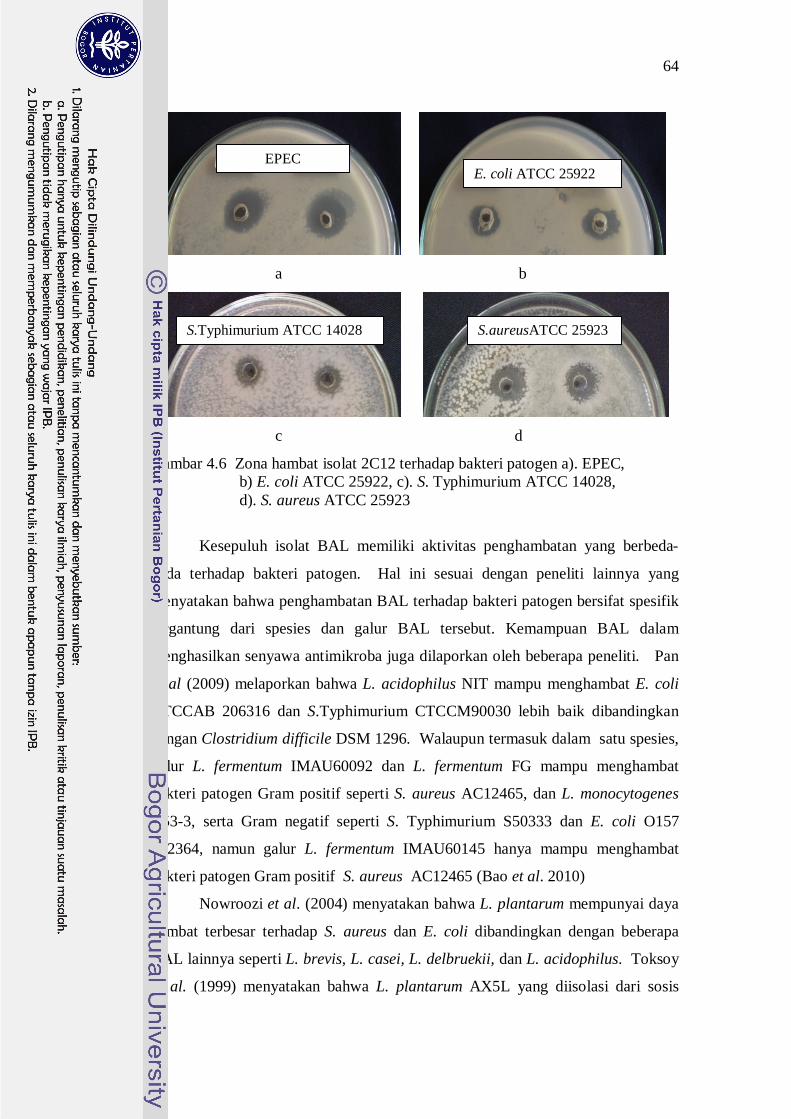
64
a b
c d
Gambar 4.6 Zona hambat isolat 2C12 terhadap bakteri patogen a). EPEC, b) E. coli ATCC 25922, c). S. Typhimurium ATCC 14028, d). S. aureus ATCC 25923
Kesepuluh isolat BAL memiliki aktivitas penghambatan yang berbeda-
beda terhadap bakteri patogen. Hal ini sesuai dengan peneliti lainnya yang
menyatakan bahwa penghambatan BAL terhadap bakteri patogen bersifat spesifik
tergantung dari spesies dan galur BAL tersebut. Kemampuan BAL dalam
menghasilkan senyawa antimikroba juga dilaporkan oleh beberapa peneliti. Pan
et al (2009) melaporkan bahwa L. acidophilus NIT mampu menghambat E. coli
CTCCAB 206316 dan S.Typhimurium CTCCM90030 lebih baik dibandingkan
dengan Clostridium difficile DSM 1296. Walaupun termasuk dalam satu spesies,
galur L. fermentum IMAU60092 dan L. fermentum FG mampu menghambat
bakteri patogen Gram positif seperti S. aureus AC12465, dan L. monocytogenes
C53-3, serta Gram negatif seperti S. Typhimurium S50333 dan E. coli O157
882364, namun galur L. fermentum IMAU60145 hanya mampu menghambat
bakteri patogen Gram positif S. aureus AC12465 (Bao et al. 2010)
Nowroozi et al. (2004) menyatakan bahwa L. plantarum mempunyai daya
hambat terbesar terhadap S. aureus dan E. coli dibandingkan dengan beberapa
BAL lainnya seperti L. brevis, L. casei, L. delbruekii, dan L. acidophilus. Toksoy
et al. (1999) menyatakan bahwa L. plantarum AX5L yang diisolasi dari sosis
S.Typhimurium ATCC 14028 S.aureusATCC 25923
EPEC E. coli ATCC 25922

65
dapat menghambat E. coli, S. aureus dan B. subtilis karena L. plantarum AX5L
mampu menghasilkan H2O2, asam laktat sebesar 0.88% dan bakteriosin
plantarisin. Streptococcus lactis memiliki aktivitas bakterisidal terhadap bakteri
Gram positif maupun Gram negatif antara lain Enterococcus faecalis, Bacillus
subtilis, S. Typhimurium dan E. coli (Suarsana et al. 2001).
Aktivitas antimikroba setiap isolat BAL yang berbeda terhadap spesies
bakteri patogen yang berbeda disebabkan oleh komponen antimikroba yang
dihasilkan oleh setiap isolat yang juga berbeda. Aktivitas antimikroba BAL
disebabkan terutama oleh asam organik yang diproduksi sebagai hasil
metabolisme glukosa. Pada penelitian ini, nilai total asam laktat berkisar pada
0.28-0.57% dengan kisaran pH 3.90-4.70. Asam laktat merupakan asam lemah
tidak terdisosiasi yang mempunyai nilai pKa 3.86 pada suhu 25 °C (Bogaert &
Naidu 2000). Mekanisme penghambatan asam laktat terhadap sel bakteri karena
asam laktat mempunyai sifat hidrofobik, sehingga memudahkan difusi dalam
bentuk proton ke dalam sel melalui membran sel. Akibatnya pH intraseluler lebih
tinggi dibandingkan dengan pH ekstraseluler. Selanjutnya, di dalam sel, asam
laktat terdisosiasi dan menurunkan pH intraseluler dengan melepaskan proton
(Bogaert & Naidu 2000). Pelepasan proton/ ion hidrogen dapat mengganggu
fungsi metabolik seperti translokasi substrat dan fosforilasi oksidatif, sehingga
menyebabkan sel bakteri tersebut terhambat pertumbuhannya.
Selain asam laktat, BAL juga memproduksi senyawa antimikroba lainnya,
di antaranya hidrogen peroksida dan senyawa bakteriosin (Bao et al. 2010). BAL
memproduksi H2O2 (hidrogen peroksida) melalui transport elektron via enzim
flavin. Dengan adanya H2O2, bentuk anion superoksida merusak radikal hidroksi.
Proses antimikrobanya melibatkan peroksidase lipid membran dan meningkatkan
permeabilitas membran. Hasilnya adalah efek bakterisidal dari metabolit oksigen
yang mengakibatkan terjadinya oksidasi sel bakteri dan akhirnya merusak asam
nukleat dan protein sel (Naidu & Clemens 2000).
Senyawa antimikroba lainnya yang diproduksi oleh BAL adalah bakteriosin.
Bakteriosin merupakan molekul protein atau peptida ekstraseluler yang
mempunyai aksi bakterisidal atau bakteriostatik terhadap bakteri yang mempunyai
kekerabatan dekat (Savadogo et al. 2006; Albano et al. 2007). Bakteriosin dapat

66
didegradasi oleh enzim protease dalam saluran pencernaan. Bakteriosin bersifat
irrevesible, mudah dicerna, berpengaruh positif terhadap kesehatan dan aktif pada
konsentrasi rendah. (Pal et al. 2005; Savadogo et al. 2006). Mekanisme
antimikroba bakterisoin dimulai dengan masuknya ke dalam sel sasarannya
dengan cara membentuk pori di membran sel yang sensitif dan menurunkan
potensial atau gradien pH yang menyebabkan rusaknya material seluler sehingga
mampu menghambat pertumbuhan sel target (Ogunbawo et al. 2003; Moll et al.
1999).
Koagregasi BAL terhadap Bakteri Patogen
Bakteri probiotik dan patogen dapat membentuk gabungan agregrat yang
disebut dengan koagregasi (Surono 2004). Probiotik yang mampu melakukan
koagregasi dengan patogen akan efektif menghambat dan membunuh bakteri
patogen karena senyawa antimikroba yang dihasilkan dapat beraktivitas langsung
pada patogen (Surono 2004; Bao et al. 2010). Pada penelitian ini, dilakukan
pengujian koagregasi antara sepuluh isolat indigenus BAL dengan bakteri patogen
E. coli ATCC 25922, EPEC, S.Typhimurium ATCC 14028 dan S. aureus ATCC
25923. Pengujian ini juga bertujuan untuk mengetahui kemampuan interaksi
antara isolat BAL dengan bakteri patogen. Nilai koagregasi isolat indigenus BAL
terhadap bakteri patogen ditunjukkan pada Gambar 4.7-4.10.
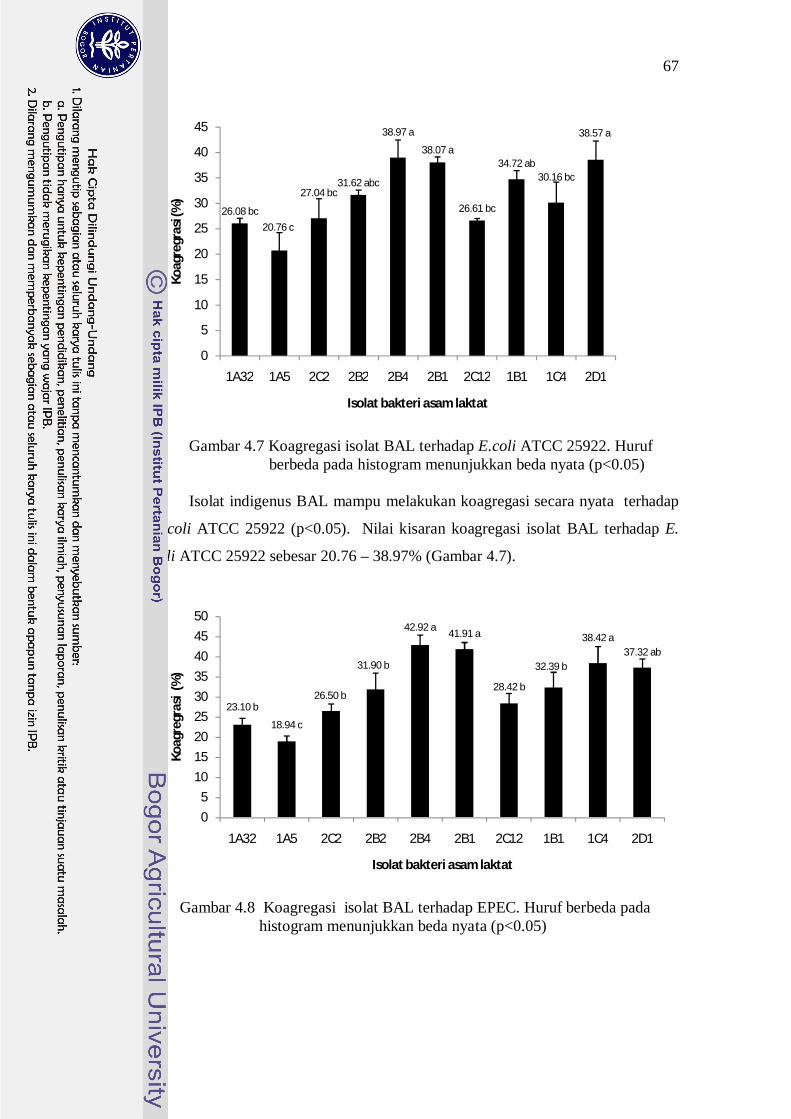
67
Gambar 4.7 Koagregasi isolat BAL terhadap E.coli ATCC 25922. Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05) Isolat indigenus BAL mampu melakukan koagregasi secara nyata terhadap
E coli ATCC 25922 (p<0.05). Nilai kisaran koagregasi isolat BAL terhadap E.
coli ATCC 25922 sebesar 20.76 – 38.97% (Gambar 4.7).
Gambar 4.8 Koagregasi isolat BAL terhadap EPEC. Huruf berbeda pada
histogram menunjukkan beda nyata (p<0.05)
26.08 bc20.76 c
27.04 bc31.62 abc
38.97 a
38.07 a
26.61 bc
34.72 ab30.16 bc
38.57 a
0
5
10
15
20
25
30
35
40
45
1A32 1A5 2C2 2B2 2B4 2B1 2C12 1B1 1C4 2D1
Koag
regr
asi (
%)
Isolat bakteri asam laktat
23.10 b
18.94 c
26.50 b
31.90 b
42.92 a 41.91 a
28.42 b
32.39 b
38.42 a37.32 ab
05
101520253035404550
1A32 1A5 2C2 2B2 2B4 2B1 2C12 1B1 1C4 2D1
Koag
regr
asi (
%)
Isolat bakteri asam laktat
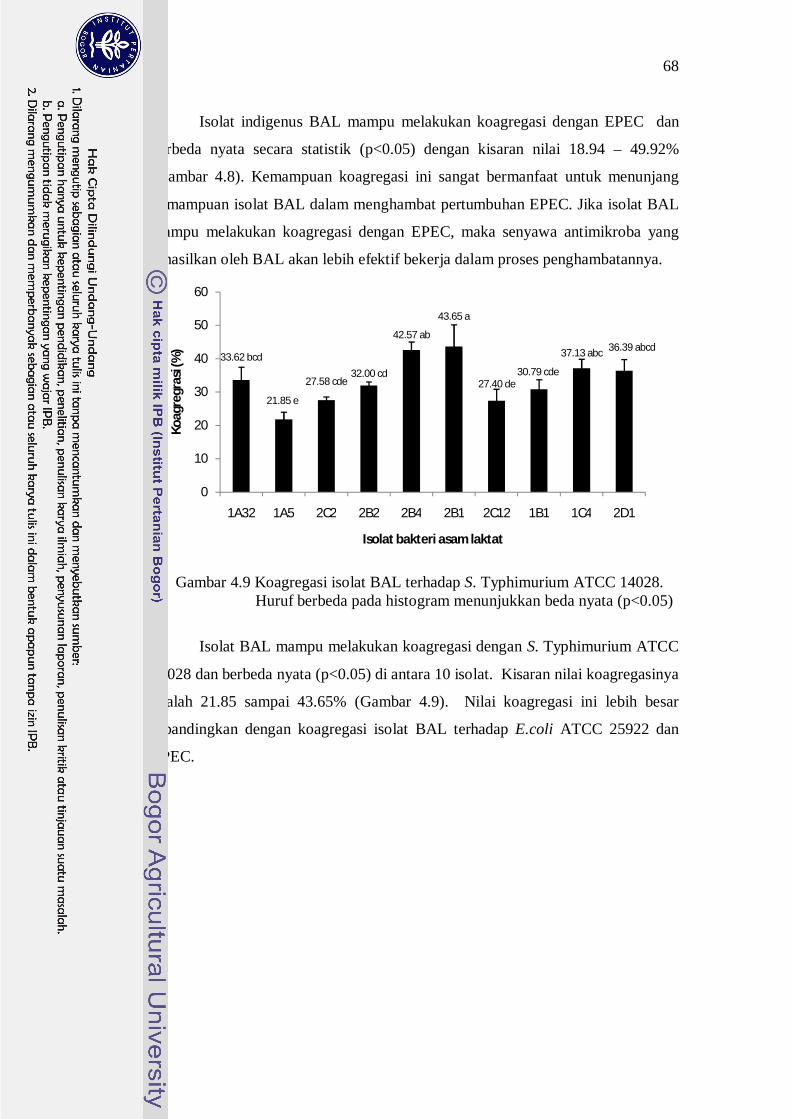
68
Isolat indigenus BAL mampu melakukan koagregasi dengan EPEC dan
berbeda nyata secara statistik (p<0.05) dengan kisaran nilai 18.94 – 49.92%
(Gambar 4.8). Kemampuan koagregasi ini sangat bermanfaat untuk menunjang
kemampuan isolat BAL dalam menghambat pertumbuhan EPEC. Jika isolat BAL
mampu melakukan koagregasi dengan EPEC, maka senyawa antimikroba yang
dihasilkan oleh BAL akan lebih efektif bekerja dalam proses penghambatannya.
Gambar 4.9 Koagregasi isolat BAL terhadap S. Typhimurium ATCC 14028. Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
Isolat BAL mampu melakukan koagregasi dengan S. Typhimurium ATCC
14028 dan berbeda nyata (p<0.05) di antara 10 isolat. Kisaran nilai koagregasinya
adalah 21.85 sampai 43.65% (Gambar 4.9). Nilai koagregasi ini lebih besar
dibandingkan dengan koagregasi isolat BAL terhadap E.coli ATCC 25922 dan
EPEC.
33.62 bcd
21.85 e
27.58 cde32.00 cd
42.57 ab
43.65 a
27.40 de30.79 cde
37.13 abc 36.39 abcd
0
10
20
30
40
50
60
1A32 1A5 2C2 2B2 2B4 2B1 2C12 1B1 1C4 2D1
Koag
regr
asi (
%)
Isolat bakteri asam laktat
69
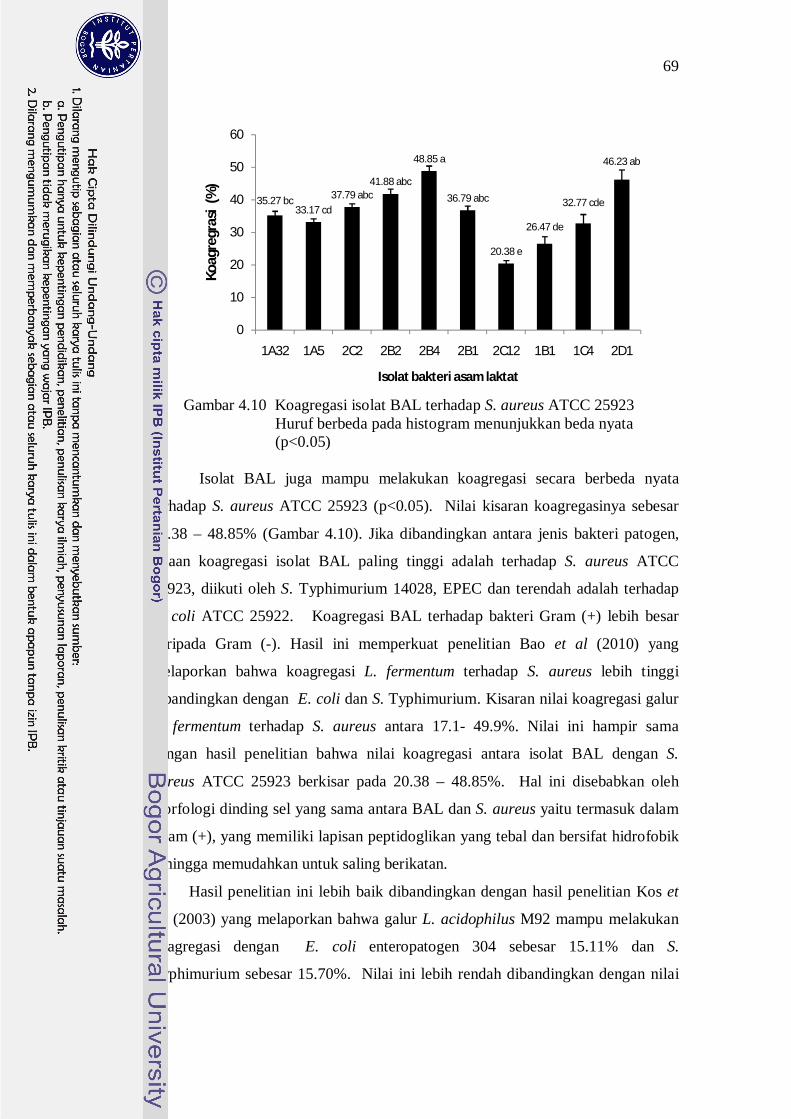
Gambar 4.10 Koagregasi isolat BAL terhadap S. aureus ATCC 25923
Huruf berbeda pada histogram menunjukkan beda nyata (p<0.05)
Isolat BAL juga mampu melakukan koagregasi secara berbeda nyata
terhadap S. aureus ATCC 25923 (p<0.05). Nilai kisaran koagregasinya sebesar
20.38 – 48.85% (Gambar 4.10). Jika dibandingkan antara jenis bakteri patogen,
rataan koagregasi isolat BAL paling tinggi adalah terhadap S. aureus ATCC
25923, diikuti oleh S. Typhimurium 14028, EPEC dan terendah adalah terhadap
E. coli ATCC 25922. Koagregasi BAL terhadap bakteri Gram (+) lebih besar
daripada Gram (-). Hasil ini memperkuat penelitian Bao et al (2010) yang
melaporkan bahwa koagregasi L. fermentum terhadap S. aureus lebih tinggi
dibandingkan dengan E. coli dan S. Typhimurium. Kisaran nilai koagregasi galur
L. fermentum terhadap S. aureus antara 17.1- 49.9%. Nilai ini hampir sama
dengan hasil penelitian bahwa nilai koagregasi antara isolat BAL dengan S.
aureus ATCC 25923 berkisar pada 20.38 – 48.85%. Hal ini disebabkan oleh
morfologi dinding sel yang sama antara BAL dan S. aureus yaitu termasuk dalam
Gram (+), yang memiliki lapisan peptidoglikan yang tebal dan bersifat hidrofobik
sehingga memudahkan untuk saling berikatan.
Hasil penelitian ini lebih baik dibandingkan dengan hasil penelitian Kos et
al. (2003) yang melaporkan bahwa galur L. acidophilus M92 mampu melakukan
koagregasi dengan E. coli enteropatogen 304 sebesar 15.11% dan S.
Typhimurium sebesar 15.70%. Nilai ini lebih rendah dibandingkan dengan nilai
35.27 bc33.17 cd
37.79 abc41.88 abc
48.85 a
36.79 abc
20.38 e
26.47 de
32.77 cde
46.23 ab
0
10
20
30
40
50
60
1A32 1A5 2C2 2B2 2B4 2B1 2C12 1B1 1C4 2D1
Koag
regr
asi
(%)
Isolat bakteri asam laktat

70
koagregasi yang diperoleh pada rataan hasil penelitian ini yaitu 31.26% terhadap
E. coli ATCC 25922, 32.18% terhadap EPEC, 33.30% terhadap S. Typhimurium
ATCC 14028, serta 35.96% terhadap S. aureus ATCC 25923. El-Naggar (2004)
melaporkan bahwa L. acidophilus dan L. plantarum memiliki sifat koagregasi
yang baik dengan E. coli dan S. Typhimurium.
Koagregasi yang tinggi antara BAL dengan bakteri enteropatogen akan
meningkatkan mekanisme pertahanan di saluran pencernaan dengan jalan
mencegah kolonisasi bakteri enteropatogen di saluran pencernaan. Secara umum,
isolat 2B4 mempunyai koagregasi terhadap bakteri patogen yang lebih baik
daripada isolat lainnya. Hal ini sangat penting untuk mendukung efektivitas
penghambatan bakteri patogen di saluran pencernaan.
Kemampuan koagregasi sangat dipengaruhi oleh spesies dan galur yang
berbeda (Rinkinen et al. 2003; Bao et al. 2010). Penelitian Bao et al. (2010)
menunjukkan bahwa sebanyak 11 galur L. fermentum mempunyai sifat
koagregasi yang bervariasi terhadap lima jenis bakteri patogen yaitu L.
monocytogenes, S. aureus, E. coli, S.Typhimurium dan Shigella flexneri dan
hanya dua galur L. fermentum yang mempunyai nilai koagregasi yang tinggi (>
30%).
Penempelan BAL pada Permukaan Usus
Selain sifat ketahanannya terhadap pH rendah dan garam empedu, sifat
BAL sebagai probiotik di saluran pencernaan juga ditentukan dengan uji
penempelan. Berbagai model penempelan secara in vitro dilakukan antara lain
penempelan ke permukaan usus babi oleh Kos et al. (2003), Mishra dan Prasad
(2005) yang melakukan uji penempelan Lactobacillus casei secara in vitro ke sel
usus tikus dan pemodelan penempelan ke stainless steel, Nitisinprasert et al.
(2006) yang melakukan pengujian penempelan BAL menggunakan usus ayam,
serta Blum et al. (1999) yang menggunakan model penempelan BAL sebagai
probiotik secara in vitro ke sel Caco-2.
Kemampuan BAL untuk menempel pada permukaan sel epitel dan mukosa
sangat penting, bukan hanya untuk menjaga keseimbangan jumlah bakteri dalam
saluran pencernaan, tapi juga untuk mencegah asosiasi sel dan invasi bakteri
71
patogen (Ouwehand et al. 2001). Pada pengujian sifat penempelan ini digunakan
permukaan usus tikus bagian ileum yang sudah dihilangkan mukosanya untuk
mengetahui populasi BAL yang menempel. Usus tikus yang tidak diberikan
perlakuan perendaman dengan suspensi BAL digunakan sebagai kontrol. Besaran
penempelan BAL pada permukaan usus dinyatakan dengan menghitung selisih
populasi BAL kontrol dengan populasi BAL pada perlakuan, selanjutnya dihitung
persentase penempelannya. Hasil pengujian penempelan BAL pada permukaan
usus tikus secara in vitro disajikan pada Tabel 4.6.
Tabel 4.6 Penempelan BAL pada permukaan usus tikus secara in vitro
Sampel/isolat BAL
Rataan populasi BAL (log 10 cfu/cm2)
Persentase penempelan BAL ke permukaan usus (%)*
Kontrol 5.53 ± 0.19 - 2B1 7.31 ± 0.21 24.18 ± 2.17 b 1A5 6.62 ± 0.19 15.90 ± 1.54 c 2B2 7.31 ± 0.36 24.08 ± 3.15 b 2B4 6.99 ± 0.28 20.88 ± 1.14 b 1B1 6.69 ± 0.05 16.85 ± 0.56 c 2D1 7.47 ± 0.44 25.62 ± 3.28 b 1C4 6.66 ± 0.33 16.35 ± 1.31 c 2C12 8.15 ± 0.31 31.57 ± 2.22 a 1A32 7.33 ± 0.24 23.87 ± 3.62 b 2C2 7.02 ± 0.26 20.94 ± 2.20 b * = huruf berbeda pada kolom yang sama menyatakan beda nyata (p<0.05)
Kemampuan penempelan kesepuluh isolat BAL pada permukaan usus
tikus bagian ileum secara in vitro berbeda nyata (p<0.05). Kemampuan
penempelan BAL sangat berarti jika BAL dikonsumsi sebagai pangan ataupun
suplemen fungsional, karena dengan adanya BAL yang menempel maka BAL
tersebut mampu bertahan hidup lebih lama di saluran pencernaan yang selanjutnya
dapat tumbuh, sedangkan BAL yang tidak mampu menempel dengan baik akan
ikut gerakan peristaltik usus bersama sisa makanan untuk selanjutnya terbuang
bersama feses. Implikasi selanjutnya adalah BAL yang mampu menempel di usus
akan mampu memberikan efek pertahanan usus yang lebih baik untuk menolak
adanya kemungkinan bakteri patogen yang mampu melakukan penempelan ke

72
usus. Dengan demikian, sifat penempelan yang lebih tinggi akan memberikan efek
menguntungkan yang lebih tinggi.
Isolat BAL 2C12 mampu menempel pada permukaan usus lebih baik
sebanyak 2.62 log cfu/cm2 atau sebesar 31.57% dibandingkan dengan isolat
lainnya. Sebanyak enam isolat (2B1, 2B2, 2B4, 2D1, 1A32 dan 2C2) memiliki
kemampuan penempelan pada permukaan usus yang tidak berbeda dengan kisaran
20.88-25.62%, sedangkan tiga isolat lainnya (1A5, 1B1 dan 1C4) memiliki
penempelan terendah dengan kisaran 15.90 – 16.85%. Nilai penempelan di atas
20% dinyatakan tinggi oleh Nitisinprasert et al. (2006). Berdasarkan kriteria
Nitisinprasert et al. (2006), sebanyak tujuh isolat BAL dinyatakan memiliki nilai
penempelan tinggi (>20%) yaitu 2C12, 2B1, 2B2, 2B4, 2D1, 1A32 dan 2C2,
sedangkan tiga isolat lainnya (1A5, 1B1 dan 1C4) mempunyai nilai penempelan
rendah
Hasil penelitian ini menunjukkan bahwa kemampuan penempelan BAL
pada permukaan usus dipengaruhi oleh spesies dan galur. Hasil penelitian ini
memperkuat pendapat Morelli (2007) yang menyatakan bahwa kemampuan BAL
pada sel epitel usus tergantung pada spesifik galur, dan tidak tergantung pada asal
BAL tersebut diisolasi. Penelitiannya membuktikan bahwa sembilan galur L.
acidophilus mempunyai kemampuan menempel yang berbeda pada sel Caco-2.
Sebanyak dua galur L. acidophilus yang diisolasi dari manusia menunjukkan
kemampuan menempel pada sel Caco-2 lebih baik daripada galur L. acidophilus
yang diisolasi dari ayam, namun terdapat juga lima galur L. acidophilus dari
manusia yang mempunyai nilai penempelan lebih rendah daripada L. acidophilus
yang diisolasi dari ayam (Morelli 2007). Mishra dan Prasad (2005) yang
melakukan penelitian penempelan BAL L. casei pada sel usus tikus juga
menemukan bahwa galur L. casei yang diisolasi dari susu mempunyai nilai
penempelan yang tinggi berkisar dari 52.8-66%. Nitisinprasert et al. (2006)
melaporkan bahwa penempelan L. reuteri KUB-AC5, AC-16, AC 20 berkisar
pada 0.87%-21.58%, sedangkan L. acidophilus KV-1 memiliki kemampuan
penempelan sebesar 23.1%.
Hasil penelitian ini juga memperkuat hasil penelitian yang dilakukan oleh
Pennacchia et al. (2006) yang menyatakan bahwa galur BAL mempengaruhi

73
kemampuan penempelan pada sel usus Caco-2 secara in vitro. Sebanyak 16 galur
Lactobacillus dengan populasi awal 109 cfu/ml diuji kemampuan penempelannya
pada sel Caco-2 dan hasilnya menunjukkan bahwa sebanyak enam galur L.
plantarum, satu galur L. paraplantarum dan satu galur L. pentosus mempunyai
tingkat penempelan yang tinggi yaitu sebesar ( 6.5 x 107 cfu), sedangkan dua
galur L. brevis mempunyai tingkat penempelan sedang (1.5 x 107 dan 2.1 x 107
cfu) dan satu galur L. paracasei mempunyai tingkat penempelan yang rendah (1.4
x 106 cfu).
Hasil penelitian ini juga sejalan dengan penelitian Lin et al (2006) yang
menunjukkan bahwa isolat BAL yang terdapat pada produk yogurt B mampu
menempel pada sel epitel usus manusia dengan populasi berkisar dari 105 – 106
cfu per sel Int-407, sedangkan isolat BAL pada produk A, C, D, dan E tidak ada
yang mampu menempel pada sel epitel Int-407. Setelah diidentifikasi secara
biokimiawi (API 50 CHL kit) dan molekuler (pita kromosom DNA dengan
metode pulsed field gel electrophoresis/ PFGE), isolat BAL yang terdapat pada
produk yogurt B adalah Lactobacilus acidophilus. Hal ini menunjukkan bahwa
setelah mampu melewati kondisi asam lambung dan garam empedu di saluran
pencernaan secara alami, L. acidophilus mampu menempel di mukosa usus
selanjutnya berproliferasi sehingga populasinya semakin meningkat.
Penempelan bakteri pada permukaan epitel usus dipengaruhi oleh sel
epitel usus, matriks ekstraseluler dan lapisan mukus. Lapisan mukus yang
menutupi sel epitel merupakan kontak pertama di usus bagi mikroorganisme
untuk melakukan penempelan dan kolonisasi di usus. Jika mukosa rusak, maka sel
epitel usus merupakan tempat penempelan bakteri. Permukaan sel epitel usus
diselimuti oleh lapisan yang bersifat visko-elastik yang terdiri dari glikoprotein
yang merupakan sisi tempat penempelan bakteri. Setelah bakteri berhasil
menempel dengan adanya interaksi antara adhesin dengan reseptor yang berupa
glikoprotein di sel epitel usus, maka bakteri tersebut akan berkolonisasi dan
memanfaatkan musin serta glikoprotein lainnya untuk pertumbuhannya
(Adlerberth et al. 2000).
Adhesin merupakan suatu protein yang mengenali glikoprotein membran
sel usus. Pada bakteri Gram positif, adhesin berada di dinding sel. L. plantarum

74
mengekspresikan manosa-binding adhesin yang lebih baik daripada spesies
Lactobacillus lainnya. Oleh karenanya L. plantarum cenderung memiliki sifat
penempelan ke permukaan usus yang baik (Adlerberth et al. 2000).
SIMPULAN
BAL indigenus asal daging sapi (28 isolat) menunjukkan ketahanan yang
berbeda terhadap pH 2.0, 2.5, 3.2 dan 7.2, namun hanya 10 isolat (2B4, 1B1, 1A5,
2C2, 2D1, 2B2, 1C4, 2B1, 1A32 dan 2C12) yang mampu bertahan pada pH 2.0
dengan ketahanan hidup di atas 50% ( lebih dari 105 cfu/ml), pada pH 2.5 ( di atas
70%) serta pH 3.2 dan 7.2 ( di atas 80%). Kesepuluh isolat BAL juga mampu
tahan garam empedu 0.5% dengan baik ( di atas 70%), mempunyai aktivitas
antimikroba dan koagregrasi yang baik terhadap bakteri patogen E. coli ATCC
25922, S. Typhimurium ATCC 14028, EPEC dan S. aureus ATCC 25923
(diameter penghambatan lebih dari 6mm, kecuali isolat 2B1 terhadap EPEC;
koagregasi lebih dari 20% kecuali isolat 1A5 terhadap EPEC). Kesepuluh isolat
BAL mampu menempel pada permukaan usus sebesar 15.90-31.57%. Dengan
demikian, 10 isolat BAL indigenus yang diisolasi dari daging sapi lokal
mempunyai sifat sebagai kandidat probiotik.
DAFTAR PUSTAKA
Adlerberth I, Cerquetti M, Poilane I, Wold A, Collignon A. 2000. Mechanisms of
colonization and resistance of the digestive tract. Part 1: bacteria/host interactions. Microbial Ecology in Health and Disease. 2 : 223-239.
Albano H et al. 2007. Characterization of two bacteriocins produced by Pediococcus acidilactici isolated from ‘Alheira’ a fermented sausage traditionally produced in Portugal. Int J of Food Microbiol 116 : 239-247.
Arief II, Maheswari RRA, Suryati T. 2007. Karakteristik dan Nilai Gizi Protein Daging Sapi Dark Firm Dry (DFD) yang Difermentasi oleh Lactobacillus plantarum yang Diisolasi dari Daging Sapi. Laporan Penelitian Hibah Bersaing XIII. LPPM-IPB.

75
Axelsson L. 1993. Lactid Acid Bacteria : classification and physiology. Di dalam: Lactid Acid Bacteria: Microbiology and Functional Aspects. 2nd Edition, Revised and Expanded. Salminen, S., and von Wright, A. (Editors.). New York : Marcell Dekker Inc.
Bao Y et al. 2010. Screening of potential probiotics properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control 21 : 695-708.
Bogaert JC, Naidu AS. 2000. Lactic acid. Di dalam: Natural Food Antimicrobial System. AS. Naidu (editor). Florida : CRC Press.
Begley M, Hill C, Grahan CGM. 2006. Bile salt hydrolase activity in probiotics. Appl & Environ Microbiol 72: 1729-1738
Blum S et al. 1999. Adhesion studies for probiotics : need for validation and refinement. Trends in Food Sci & Technol 10 : 405-410.
Bourlioux P, Koletzko B, Guarner F, Braesco V. 2003. The intestine and its microflora are partners for the protection of the host : report on the Danone symposium ‘ The Intelligent Intestine’, held in Paris, June 14, 2002. Am J Clin Nutr 78: 675-683.
Bron PA et al. 2004. Genetic characterization of the bile salt response in Lactobacillus plantarum and analysis of responsive promoters in vitro and in situ in the gastrointestinal tract. J Bacteriol 186: 7829-7835
Cotter PD, Hill C. 2003. Surviving the acid test : responses of Gram-positive bacteria to low pH. Microbiol Molecular Biol Rev 67 (3): 429-453.
El-Naggar MYM. 2004. Comparative study of probiotic cultures to control the growth of Escherichia coli O157:H7 and Salmonella typhimurium. Biotechnol 3: 173-180.
FAO/ WHO. 2002. Guidelines for the evaluation of probiotics in food. Report of Joint FAO/WHO Working Group on drafting Guidelines for the evaluation of probiotics in food. London Ontario, Canada.
Fitrial Y. 2009. Analisis potensi biji dan umbi teratai (Nymphaea pubescens wild) untuk pangan fungsional prebiotik dan antibakteri Escherichia coli enteropatogenik K.1.1. [Disertasi] Sekolah Pascasarjana Institut Pertanian Bogor.
Gbassi GK, Vandamme T, Yolou FS, Marchioni E. 2011. In vitro effects on pH, bile salts and enzymes on the release and viability of encapsulated Lactobacillus plantarum strains in a gastrointestinal tract model. Int Dairy J 21: 97-102.
Herich R, Levkut M. 2002. Lactic acid bacteria, probiotics and immune system. Vet Med-Czech 47 :169-180.

76
Kimoto-Nira H et al. 2007. Lactococcus sp. as potential probiotic lactic acid bacteria. JARQ 41: 181-189.
Kos B et al. 2003. Adhesion and agggregration ability of probiotic strain Lactobacillus acidophilus M92. J Appl Microbiol 94: 981-987.
Lin WH, Hwang CF, Chen LW, Tsen HY. 2006. Viable counts, characteristic evaluation for commercial lactic acid bacteria products. Food Microbiol 23: 78-81.
Miksusanti. 2009. Aktivitas dan mekanisme antibakteri minyak atsiri temu kunci (Kaempferia pandurata roxb) serta inkorporasinya dalam pati sagu sebagai film edibel antibakteri. [Disertasi]. Sekolah Pascasarjana Institut Pertanian Bogor.
Mishra V, Prasad DN. 2005. Application of in vitro methods for selection of Lactobacillus casei strains as potential probiotics. Int J Food Microbiol 103 : 109-115.
Moll GN, Konnings WN, Driessen AJM. 1999. Bacteriocins: mechanism of membrane insertion and pore formation. Antonie van Leeuwenhoek 76: 185-198.
Morelli L. 2007. In vitro assessment of probiotic bacteria : from survival to functionally. Int Dairy Journal 17: 1278-1283.
Moser SA, Savage DC. 2001. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in Lactobacilli. Appl & Environ Microbiol 67: 3476-3480.
Mourad K, Eddine KN. 2006. In vitro preselection criteria for probiotic Lactobacillus plantarum of fermented olives origin. Int J Probiotics and Prebiotic 1 : 27-32.
Mourad K, Meriem KH. 2008. Probiotics characteristics of Lactobacillus plantarum strains from traditional butter made from camel milk in arid region (Sahara) of Algeria. Grasas Y Aceites 59 : 218-224.
Moyano SR, Martin A, Benito MJ, Nevado FP, Cordoba MGG. 2008. Screening of lactic acid bacteria and bifidobacteria for potential probiotic use in Iberian dry fermented sausages. Meat Sci 80: 715-721.
Naidu AS, Clemens RA. 2000. Probiotics. Di dalam : Natural Food Antimicrobial Systems. A.S. Naidu (editor). Florida : CRC Press.
Nitisinprasert S, Pungsungworn N, Wanchaitanawong P, Loiseau W, Montet D. 2006. In vitro adhesion assay of lactic acid bacteria, Escherichia coli and Salmonella sp. by microbiological and PCR methods. Songklanakarin J Sci Technol 28 (suppl.1) : 99-106.
Nowroozi J, Mirzaii M, Norouzi M. 2004. Studi of Lactobacillus as Probiotic Bacteria. Iranian J Publ Health 33 : 1-7.

77
Ogunbawo ST, Sanni AI, Onilude AA. 2003. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr J Biotechnol 2 :219-227
Ouwehand AC, Tuomola EM, Tolkko S, Salminen S. 2001. Assessment of adhesion properties of novel probiotics strains to human intestinal mucus. Int J Food Microbiol 64 : 119-126
Pal V, Jamuna M, Jeevaratnam K. 2005. Isolation and Characterization of Bacteriocin Producing Lactic Acid Bacteria from a South Indian Special Dosa (APPAM) Batter. J Culture Collection 4: 53-60
Pan X, Chen F, Wu T, Tang H, Zhao Z. 2009. The acid, bile tolerance and antimicrobial property of Lactobacillus acidophilus NIT. Food Control 20 : 598-602.
Pennachia C, Vaughan EE, Villani F. 2006. Potential probiotic Lactobacillus galurs from fermented sausages: Further investigations on their probiotic properties. Meat Sci 73: 90-101.
Pereira DIA, McCartey AL, Gibson GR. 2003. An in vitro study of the probiotic potential of a bile-salt-hydrolyzing Lactobacillus fermentum strain, and determination of its cholesterol-lowering properties. Appl & Environ Microbiol. 69: 4743-4752.
Rinkinen M, Jalava K, Westermarck E, Salminen S, Ouwehand AC. 2003. Interaction between probiotic lactic acid bacteria and canine enteric pathogens: a risk factor for intestinal Enterococcus faecium colonization?. Vet Microbiol 92: 111-119.
Roberfroid MB. 2000. Prebiotics and probiotics: are they functional foods?. Am Clin Nutr 71(suppl) : 1682S-1687S.
Salminen S, Wright AV. 2004. Lactic Acid Bacteria. Microbiology and Functional Aspects. 2nd Edition, Revised and Expanded. New York : Marcell Dekker, Inc.,
Savadogo A, Outtara CAT, Bassole IHN, Traore AS. 2006. Bacteriocins and lactic acid bacteria – a minireview. Afr J Biotechnol 5 (9): 678-683.
Savadogo A, Outtara CAT, Bassole IHN, Traore AS. 2004. Antimicrobial activities of lactic acid bacteria strains isolated from Burkina Faso fermented milk. Pak J Nutr 3: 174-179.
Steel RGD, Torrie JH. 1995. Principles and Procedures of Statistic a Biomedical Approach, 3rd Edition. McGraw Hill,Inc. Singapore.
Suarsana IN, Utama IH, Suartini NGAA. 2001. Aktivitas in vitro senyawa antimikroba dari Streptococcus lactis. J Vet 2 : 25-31.

78
Sunny-Roberts EO, Knoor D. 2008. Evaluation of the response of Lactobacillus rhamnosus VTT E-97800 to sucrose-induced osmotic stress. Food Microbiol 25 : 183-189.
Surono IS. 2004. Probiotik, Susu Fermentasi dan Kesehatan. YAPPMI. Jakarta.
Toksoy A, Beyatli Y, Aslim B. 1999. Study on metabolic and antimicrobial activities of some L. plantarum strains isolated from sausages. Tr J Vet An Sci 533-540.
Tuomola E, Crittenden R, Playne M, Isolauri E, Salminen S. 2001. Quality
assurance criteria for probiotic bacteria. Am J Clin Nutr 73 (suppl): 393S-398S.