3embryologie aus sicht der craniosacralen osteopathie · embryologie aus sicht der craniosacralen...
TRANSCRIPT

Embryologie aus Sicht der Craniosacralen Osteopathie 1/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
EMBRYOLOGIE aus Sicht der
CRANIOSACRALEN OSTEOPATHIE
Diplomarbeit
von
Rosmarie Walthert
Schule für Craniosacrale Osteopathie, Rudolf Merkel, Zürich

Embryologie aus Sicht der Craniosacralen Osteopathie 2/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Inhaltsverzeichnis Inhaltsverzeichnis....................................................................................................................... 2
I Zusammenfassung.......................................................................................................... 4
II Einführung...................................................................................................................... 5
III Meine Frage: Wie manifestiert sich der Primäre Atemmechanismus in der embryonalen Entwicklung des Menschen? .................................................................... 7
IV Der Primäre Atemmechanismus..................................................................................... 8
IV 1. Zum Begriff des Primären Atemmechanismus (PAM)............................................... 8 IV 2. Die fünf Merkmale des Primär Respiratorischen Atemmechanismus ........................ 9 IV 3. Die Fluktuation der Zerebrospinalen Flüssigkeit ........................................................ 9 IV 4. Die Reziproke Spannungsmembran .......................................................................... 10 IV 5. Die Motilität der Neuralröhre.................................................................................... 12 IV 6. Die Mobilität der Schädelknochen............................................................................ 13 IV 7. Die unwillkürliche Mobilität des Sakrums zwischen den Beckenknochen .............. 13 IV 8. Die Funktion des Primären Atemmechanismus ........................................................ 13 IV 9. Der Rhythmus ........................................................................................................... 14 IV 10. Erklärungsmodelle für den Primären Atemmechanismus......................................... 14
V Embryologie ................................................................................................................. 16
V 1. Die Bedeutung der Embryologie für die Praxis ........................................................ 16 V 2. Geschichtlicher Überblick über die Untersuchung der Humanembryologie ............ 17 V 3. Der Mensch als Embryo zwischen Himmel und Erde .............................................. 19 V 4. Der Embryo – eine Dokumentation der kinetischen Anatomie ................................ 21 V 5. Die Forschung ........................................................................................................... 21 V 6. Die Kinetische Entwicklungstheorie......................................................................... 22 V 6.1. Materialbewegungen und Stoffwechselfelder ........................................................... 22 V 6.2. Grenzgewebe und Binnengewebe ............................................................................. 23 V 6.2.1. Grenzgewebe....................................................................................................... 24 V 6.2.2. Keilepithelien ...................................................................................................... 26 V 6.2.3. Die Bedeutung der Flüssigkeitsräume................................................................. 27 V 6.2.4. Binnengewebe ..................................................................................................... 28 V 6.2.5. Die acht Stoffwechselfelder ................................................................................ 28 V 6.2.6. Kinetische Entwicklungstheorie und der Primäre Atemmechanismus ............... 32
VI Die embryonale Entwicklung des Nervensystems ....................................................... 34
VI 1. Erste Leistungen des Nervensystems sind Wachstumsleistungen ............................ 34 VI 2. Die Bildung des Neuralrohrs..................................................................................... 34 VI 2.1. Wachstumsleistungen des Neuralrohrs ............................................................... 35 VI 2.2. Wachstumsleistungen von Gehirn und Rückenmark .......................................... 36 VI 2.3. Gliederung des Nervensystems ........................................................................... 39
VII Der Primäre Atemmechanismus und seine Entstehung in der embryonalen Entwicklungsphase....................................................................................................... 41
VII 1. Gestaltende Energie oder „breath of life“ ................................................................. 41 VII 1.1. Anfänge des energetischen Aspektes .................................................................. 42

Embryologie aus Sicht der Craniosacralen Osteopathie 3/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 1.2. Primitivknoten als Organisationszentrum ........................................................... 43 VII 1.3. Die ventrale Mittellinie oder Chorda dorsalis ..................................................... 44 VII 1.4. Die Bildung der Somiten..................................................................................... 45 VII 1.5. Die dorsale Mittellinie oder Zentralkanal des Neuralrohrs ................................. 46 VII 1.6. Abschliessende Gedanken zum energetischen Aspekt........................................ 47 VII 2. Gestaltende Bewegung.............................................................................................. 51 VII 2.1. Blutbildung und Herzkreislauf ............................................................................ 52 VII 2.2. Die Gestaltungsfunktion des Gefässsystems....................................................... 53 VII 2.3. Die Ausgestaltung der Körpergrundgestalt ......................................................... 56 VII 2.3.1. Die laterale Abfaltung ......................................................................................... 57 VII 2.3.2. Die kraniokaudale Krümmung ............................................................................ 58 VII 2.3.3. Embryonales Üben des Primären Atemmechanismus (PAM) ............................ 59 VII 2.3.4. Embryonales Üben der sekundären Lungenatmung............................................ 60 VII 3. Parallelen und Unterschiede von Primärem und Sekundärem Atemmechanismus. . 62 VII 3.1. Wechselwirkungen der Gewebeschichten........................................................... 62 VII 3.2. Relevanz für meine praktische Arbeit. ................................................................ 62
Bildnachweis: ........................................................................................................................... 64
Literaturangabe: ....................................................................................................................... 65

Embryologie aus Sicht der Craniosacralen Osteopathie 4/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
I Zusammenfassung William Garner Sutherland hat seinem Konzept der cranialen Arbeit in der Osteo-pathie den Primären Atemmechanismus als inhärente Bewegung von Gehirn und Rückenmark zugrunde gelegt. Diese Bewegung ist als subtile Bewegung mit ent-sprechend sensibilisierter Palpationsfähigkeit an den beweglichen Schädelknochen und als unwillkürliche Bewegung am Sakrum und dank der Verbindung zu den Kör-perfaszien auch im übrigen Körper wahrnehmbar. Sutherland hat den Ursprung dieses Mechanismus, entsprechend seiner Funktion (Pumpe für das venöse Gefässsystem im Schädelbereich, Homöostase im ganzen Körper) in der embryonalen Phase angelegt. Bis heute bleibt dies Gegenstand von Diskussionen und Untersuchungen. Mit der vorliegenden Arbeit hoffe ich, etwas zu dieser Diskussion beitragen zu kön-nen. Das biodynamische Modell der Osteopathie (James S. Jealous, DO) und der cranio-sacralen Osteopathie (Franklin Sills) spricht von zwei Mittellinien, einer ventralen (An-lage der Chorda dorsalis) und einer dorsalen (Zentralkanal). Beide Mittellinien wer-den mit dem wandernden Chordafortsatz und Axialkanal induziert. Zuerst entsteht die Chorda dorsalis (späterer Nucleus pulposus). Wenig später faltet sich die Neuralplat-te auf und schliesst sich zum Neuralrohr. Der Zentralkanal in seiner Mitte wird die dorsale Mittellinie. Beide Linien stehen für ein feinstoffliches Energieprinzip, das für die Aufrechterhaltung des Organismus während des ganzen Lebens sorgt. In fernöst-lichen jahrtausende alten medizinischen Traditionen findet man dieses Prinzip in Form von zum Beispiel Chi in China oder Prana in Indien. In der vorliegenden Arbeit stelle ich die These auf, dass der Ursprung dieser Bewe-gung in der einzigen Krümmungsphase des Embryos liegt. Für diese Krümmung ist das ungeheure Wachstum des Neuralrohrs verantwortlich, das sich in unmittelbarer Zusammenarbeit mit der Herz-/Gefässanlage (Blechschmidt) entwickelt. In dieser Phase geschieht die Organogenese schlechthin. Damit verbunden ist die Vernetzung des wachsenden Nervensystems mit den Organlagen des ganzen Körpers. Die Embryologen Jaap van der Wal und Erich Blechschmidt haben mir wichtige In-strumente für meine Schlussfolgerungen geliefert: Von Jaap van der Wal habe ich das philosophische Empfindungskonzept der „partizipierenden Phänomenologie“ und von Erich Blechschmidt die Untersuchungsergebnisse seiner Forschungsarbeiten zur „kinetischen Entwicklungstheorie mit einer raum-zeitlich beschreibenden Gestal-tungsanatomie“ kennengelernt. Der Embryo übt seine körperlichen und seelischen Funktionen voraus. Vor dem Hintergrund dieser Aussage bin ich zu folgendem Schluss gekommen: der Embryo übt während der Entwicklungs-Gebärde der Krümmung mit der Wachstums-tätigkeit des Neuralrohrs eine alternierende Bewegung gegen den Widerstand der Herz-/Gefässanlage aus. Es ist das Neuralrohr, das den Bewegungsimpuls setzt (Flexionsphase). Das Gefässsystem wirkt wachstumshemmend (Extensionsphase) bis es die nötige Sauerstoffversorgung wieder erbringt. Aus dieser Wachstumszu-sammenarbeit bleiben später die inhärente zyklische Flexionsbewegung (nach mei-ner Erfahrung stets deutlicher palpierbar), gefolgt von der (schwächer palpierbaren, manchmal kürzer dauernden) Extensionsbewegung des Primären Atemmechanis-mus weiter bestehen.

Embryologie aus Sicht der Craniosacralen Osteopathie 5/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
II Einführung Am Anfang empfand ich die Embryonalentwicklung, wie sie in der Literatur als Be-schreibung der bewegungslosen materiellen Ebene einer einzelnen Momentanauf-nahme dargestellt wird als Stückwerk. Es schien mir kaum möglich, in irgendeiner Form einen Bogen schlagen zu können zum sogenannten Primären Atemmechanis-mus. Der Primäre Atemmechanismus oder „Breath of life“ ist das Konzept wie es Dr. William Garner Sutherland, Osteopath (1873 -1954), für die Wissenschaft der krania-len Osteopathie geschaffen hat (Kapitel III). Der Sprung zwischen der subtil wahr-nehmbaren Eigenbewegung der Gewebe während meiner Arbeit zum extrem detail-lierten Stückwerk der anatomischen Fachsprache der Embryologen, die Zustände an der leblosen Materie beschreiben, erschien mir unüberwindbar. Eine erste grosse Hilfe und Unterstützung war das Seminar „Der Mensch als Embryo zwischen Himmel und Erde“ mit Jaap van der Wal, Arzt, Embryologe, Philosoph, Pri-vatdozent an der Universität Maastricht. Kapitel IV, 3. beschreibt seine Auffassung vom Embryo als Wesen, das sich durch eine ständige Metamorphose seiner Form verhält. Damit ist gemeint, dass sich das Erscheinungsbild im Zeitverlauf verändern kann, das eigentliche Wesen aber unverändert gegenwärtig und aktiv innerhalb die-ser äusseren Gestalten und Formen bleibt. Sein Plädoyer für die Methode der parti-zipierenden Phänomenologie im Hinblick auf das Verständnis des embryonalen Menschen angewandt, bewegt mich sehr. Mit der Haltung einer teilnehmenden Wahrnehmung der Erscheinungswelt gewinnen die rasch sich verändernden Formen und Gewebe in der menschlichen embryonalen Entwicklung an Lebendigkeit und an Sinn. Jaap van der Wal vermag bald schon das Werden und Wachsen von Keim-schichten, Geweben, Formen und Gestalt in Bewegung innerlich vorstellbar und nachvollziehbar werden zu lassen und setzt es in Zusammenhang mit der individuel-len Entwicklung des Menschen - seiner Individuation grundsätzlich. Eine unglaubli-che Kraft geht von den Embryonen in ihrer aktiven Entwicklungstätigkeit aus. Ein weiteres prägendes und inspirierendes Erlebnis war der Besuch im anatomi-schen Institut der Universität Göttingen, wo ich die Blechschmidt-Sammlung der To-talrekonstruktionen von Embryonen besuchte. Herr Prof. Dr. med. Erich Blechschmidt (1902-1992), von 1942 bis 1973 Direktor des Anatomischen Instituts der Universität Göttingen, stellte sich in umfangreichen Forschungsarbeiten die Fra-ge, inwieweit eine fruchtbare Synthese von der Leichenanatomie zur Physiologie möglich ist. Mit dem Studium seines Lebenswerks, seiner Bücher, zahlreicher Veröf-fentlichungen und der Dokumentation zur Kinetischen Anatomie (Kapitel IV, 4.) des menschlichen Embryos eröffnete sich mir eine plastische und sprachliche Aus-drucksweise der Morphologie als lebendigem Werdeprozess – in kontinuierlicher Gestalt annehmender Bewegung, die mich sehr berührt. Erich Blechschmidt begrün-dete die kinetische Entwicklungstheorie (Kapitel V, 6.). Der Aspekt der Bewegung als Entwicklungsbewegung mit der funktionellen Bedeutung der Gestaltungsfunktion als Grundfunktion der Organe wirkt extrem elektrisierend auf mich. Ein ungeheurer Wis-sensdurst wird in mir genährt. Die Herren Blechschmidt und van der Wal sind beide der gleichen Ansicht, dass die Gene allein nicht genügen für die Entwicklung. Beide beschreiben sie den Entwick-

Embryologie aus Sicht der Craniosacralen Osteopathie 6/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
lungsreiz als von ausserhalb in die Zelle eintretend. Die Gene leiten daraufhin den nächsten Entwicklungsschritt ein. Ohne Gene fehlen die zentralen Angriffspunkte für die Entwicklungsreize und ohne Entwicklungsreize bleiben Gene bedeutungslos. Im Wechselspiel zwischen unspezifischem Reiz von aussen und spezifischer Reaktion im Zellinneren regulieren die hochmolekularen Gene Reizwirkungen, die sonst Stör-momente bedeuten würden. Die Kompensationen äussern sich als Entwicklungs-schritte. Blechschmidt nimmt als Grundgeschehen von diesen Vorgängen einen in-trazellulären Kreislauf von Stoffwechselbewegungen in Richtung von der Zellgrenz-membran zum Zellkern und von da wieder zurück zur Zellgrenzmembran an. Diesen Kreislauf sieht er als eine Voraussetzung für die Beziehung zwischen den Anlagen im Inneren der Zellen und ihrem äusseren Erscheinungsbild. Entgegen der üblichen Anschauung, wonach ein Embryo gerade noch nicht mensch-lich ist, weil noch nicht alle Organe angelegt sind oder das anatomische Hirnsubstrat gerade noch nicht ausreichend gereift ist, bejahen beide den Embryo als menschli-chen Organismus vom ersten Tag an - aktiv arbeitend am Akt der Differenzierung „aus seiner Ganzheit heraus“ (Jaap van der Wal) und „wachsend von Anfang an durch menschlich spezifische Stoffwechselvorgänge, die der Embryo gegen Wider-stand, also durch Arbeit leistet“ (E. Blechschmidt). Beide lehnen die darwinsche Evo-lutionstheorie ab - Blechschmidt mit der Begründung, Darwin habe nicht „exakt ge-nug geforscht“?.

Embryologie aus Sicht der Craniosacralen Osteopathie 7/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
III Meine Frage: Wie manifestiert sich der Primäre Atemmechanismus in der embryonalen Entwicklung des Menschen?
Die Idee zu dieser Fragestellung hat in Jaap van der Wals Seminar Bestärkung er-fahren. Die partizipierenden Phänomenologie anwendend habe ich mir gedacht: Wenn der Mensch in der embryonalen Phase seine seelischen und seine körperli-chen Funktionen vorübt, dann muss es für einen physiologischen Vorgang, wie den Primären Atemmechanismus auch eine frühe Vorform als Wachstumsgebärde oder eine Entwicklungsphase geben. Um eine mögliche Antwort zu finden, musste ich mich viel tiefer greifend mit der Hu-manembryologie auseinandersetzen und erst einmal viel lernen. Weil das immer spannender und interessanter wurde, sind die Ausführungen zur embryologischen Entwicklung aus verschiedenen Blickwinkeln recht umfangreich geworden. In Blechschmidt’s Forschungsaufzeichnungen hat sich meine Frage als berechtige These erwiesen. Er spricht von den Wachstumsfunktionen der Zellen und Organen als deren Grundfunktion, die als frühe Tätigkeit – Gestaltungsleistungen - die späte-ren organspezifischen Leistungen einleiten. Warum also sollte das Zusammenwirken von Gehirn/Liquor/cranialem und spinalem Membransystem und ihre inhärente Be-wegung mit den beweglichen Schädelknochen nicht schon in diesem Werdeprozess anlagemässig gefunden werden können? Ich gehe der Forschungsarbeit von Erich Blechschmidt so ausführlich nach, wie ich es für mein inneres Vorstellungsvermögen brauche, bis Werden, Wachsen von Zel-len, Geweben und Stoffwechselabläufen innerlich zusammenhängend verständlich und lebendig werden. Meine Frage hat ihre Wurzeln in meiner alltäglichen, beruflichen Tätigkeit. Sie ist verbunden mit Palpation von subtilen Bewegungsvorgängen, hat also mit Sinnes-wahrnehmung zu tun. Ich nähere mich Blechschmidt’s Forschungsergebnissen in Schriftsprache ausgedrückt daher in einer eher empfindenden Art und Weise, eher im Sinn von innerlichem Ertasten und Abtasten seiner Untersuchungen und Gedan-kengängen. Innerlich abwägend suche ich sie nach Aufschluss, Illustration oder auch Erklärung zu Ursprung und Beginn dieses physiologischen Vorganges des Primären Atemmechanismus ab. Weitere Grundlagen bieten mir die „Funktionelle Embryologie“ und die „Morphologie des menschlichen Organismus“ von Prof. (em.) Dr. med. Johannes W. Rohen, Ana-tomisches Institut, Universität Erlangen-Nürnberg sowie Bücher von Prof. Dr. med. Ulrich Drews, Universität Tübingen, Prof. Dr. med. Thomas W. Sadler, Direktor für Zellbiologie und Anatomie an der University of North Carolina, USA, und Prof. Dr. med. Ronan O’Rahilly, Institut für Spezielle Embryologie, Universität Fribourg, Schweiz. Weitere Literaturangaben finden sich im Verzeichnis im Anhang.

Embryologie aus Sicht der Craniosacralen Osteopathie 8/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
IV Der Primäre Atemmechanismus Dr. William Garner Sutherland, Doctor of Osteopathy (D. O.), 1873 – 1954 hat an-hand langjähriger Studien erforscht, dass den Schädelknochen eine spezifische Ei-genbewegung bis ins hohe Alter erhalten bleibt. „Die Annahme, dass der Schädel ei-ne rigide Struktur ist, entspricht der Schlussfolgerung aus trockenen, entfetteten Kör-pern aus dem Labor. Bei Forschungsexperimenten an lebenden Strukturen ist die physiologische Präsenz einer normalen, wenn auch minimalen Bewegung oder ihre pathologische Restriktion ziemlich offensichtlich.“ (Magoun, Osteopathie in der Schädelsphäre).
Der Primäre Atemmechanismus (PAM) Cranio-rhytmyc-impuls (CRI) Der Primäre Atemmechanismus (PAM) ist der bildliche Ausdruck, den William G. Sutherland seinem cranialen Konzept in der Wissenschaft der Osteopathie zugrunde gelegt hat. Sein Leben war gänzlich der Forschung (Beobachtungen und Experimen-ten am lebenden Menschen - an sich selbst und später auch an Patienten) gewid-met. 1939 trat er mit einem kleinen Buch „The cranial Bowl“ erstmals an die Öffent-lichkeit. Das Buch ist eine wichtige Grundlage für Osteopathen, Therapeuten und Wissenschafter. Darin ist vom Schädel als einem membranösen Gelenkmechanis-mus die Rede, welcher die Existenz einer simultan stattfindenden Bewegung in den Schädelsuturen und im Kreuzbein beschreibt. Schädel und Kreuzbein sind verbun-den durch die kraniale und die spinale Dura. Eine muskuläre Ursache für eine derar-tige Bewegungssynchronisation existiert nicht – die Quelle oder der Motor muss also woanders gesucht werden, was bis heute Gegenstand von Untersuchungen und Dis-kussionen ist. Der Cranio-rhythmic-Impuls (CRI) sollte ursprünglich unabhängig von den Erklä-rungsversuchen dieses Rhythmus nur die messbare, physiologische unwillkürliche und rhythmische Expansions- und Retraktionsbewegung am Schädel, insbesondere am Asterion (Verbindungsstelle von Scheitel-, Schläfen- und Hinterhauptsbein) um-schreiben. Der Begriff wurde von den Osteopathen und Psychiatern Woods, J. M. und Woods, R. H. geprägt, damit andere Ärzte diese Bewegung palpieren und be-werten konnten, ohne mit der Idee des Primär Respiratorischen Mechanismus kon-frontiert zu werden.
IV 1. Zum Begriff des Primären Atemmechanismus (PAM) (Liem, Torsten, Kraniosakrale Osteopathie) Anstelle von Primärem Atemmechanismus wird häufig auch der Begriff Primärer re-spiratorische Mechanismus (PRM) gebraucht. Sie meinen beide das Gleiche. Der Mechanismus wird „primär“ genannt, weil er direkt mit der inneren Gewebeat-mung des Zentralnervensystems, das die Lungenatmung und die gesamte Körper-funktionen reguliert, verbunden ist. Dabei ist die Anwesenheit von physiologischen Zentren am Boden des 4. Ventrikels, die für die Lebensprozesse notwendig sind, für Sutherland’s Einschätzung von primärer Bedeutung für den gesamten Mechanismus. Ausserdem tritt der Primäre Atemmechanismus nach Angabe einiger Osteopathen bereits ungefähr im 5. Fetalmonat auf und ist noch Minuten bis Stunden nach dem Tod wahrnehmbar. (Liem, Torsten)

Embryologie aus Sicht der Craniosacralen Osteopathie 9/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Sutherland selber bezeichnet im Gegensatz dazu die Lungenatmung als sekundär respiratorisches System, da diese durch die primäre Atmung kontrolliert wird. Der Mechanismus wird „respiratorisch“ genannt, weil er einen rhythmischen Vor-gang darstellt, der mit Austauschprozessen zu tun hat. Er stellt einen Stoffwechsel dar, der sich zuerst intrakranial abspielt und in Verbindung steht mit dem Nervensys-tem und dem Liquor cerebrospinalis (LCS). Aber durch die rhythmische Drainage der gesamten Körpergewebe spielt er auch eine bedeutende Rolle bei der Gewebeat-mung des gesamten Organismus. Die Gewebeatmung des Nervensystems wie des übrigen Körpers verläuft autonom und unwillkürlich. „Mechanismus“ wird er genannt, da er aus Teilen besteht, die zusammen den Me-chanismus oder Motor für den CRI bilden. Der Mechanismus betrifft den ganzen Kör-per als physiologische Funktionseinheit. Sutherland beschreibt fünf Phänomene, die ihm zugehörig sind.
IV 2. Die fünf Merkmale des Primär Respiratorischen Atemmechanismus (Hartmann, Christian, Das grosse Sutherland-Kompendium, Unterweisung der Wissenschaft der Osteopathie)
1. Die Fluktuation der zerebrospinalen Flüssigkeit (Liquor cerebrospinalis, LCS)
2. Die „Funktion der Reziproken Spannungsmembran“, eine Mobilität (Beweg-lichkeit) der intrakranialen und intraspinalen Membranen
3. Die „Motilität des Neuralrohrs“, eine angeborene Motilität (inhärente Bewe-gung) von Gehirn und Rückenmark
4. Die Mobilität, Beweglichkeit der Schädelknochen 5. Die unwillkürliche Mobilität (Beweglichkeit) des Kreuzbeines zwischen den
Beckenknochen
IV 3. Die Fluktuation der Zerebrospinalen Flüssigkeit Sutherland bezeichnet ein unsichtbares Element innerhalb der Zerebrospinalen Flüs-sigkeit als den Atem des Lebens – eine Art Flüssigkeit innerhalb der Flüssigkeit, et-was, das sich nicht vermischt. Er vergleicht es mit den Meeresgezeiten – ein Herein-kommen und Hinausgehen. Die Fluktuation der Tide ist eine Bewegung, die durch Inhalation anflutet und durch Exhalation verebbt. Nicht die Wellen, welche der Küste entlangrollen, sind die Gezeiten. Die Bewegung der Tide ist die Bewegung des ge-samten Wasserkörpers namens Ozean. Die Kraft in der Tide nennt Sutherland Po-tency, Intelligenz in den Gezeiten. Er wollte die Potency, die Intelligenz in der Fluktu-ation der zerebrospinalen Flüssigkeit verstanden wissen. „Und dass der Atem des Lebens in diese Gestalt aus Lehm geblasen und der Mensch so zu einer lebendigen Seele wurde“. Ein Punkt in der Mitte zwischen Inhalation und Exhalation – ein Balan-cepunkt im Zentrum der Tide wollte er seinen Studenten verständlich machen als „The Still Small Voice“. Mit Verstehen dieser kleinen ruhigen Stimme sollte es mög-lich werden, die Reziproke Spannungsmembran an den Balancepunkt hinzuführen. Das Ventrikelsystem mit dem Liquor cerebrospinalis (LCS) ist besonders schwierig zu untersuchen, denn beim Öffnen werden die hydrodynamischen Bedingungen, un-ter denen es normalerweise existiert, verändert. Trotzdem gibt es nach Liem (Kraniosakrale Osteopathie) Anhaltspunkte, die dafür sprechen, dass die Flüssigkeit nicht oder nicht nur zirkuliert, sondern vielmehr fluktuiert. Die Fluktuation kommt durch die rhythmische Füllungs- und Entleerungsphase der

Embryologie aus Sicht der Craniosacralen Osteopathie 10/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Ventrikel zustande. Liem beschreibt weiter, dass sich die Fluktuation nicht nur in Ge-hirn und Rückenmark ausbreitet, sondern durch die Mikrotubuli in den Körperfaszien im gesamten Körper. Die Mikrotubuli sind hohle Kollagenfasern mit einem Durch-messer von ca. 0.5 µ. Die intrakranialen und intraspinalen Membranen setzen sich an den Nervenaustrittsstellen des Schädels und der Wirbelsäule in den Nervenschei-den der austretenden Nerven fort. Dabei gelangt auch LCS entlang dieser Nerven-scheiden in das extrakraniale System. Von den Nervenscheiden kommt der LCS ü-ber diese Mikrotubuli mit den extrazellulären Räumen und sogar der Lymphe in Kon-takt.
IV 4. Die Reziproke Spannungsmembran Mangels einer muskulären Unterstützung für die Bewegung zwischen den Schädel-knochen an den Suturen muss die Erklärung für diese Gelenkbewegung anderweitig gesucht werden. Die knochenverbindende Struktur der einzelnen Schädelknochen untereinander ist die intrakraniale Dura. Sutherland spricht von ihr einerseits unter dem Gesichtspunkt der kontinuierlichen Spannung und andererseits aus dem Blick-winkel des Fulkrum. Die Funktion der beiden zusammen stellt das zweite Prinzip des primären Atemmechanismus dar. Abb. 1 Kraniale Reziproke Spannungsmembran

Embryologie aus Sicht der Craniosacralen Osteopathie 11/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Laboratorische Dissektion geben Falx cerebri und Tentorium cerebelli die Bedeutung der Grosshirnabstützung, der Abgrenzung vom Kleinhirn und der zusätzlichen Auf-gabe der Schockabsorption und Stressverarbeitung. In ihrer lebendigen Physiologie sollten die cranialen Membranen darüber hinaus als drei untereinander verbundene, sichelförmige Strukturen betrachtet werden. Um das Gleichgewicht der Membranbe-wegung und –spannung in allen Richtungen zu gewährleisten, müssen die Membra-nen von einem Fulkrum, einem Ruhepunkt aus operieren. Dieser Ruhepunkt muss schwebend aufgehängt sein, um sich automatisch bewegen zu können, damit eine gleichmässige physiologische Bewegung der Schädelknochen gesichert ist, wenn Veränderungen auftreten. Er liegt im Verlauf des Sinus rectus, der durch die Vereini-gung der Falx cerebri mit dem Tentorium cerebelli und der Falx cerebelli gebildet wird. Dieser Ruhepunkt wurde zu Ehren ihres Entdeckers „Sutherland-Fulcrum“ ge-nannt. Ein weiterer Begriff dafür ist „automatic shifting suspended fulcrum“. Die Membranen besitzen sekundäre Anheftungspole an verschiedenen Schädelkno-chen, um ihre Leit- und Regulierungsfunktion für die kraniale Gelenkbewegung erfül-len zu können. Ihre Anheftungen sind V-förmig, umschliessen die grossen Hirnve-nenleiter und können damit die Drainagefunktion im Schädelbereich beeinflussen. Viele Anatomiebücher illustrieren das. Die Falx cerebelli erstreckt sich vom Sinus rectus nach caudal zum Foramen mag-num und vereint sich hier mit der intraspinalen Membran. „So entsteht die extrem wichtige Verbindung zwischen dem membranösen Gelenkmechanismus des Schä-dels und des Kreuzbeins.“ (Magoun, Osteopathie in der Schädelsphäre). Die Verbindung ist wie ein im Wirbelkanal freihängender Schlauch, der das Rückenmark umhüllt. Dieser Schlauch ist nur am Foramen magnum, an C2 und C3 sowie ab S2 nach unten fest verankert. So besteht bei normaler Beweglichkeit der Wirbelsäule nie Verletzungsge-fahr für das Rückenmark. Die intracranialen und intraspinalen Membranen erstrecken sich als Reziproke Span-nungsmembran in die Foramina der Schädelbasis und die Foramina intervertebrales bis in die Scheiden der Nerven und Blutgefässe. Damit ist die Verbindung zum Fas-ziensystem des übrigen Körpers gegeben. Sutherland vergleicht die Funktion der Reziproken Spannungsmembran mit dem Tauziehen von Gruppen an jedem Ende eines Seils. Das Seil stellt die Reziproke Spannungsmembran dar. Das Seil wird hierhin und dorthin gezogen – und es bleibt dabei kontinuierlich unter Spannung. Die Gruppen kommen vielleicht vorübergehend an einen Punkt der Balance, zu einem Punkt der Stille. Dies entspräche dem Fulkrum einer Waage. Folgen abnormer duraler Spannungen (Liem, Kraniosakrale Osteopathie)
• Venöse Abflussstörungen des Schädels, verminderte Drainage des Gehirns • Vaskuläre Versorgungsstörungen der Hirngewebe • Störungen der Fluktuation der Hirn- und Rückenmarksflüssigkeit • Kopfschmerzen, intrakraniale und retroorbitale Schmerzen über die sensible Innerva-
tion der Duralmembran (Hirnnerve V, X und erste drei Zervikalnerven) • Gesichtsschmerzen, abnorme Spannungen der Kaumuskulatur (Hirnnerv V) • Funktionsbeeinträchtigung aller Hirnnerven und Hirnnervenganglien, z.B. an Durch-
trittstellen im Schädel und an den intrakranialen Duralmembranen sowie an den Um-hüllungen der Hirnnerven
• Bewegungseinschränkungen der Schädelknochen und des Sakrums • Funktionsstörungen der Spinalnerven (Druchtrittsstelle durch die Dura mater) • Beeinträchtigung der Hypophyse (Diaphragma selli)

Embryologie aus Sicht der Craniosacralen Osteopathie 12/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
IV 5. Die Motilität der Neuralröhre (Hartmann, Das grosse Sutherland-Kompendium: Unterweisung in der Wissenschaft der Osteopathie, I-30-31; Die Schädel-sphäre III-28) Physiologische Aktivität manifestiert sich als Bewegung der Zellen, die mechanische Merkmale besitzen, also Motilität – eine inhärente aktive Eigenbewegung. Die Motili-tät des Neuralrohrs, das heisst, die Motilität von Gehirn und Rückenmark ist eine wichtige Aktivität des lebendigen Körpers, deren Ursprung sinnvollerweise in der embryonalen Phase mit der Entwicklung des Neuralrohrs gesucht werden kann (Sut-herland). Die Motilität der Nervenzellen, besonders der Gehirnzellen übernimmt im Ausdruck des Primären Atemmechanismus eine mechanische Funktion. Deshalb hat das Neuralrohr neben seiner neurophysiologischen Aktivität, der Übermittlung von Nachrichten, auch eine mechanische Aktivität. Dies ist eine unwillkürliche rhythmische Bewegung, die sich auch auf das Ventrikel-system auswirkt. Seine Form ändert sich rhythmisch und beeinflusst so die zirkulie-rende Aktivität der Zerebrospinalen Flüssigkeit. Diese zirkulierende Aktivität bewirkt eine Bewegung der Membrana arachnoidea und der Dura mater. Als Reziproke Spannungsmembran (siehe oben) bewirkt sie eine Mobilität in den Gelenken der Schädelbasis und des Gesichtsschädels. Auch die Mobilität der direkt aus Kopfme-soderm entstehenden Knochen des Schädeldaches passt sich dieser Motilität inner-halb von Gehirn und Rückenmark und der Fluktuation der zerebrospinalen Flüssig-keit an.
Flexions- und Extensions-phase im Gehirn manifestie-ren sich aus einer langsa-men und rhythmischen Auf- und Entrollung der Gross-hirnhemisphären. In der ei-nen Phase verkürzt sich ihr longitudinaler Durchmesser, während sie sich nach lateral verbreitern, in der anderen Phase verlängern sie sich in ihrem longitudinalen Durch-messer und verengen sich lateral. Abb. 2
Das Ventrikelsystem wird in diese rhythmische Bewegung mit einbezogen. Die late-ralen Ventrikel expandieren in ihrer widderhornartigen Form. Der dritte Ventrikel ex-pandiert V-förmig und der vierte rautenförmig, während das am Foramen magnum fest verankerte Rückenmark nach oben gezogen wird und die Zerbrospinale Flüssig-keit innerhalb des Spatium subarachnoidale und den Ventrikeln fluktuiert. Während der Phase der Exhalation des Rückenmarks entspannen sich die Gyri, die Ventrikel ziehen sich zusammen, das Rückenmark sinkt abwärts und die Zerebrospi-nale Flüssigkeit fluktuiert erneut innerhalb des Spatium subarachnoidale und den Ventrikeln.

Embryologie aus Sicht der Craniosacralen Osteopathie 13/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Sutherland beschreibt den Spatium subarachnoidale unterhalb des Gehirns als Was-serbett, auf dem das Gehirn nicht nur ruht, sondern als seine basiläre Wiege schau-kelt.
IV 6. Die Mobilität der Schädelknochen Sutherland hat die Nähte zwischen den Schädel-knochen in ihrer Ver-schiedenheit aufs Ge-naueste erforscht und für jeden Knochen ihm zu-gehörige Bewegungen entlang seiner Nähte de-finiert. Die unpaaren Knochen (Sakrum, Okci-put, Sphenoid, Os fronta-le und ethmoidale bewe-gen sich in der Flexions-phase um eine transver-sale Achse entlang einer Mittellinie. Paare Kno-chen (Ilea, Ossa parieta-lia, temporalia, Maxillae) drehen um bestimmte Drehachsen von der Mit-
tellinie weg. In der Ex-tensionsphase bewegen sich die Knochen zurück.
IV 7. Die unwillkürliche Mobilität des Sakrums zwischen den Beckenknochen Es handelt sich hier um eine Bewegung, die durch die Reziproke Spannungs-membran vermittelt wird – und nicht um eine haltungsbedingte Mobilität der Becken-schaufeln in Bezug auf das Sakrum. (Abb. 3)
IV 8. Die Funktion des Primären Atemmechanismus Der Bewegungsausschlag ist zwar minimal, nichtsdestotrotz ist er bedeutend für fol-gende Funktionen:
• Homöostase, Gleichgewichtsregulation der physiologischen Körperfunktionen • Optimales Funktionieren der Organe • Wesentliche Rolle bei der inneren Gewebsatmung • Rhythmische Förderung des Stoffwechselaustausches in jeder einzelnen Kör-
perzelle Die Funktionen werden dank den offenen Verbindungskanälen in Schädel- und Wir-belsäulenbereich vom Fasziensystem ins gesamte Körpersystem übertragen.
Abb. 3 Bewegungsrichtung der unpaaren Knochen in der Inspirations-phase

Embryologie aus Sicht der Craniosacralen Osteopathie 14/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
IV 9. Der Rhythmus Puls und Atmung beeinflussen die Druckverhältnisse im Gehirn. Es gibt darüber hin-aus wellenförmige Bewegungen, die weder mit dem Herz- noch mit dem Atemrhyth-mus zusammenhängen, sondern konstant ihren eigenen Zyklus beibehalten. Es gibt unterschiedliche Beobachtungen zum Zyklus des cranio-rhythmic-impuls. Man kann zwischen 6 – 12 Bewegungsphasen pro Minute zählen. Andere Osteopathen haben von einer grossen Gezeitenbewegung gesprochen mit einer Frequenz von 6 – 10 Zyklen. James Jealous beschreibt eine Potency Tide (Mid Tide) mit 2.5 Zyklen pro Minute und die noch langsamere Long Tide mit einem Zyklus von 90 bis 100 Sekun-den. Diese letzte Tide ist von Jealous als Manifestation der Dynamischen Stille be-zeichnet worden. Er legt sie in der ventralen Mittellinie, der Chorda dorsalis an (vgl. Kapitel VII, 1.3.).
IV 10. Erklärungsmodelle für den Primären Atemmechanismus (Liem, Torsten, Kraniosakrale Osteopathie)
Die Motilität entsteht durch die rhythmische Kontraktion der Oligodendroglia-zellen. Schon Leland Clark hatte an Tierexperimenten mit Katzen solche Aktivitäten festgestellt. Andere Forscher haben Neurogliazellen in Gewebekulturen angesetzt und ihre rhythmische Pulsation beobachtet.
Upledger, John E. räumt diesen Kontraktionen nicht genügend Kraft ein, den CRI aufrechterhalten zu können. Er hat das Druckausgleichsmodell geschaffen. Es besagt, dass die Zerebrospinale Flüssigkeit durch den Plexus choroideus im Ventri-kelsystem wesentlich schneller erzeugt wird, als die Liquorresorption durch die A-rachnoidkörper zurück in den venösen Kreislauf stattfinden kann. Erfolgt die Liquo-rerzeugung schneller als die Resorption, so entsteht eine obere Druckgrenze. Sobald der hydrostatische Druck im Schädelinneren die Schädelnähte auf eine bestimmte Weite auseinander drückt, wird ein Streckreflex in der Knochennaht aktiviert, der dem Ventrikelsystem des Gehirns befiehlt, die Liquorerzeugung einzustellen. Der hydro-statische Druck sinkt, weil bei unterbrochener Produktion die Resorption weiterhin stattfindet. Sobald ein unterer Schwellenwert erreicht wird, schaltet sich die Liquorer-zeugung wieder ein. Diese rhythmische Druckerhöhung und –senkung verursacht ih-rerseits die rhythmische Veränderungen der Grenzen des teilweise geschlossenen hydraulischen Systems. B. Cabarel und M. Roques untersuchten die Beziehung des PRM auf die Fas-zien. Sie stellten eine Hypothese auf über den Einfluss des PRM und der Flexion- und Exspirationsphase auf die Bindegewebe und Faszien des Körpers. Danach be-steht ein Zusammenhang zwischen dem Elektrolytspiegel in der Grundsubstanz und der Exspirationsphase, wo der Liquor cerebrospinalis und die extrazelluläre Flüssig-keit aus den extrazellulären Räumen verdrängt wird. Besonders untersucht haben sie in diesem Zusammenhang Hyaluronsäure und das Enzym Hyaluronidase in der Grundsubstanz des Bindegewebes und der Gelenkflüssigkeit. Sie beeinflussen Ge-websviskosität und Permeabilität. Wenn man auf der Entwicklungswachstumsebene nach dem Ursprung der Ei-genbewegung sucht, ist es nach Liem das Stirnbein (er meint vielleicht die Dura oder darin wachsende Ossifikationszentren, Anm. der Verfasserin), das das Wachstum der Grosshirnhemisphären nach anterior begrenzt, so dass sie sich widderhornartig nach hinten einrollen und sich in ihrer Entstehung nach superior (Lobus frontalis),

Embryologie aus Sicht der Craniosacralen Osteopathie 15/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
posterior (Lobus parietalis), inferior (Lobus occipitalis) und nach anterolateral (Lobus temporalis) bewegen. Nach Blechschmidt ist es die Dura, die die neuralen Strukturen im Längen-wachstum hemmend beeinflusst und sie dazu bringt, sich einzurollen. Ausserdem führt die langsamer wachsende Anlage von Herz und Arterien dazu, dass das Neu-ralrohr sich mehr und mehr einrollt. Hier setze ich mit meinen Überlegungen zum Ur-sprung des Primären Atemmechanismus an.

Embryologie aus Sicht der Craniosacralen Osteopathie 16/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
V Embryologie
V 1. Die Bedeutung der Embryologie für die Praxis Die Embryologie ist für uns Praktiker die Grundlage für das Verstehen vieler struktu-reller, physiologischer, funktioneller und dysfunktioneller Zusammenhänge, die wir palpatorisch und therapeutisch erfassen. Mit dem Verständnis von Eigendynamik des Gewebewachstums und der Entwick-lungsdynamik der Gewebe in Wechselwirkung zueinander können strukturelle Dys-funktionen zum Zeitpunkt der Untersuchung in Beziehung zueinander gebracht wer-den. Strukturelle Funktionsstörungen können aber auch unter dem Gesichtspunkt des Zeitfaktors verstanden, wahrgenommen und behandelt werden - also der Zeit, die seit der Dynamik von prä- und postnatal entstehenden Relationen und den jetzi-gen Verhältnissen der Strukturen vergangen ist. Die Kenntnis der embryologischen Wachstumsbewegungen gibt dem Praktizierenden eine Orientierung darüber, wie sich inhärente, eigenständige Bewegungen der ein-zelnen Strukturen im optimalen Zustand anfühlen. Neben biomechanischen Bewe-gungsachsen in Gelenken und anderen Strukturen existieren auch embryologische dynamische Bewegungsachsen. Sie beruhen auf dem jeweiligen Wachstums- bzw. Gestaltungsprozess der Strukturen und bestehen auch nach Abschluss des Wachs-tums als inhärente Bewegungen in den Geweben weiter. So wie in der Psychotherapie traumatische Erlebnisse besonders der frühen Lebens-jahre die weitere Persönlichkeitsentwicklung beeinflussen können und durch Be-wusstwerdungsprozesse die Einschränkungen aufgelöst bzw. integriert werden kön-nen, so ist eine Herangehensweise auch bei der Arbeit mit dem Gewebe grundsätz-lich möglich. Indem der Praktizierende seine Aufmerksamkeit auf die inhärenten Spannungen und Gewebebewegungen richtet, ist er in der Lage, durch nach- oder vorgeburtliche Einflüsse oder Traumata entstandene Gewebespannungen zu palpie-ren und sich auflösen zu lassen. Wenn in der Pränatalpsychologie von pränataler Existenz oder pränataler Erfahrung gesprochen wird, ist meist die fetale Existenz gemeint, also erst die Phase mit prinzi-piell vollständig ausgebildetem Körperplan und angelegten Organsystemen. In der Pränatalpsychologie geht man davon aus, dass Transformationen oder Variationen der frühesten pränatalen Erfahrungen in späteren Lebensphasen erneut erlebbar sind, sogar solche, die stattfanden, bevor unser Nervensystem sich entwickelt hat. Dabei stellt sich die Streitfrage, ab wann ein Embryo in der Lage ist, „zu erleben“ und motiviert zu handeln, wenn man konventionellerweise davon ausgeht, dass Seelen-leben und Verhalten nur bei einem funktionsfähigen Nervensystem möglich sind. Biologisch gesehen umfasst das pränatale Leben aber auch die Phase des embryo-nalen Lebens der ersten 8 – 10 Wochen, das aus Organogenese und Somatogenese besteht. Ich gehe in dieser Arbeit davon aus, dass der Mensch von Anfang an „be-wusst“ seine Entwicklung erlebt und gestaltet. Jaap van der Wal und Erich Blechschmidt vertreten beide den Standpunkt, dass diese Entwicklung in der embry-onalen Phase ausschliesslich eine morphologische ist. Diese Arbeit zielt nicht darauf ab, Gewebeabnormitäten mit Aethiologie in der embry-onalen Phase (Teratologie) aufzufinden oder deren Behandlung zu erklären. Es geht vielmehr darum, Grundlagen dafür zu sammeln, wie der Primäre Atemmechanismus als physiologischer Vorgang seine Funktion aufnimmt.

Embryologie aus Sicht der Craniosacralen Osteopathie 17/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
V 2. Geschichtlicher Überblick über die Untersuchung der Humanembryologie Drews, Ulrich, Taschenatlas der Embryologie, Georg Thieme Verlag, 1993 Die Schwierigkeit für die Untersuchung der embryologischen Entwicklung des Men-schen stellte das Anschauungsmaterial dar. Erstens war es schwierig zu Einzelex-emplaren menschlicher Embryonen zu kommen, zweitens war es noch schwieriger, sie altersmässig zu bestimmen, weil das Datum der Konzeption meist nicht bekannt war. 1908 wurde eine erste „Normentafel“ der menschlichen Entwicklung von Keibel et al. herausgegeben. Es war der Versuch, die damals bekannten Einzelexemplare von menschlichen Embryonen in einer Entwicklungsreihe anzuordnen. 1914 wurde das „Department of Embryology of the Carnegie Institution of Wa-shington“, benannt nach dem Industriellen und Philanthropen Carnegie, begründet. Begründer war Franklin P. Mall, ein Schüler von Keibel 1942 G. L. Streeter ordnet die Embryonen der Carnegie-Sammlung in beschriebene Entwicklungsstadien auf Grund von Gestaltmerkmalen ein. Jedem Stadium wird entsprechend dem Auftreten von Gestaltmerkmalen ein zeitlicher Bereich zugeord-net. („Developmental Horizons I – XXIII“) 1987 setzte Ronan O’Rahilly diese Arbeit fort. 1999 wurde die Zuordnung mit den neuen Möglichkeiten der Ultraschalluntersuchung revidiert. 1942 – 1956 wurden auch die frühen Implantationsstadien des Menschen beschrie-ben. A. T. Hertig, Pathologe und J. Rock, Gynäkologe, begannen in dieser Zeit sys-tematisch die Schleimhaut von 210 aus chirurgischer Indikation entfernten Gebärmüt-tern sorgfältig nach Implantationsstellen abzusuchen. So konnten sie die Carnegie-Sammlung um 34 Embryonen erweitern. Erich Blechschmidt klassifizierte seine Sammlung menschlicher Embryonen nach den Carnegie-Stadien und fügte seine Exemplare der Carnegie-Sammlung ein. Abb. 4 zeigt eine Übersicht der Frühentwicklung in der 1. bis zur 3. Woche. Abb. 5 stellt die eigentliche Embryonalperiode bis zur 8. Woche mit den beschriebenen Ent-wicklungsstadien nach Gestaltmerkmalen dar. Für meine Diplomarbeit sind folgende Embryonalperioden von besonderer Bedeu-tung: Ab ca. 15. Tag die Bildung von Primitivstreifen, Primitivgrube, Chordafortsatz, Axialkanal und die Induktion der drei primitiven Achsenorgane Chorda dorsalis, So-miten und Neuralrohr (Neuralplatte entsteht ca. am 23. Tag, die eigentliche Neural-rohrbildung bis ungefähr am 28. Tag). Mit der Invagination der Ektodermzellen in den Primitivstreifen entsteht das Mesoderm als dritte Keimschicht. Damit wird eine ele-mentare Funktionsgliederung des späteren Organismus gespiegelt. Schon hier sei darauf hingewiesen, dass Erich Blechschmidt für seine raum-zeitlich beschreibende Gestaltungsanatomie wenig von den drei bekannten Keimschichten spricht, weil er die Wachstumsbewegungen von Zellverbänden direkt in Lagebeziehungen zueinan-der untersucht. Besonders eingehen werde ich auf die laterale und die craniocaudale Abfaltung zur „Körpergrundgestalt“ (nach Seidel) - der damit einhergehenden Anlage aller elemen-taren Funktionsbereiche des Embryos.
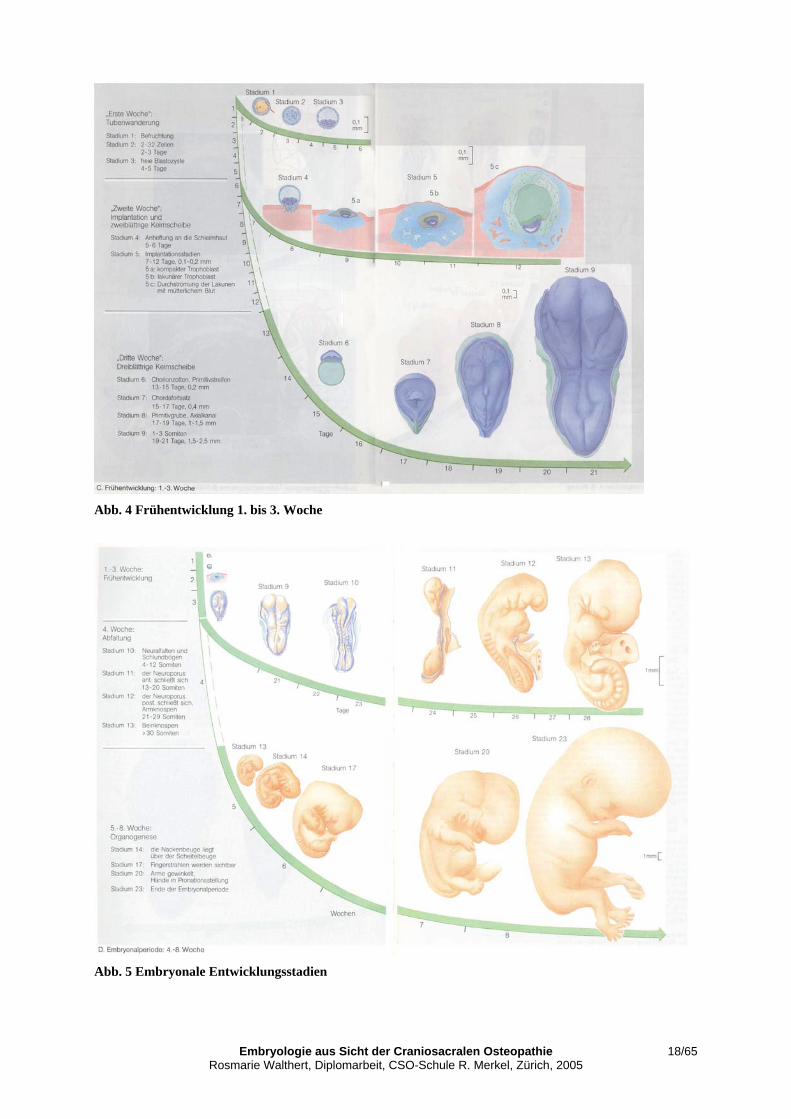
Embryologie aus Sicht der Craniosacralen Osteopathie 18/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Abb. 4 Frühentwicklung 1. bis 3. Woche
Abb. 5 Embryonale Entwicklungsstadien

Embryologie aus Sicht der Craniosacralen Osteopathie 19/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
V 3. Der Mensch als Embryo zwischen Himmel und Erde Herr Dr. med. Jaap van der Wal habilitierte an der Universität Maastricht im Bereich der menschlichen Propriocepsis (Bewegungssinn). In diesem Zusammenhang stellte sich ihm die Frage: was bewegt uns? Sein Hauptinteresse gilt den Bereichen der menschlichen Embryologie und angrenzenden Disziplinen wie Genetik, Entwick-lungsbiologie und Philosophie der Wissenschaft. Wichtige Quellen der Inspiration sind für ihn die Anthroposophie von Rudolf Steiner sowie phänomenologische Philo-sophie und die wissenschaftlich phänomenologischen Arbeiten von Goethe. Seine Ausführungen kommen von der gängigen Embryologie her. Aber er versucht, den Blickwinkel zu erweitern zu einer „Embryosophie“. Er führt zu einem mit- und nachgefühlten Verstehen der Embryonalentwicklung als fortwährend sich änderndem und metamorphosierendem Zusammenhang von Formen und Formprozessen. Mit Hilfe der goetheanischen phänomenologischen Anschauungsmethode kann die „Gestik“ dieser Prozesse als menschliches Verhalten aufgefasst werden. Einige Aussagen von Jaap van der Wal, die umfassendes Wahrnehmen der Entwick-lungsprozesse fördern:
Primär ist die Einheit, sekundär sind die Teile. Das Wissen um die Teile ist re-duzierte Kenntnis. Die partizipierende Kenntnis umfasst auch die reduzierte Kenntnis.
Das wissenschaftliche Menschenbild zieht Grenzen, weil es keine Methoden kennt, über Jenseitiges etwas auszusagen. Es sind einzig konzeptionsmässige Grenzen, weil die Methoden fehlen. Die Konsequenz daraus ist leider, dass mangels einer Erfassungsmethode nur richtig sein kann, was innerhalb der Konzeptgrenzen möglich ist.
„Biologische Systeme benehmen sich“ (Zitat des Biologen Weiss). Die Defini-tion von „sich Benehmen“ als Bezeichnung für Handeln und Tun wird weitergeführt zu „sich Ausdrücken in ununterbrochen sich veränderndem morphologischem Aus-sehen“. Das heisst, dass ich die sich ständig verändernde Form in der ganzen Zeit und in ihrem Ganzen lesen soll. Eine Analogie: Die Rose in der Vase ist nicht wirklich die Rose. Ich muss den Faktor Zeit in meinem Bild der Rose mit einbeziehen: vom Samen, zur Pflanze, zur Knospe, zur Blume und weiter zum verwelkenden Stadium. Erst die Formen all dieser Stadien in ihrer Zeit sind die Rose.
Die Lebenswelt soll nicht nur als Beobachter, also von ihr abgespaltet, be-trachtet werden. Man soll ihr nicht gleichermassen nur von aussen zuschauen. Er fordert auf, an der Realität teilzunehmen, sich herzensmässig mit ihr zu verbinden. „Nur der partizipierende Körper hat Geist.“ „Ich erfahre, also bin ich“. Oder: „Ich fühle, also bin ich“ Aus: Damasio, A. R., Descartes’ Irrtum. Die Lebenswelt kann in Verbindung erfahren werden (primäre Lebenswelt) oder in Abgrenzung beobachtet werden (sekundäre Lebenswelt). Beides gehört wertfrei zusammen. Die primäre Lebenswelt kennt die sekundäre, aber die sekundäre Lebenswelt kennt die primäre nicht. Die sekundäre Lebenswelt ist notwendig aber nicht genügend. Für die Embryonalphase gilt dem-nach das Beispiel: DNA und Gene sind notwendig aber nicht genügend.
Es ist ein Unterschied, ob ich sage, ich habe einen Körper (und dazu einzelne Muskeln zu einer Faust spanne) oder ob ich sage: ich bin ein Körper (mit unter-schiedlichen Fäusten wie jener aus Wut, Sieg/Triumph oder aus Angst, Machtlosig-keit etc.)
Embryonale Existenz ist eine Art stille, stumme und introvertierte Existenz. Als erwachsene Menschen drücken wir uns mit unserem Körper aus: Die Welt ist unser Ziel und der Körper dient dazu, dieses Ziel zu erreichen (zentrifugale Orientierung).

Embryologie aus Sicht der Craniosacralen Osteopathie 20/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Der Embryo jedoch drückt sich in seiner körperlichen Organisation aus (zentripetale Organisation). Die Vorstellung, ein Embryo „tue noch nichts“ oder „handle noch nicht“ ist eine krasse Fehleinschätzung und Abwertung. Die Handlung, die Tätigkeit ist auf ihn selbst gerichtet, nach innen. Sie stellt menschliche Handlung und menschliches Verhalten dar.
Das Seelische übt sich in der Embryonalphase voraus. In dieser Phase funkti-oniert das Seelische morphologisch, dann physiologisch (Wachstumsphase) und später psychologisch/geistig (Erwachsenenalter).
Nehmen wir ein Anschauungsbeispiel aus der Phase vom 13. bis 15. Tag. Der Keimling (Blastocyst) hat sich in der 2. Woche in der Form des zweischichtigen „Embryoblasten in sei-nem Mantel, dem Trophoblasten“ in die Schleimhaut der Gebärmutter eingewuchert und niedergelassen. Der Trophoblast ist ein Mantel an der Peripherie. Der Embryoblast ist im Zentrum. Die eine Schicht, das Ek-toderm ist dem Dottersack zugewandt. Die andere Schicht, das Entoderm ist vom Amnion, dem Darmblä-schen umgeben. Der Embryoblast ist der Kern. Aus ihm wird sich der eigentliche Embryo entwickeln. Der Mantel ist ebenfalls ein Teil des Embryos, der aber zur
Plazenta, zu Hüllen wird. Trophoblast und Embryoblast sind zuerst miteinander ver-bunden. Abb. 6: Der dünner werdende Haftstil (H) verlagert sich von hinter dem Am-nion zunehmend zum caudalen Ende der Keimplatte hin. Dort entstehen im wach-senden Mesoderm (gelb) erste Blutinseln mit Blut und Gefässen. Erst in der 3. Wo-che wird der Trophoblast definitiv zum Aussenei und kleidet sich an seiner Innenseite mit Mesoderm (gelb) aus. Wir organisieren uns aus Einheit in Vielheit, in Organe. Ohne spezielle Organe dafür zu besitzen, erfüllt der Trophoblast alle Funktionen, wie sie für einen lebendigen Organismus charakteristisch sind:
• Gasaustausch wie in der Lunge • Exkretion von Stoffwechselendprodukten und Elektrolyten wie in der Niere • Resorption von Nahrungsstoffen wie im Magen-Darm-Kanal • für Wachstum und Entwicklung notwendige Stoffwechselvorgänge wie in Le-
ber • Endokrine Steuerungsprozesse wie in den späteren sekretorisch tätigen Drü-
senorganen • Blutbildung • Wärmeregulation
Der Embryo ist gleichsam eingestülpt in seine Hülle, die für ihn Lebens und Steue-rungsprozesse übernimmt, wie er sie später mehr und mehr selber wird übernehmen müssen je mehr Organe er dafür entwickelt. Nun löst sich der Embryo von seiner ersten Umhüllung, dem Trophoblasten (Abb.6) und hüllt sich mit der Abfaltung in das Amnion ein, das zur Fruchtblase wird (vg. Abb. 36). Dabei entsteht der Nabel. Von nun an werden die überlebenswichtigen Funktio-nen über den Nabel sichergestellt - kanalisiert. Der alte Raum (Trophoblast) wird Mutterkuchen (Plazenta). Eigentlich sollte er Kinderkuchen heissen, denn er ist aus dem ersten Hüllmaterial der befruchteten Eizelle und nicht etwa aus mütterlichem Gewebe entstanden. Das Amnion bildet die neue Höhle, die Fruchtblase.
Abb. 6 Hüllen

Embryologie aus Sicht der Craniosacralen Osteopathie 21/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Dieser Vorgang kann als ein erster Ablösungsprozess oder Trennungsprozess ver-standen werden, wie sie dem sich entwickelnden Menschen sein ganzes Leben lang in seinem Individuationsprozess immer wieder geschehen wird. Jaap van der Wal spricht daher vom Menschen als Embryo auch im Erwachsenenal-ter. So wie der Embryo Hüllen abwirft in seiner frühesten Ent-Wicklung, so entwickeln sich der junge und später der erwachsene Mensch weiter. Im Laufe dieses Prozes-ses legt er auf verschiedenen Ebenen immer wieder Hüllen ab. Der Mensch bleibt sein Leben lang in Bewegung und Entwicklung.
V 4. Der Embryo – eine Dokumentation der kinetischen Anatomie Herr Prof. Dr. med. Erich Blechschmidt (1904 – 1992) entwickelt Anfang der 60er Jahre die Kinetische Anatomie – eine in Zeit und Raum vergleichende Untersuchung gleicher Strukturen und Formen von Embryonen in Bewegung – d. h. anhand von Embryonen kleinstunterschiedlichen Alters. Während mehrerer Jahrzehnte ging er in aufwändigen Forschungsarbeiten folgenden Fragen nach: Wie geschieht es, dass bei gleichem genetischem Material in den ein-zelnen Zellen sich diese verschieden entwickeln, d.h. differenzieren? Welche Vor-gänge führen dazu, dass die ursprüngliche Pluripotenz der Zellen eines wenige Tage alten Blastocysten mehr und mehr abnimmt? Was geschieht eigentlich, dass schlussendlich aus einer Zelle eine Knorpelzelle aus einer anderen eine Knochenzel-le oder Muskelzelle wird und welche Vorgänge sind an der Kapselbildung oder an der Faszienbildung von Organen beteiligt?
V 5. Die Forschung In 30 Jahren Forschung ist Erich Belchschmidt durch seine Untersuchungen zum Schluss gekommen, dass auf dem Weg von aussen durch die Zellgrenzen den Chromosomen vorgeschrieben wird, welche Gene sie jeweils in den aufeinander fol-genden Entwicklungsschritten verwenden sollen. Er geht also nicht davon aus, dass im genetischen Material gleichsam statisch schon ein Muster für jeden Entwicklungs-schritt liegt. Er widerspricht der Annahme, dass ein Zellkern gleichsam schon einen massstäblich gezeichneten Plan im Sinne einer Vorformung enthält. Technisches Vorgehen Aufbauend auf der Lehre der Körpergestalt (Morphologie) nimmt er in aufwändiger Weise unzählige räumliche und zeitliche Messungen an menschlichen Embryonen kleinstunterschiedlichen Alters und Grösse vor, um Gestaltungsvorgänge als Be-wegungsabläufe aufzuzeigen. Er hält Differenzierungen, also Formveränderungen, als Veränderungen des äusseren Erscheinungsbildes fest. Dazu wurden Serien-schnitte von fast lebensfrisch fixierten und speziell eingefärbten Embryonen einzeln fotografisch und anschliessend zeichnerisch vergrössert. Nach ihnen wurden für je-den einzelnen dieser mikroskopischen Schnitte Wachsmatrizen geschnitten, die, mit Kunststoffen ausgelegt, nach Abschmelzen des Wachses zu einem Totalmodell zu-sammengesetzt wurden. Im anatomischen Institut an der Universität in Göttingen steht die Sammlung von rund 70 Schnittserienrekonstruktionen, Totalmodelle von ca. 80 cm Höhe. Erst die Totalrekonstruktionen erlaubten einen sogenannten regionalen Vergleich, d.h. einen

Embryologie aus Sicht der Craniosacralen Osteopathie 22/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
anatomischen Vergleich der Strukturen in den verschiedenen Körperregionen unter-einander und erst damit die Ermittlung von Entwicklungsbewegungen. Mit seinen Untersuchungen löst Erich Blechschmidt die räumlich beschreibende Zu-standsanatomie mit einer raum-zeitlich beschreibenden Gestaltungsanatomie ab. Die Frage nach Art und Weise der Differenzierung des menschlichen Körpers sucht er mit biokinetischen Eigenschaften zu beantworten und nicht mit Chemie oder Ge-schichte. Dies ist für Therapeuten, die Bewegung und Funktion untersuchen und be-handeln ein erfreulich vertrauter Denkansatz in der lebendigen Struktur seitens eines Wissenschafters.
V 6. Die Kinetische Entwicklungstheorie Mit der Kinetischen Entwicklungstheorie stellt er ein neues Grundgesetz auf. Es be-sagt, dass die Eigenart eines Organismus bereits zu Beginn der Individualentwick-lung mit der Empfängnis vorausgesetzt sein muss und davon ausgegangen werden kann, dass die Erhaltung der Individualität auf der Kontinuität eines spezifischen Stoffwechsels beruht. Der Organismus versucht fortgesetzt während der ganzen Dauer seiner Entwicklung „zu bleiben, was er schon im Augenblick der Geburt war“. Die Mittel, mit denen er die Entwicklungsreize kompensiert, sind u.a. die Entwick-lungskinetik und Entwicklungsdynamik. Folgende Aussagen zeigen die verschiedenen Aspekte und die daraus ableitbaren theoretischen und praktischen Konsequenzen für Morphologie, Physiologie, Bioche-mie und insbesondere für die Genetik und Embryologie auf (Aus: Die Frühentwicklung des Men-
schen, Verlag Hogrefe, 1966): Jedes Organ hat als Bestandteil des Organismus sowohl eine entwicklungski-
netisch bedingte Lage, als auch eine entwicklungskinetisch zugehörige Form und Struktur. Lage, Form und Struktur sind zusammengehörige Eigenschaften. Ohne die-se Eigenschaften gäbe es keine Entwicklung, keine Ernährung, kein Wachstum und keine Fortpflanzung. Keine Verhaltensweisen und keine bewusste Tätigkeit vermöch-ten sich zu entwickeln.
Organe von Mensch und Embryo lassen sich systematisch als Momentanauf-nahme von den drei prägenden Eigenschaften Lage, Form und Struktur als Entwick-lungsbewegungen des Organismus untersuchen.
Jedes Organ hat dank seiner Entwicklungskinetik sowohl eine Lageentwick-lung (Topogenese), als auch eine von ihr abhängige Formentwicklung (Morphogene-se) und eine ihr zugehörige Strukturentwicklung (Tektogenese). Im Hinblick auf die Bedeutung der Entwicklungsbewegungen ergibt sich aufgrund der drei voneinander abhängigen Topogenese, Morphogenese und Tektogenese eine funktionelle Bedeutung:
Die Gestaltungsfunktionen sind die Grundfunktionen der Organe. Erich Blechschmidt unterstreicht die Individualentwicklung oder Ontogenese des Menschen und setzt sich von Darwin ab.
V 6.1. Materialbewegungen und Stoffwechselfelder Die Lageentwicklung der Zellverbände ist eine wichtige Voraussetzung für ihre struk-turelle Differenzierung. Der Zellstoffwechsel gestaltet sich unterschiedlich je nach seiner Lage und Umgebung. Die Aufnahme der Nahrung bzw. Abgabe von Stoffen ist an der Oberfläche eines Zellverbandes anders als in seinem Inneren. Entsprechend

Embryologie aus Sicht der Craniosacralen Osteopathie 23/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
unterschiedlich entwickeln sich die Zellen. Die Entwicklung des menschlichen Keims in der ersten Woche zum Beispiel kann diesen Gedankengang anschaulich verdeut-lichen. Während seiner Reise im Eileiter zur Einnistungsstelle in der Gebärmutterwand nimmt das menschliche Ei Nahrung von der äusseren Umgebung auf. Zu Beginn lie-gen alle seine Zellen (Blastomere) noch an der Oberfläche und haben so Kontakt zum äusseren Raum, dem Tubenraum. Mit fortschreitender Furchung (Unterteilung des anfangs einzelligen Keims in Teilkörper) liegen immer mehr Zellen in seinem In-neren, in der Tiefe. Die neu gebildeten Zellen halten sich durch gegenseitige Stoff-austausch aneinander (Abb. 7). Die Abbauprodukte, die beim Verbrauch der Nahrung entstehen, geben die Zellen zunächst an die Umgebung zurück oder stauen sie im Inneren des Eis (Abb. 7). Der Blastocyst bleibt bis zur Einnistung gleich gross. Die Grösse seiner Zellen hingegen nimmt ab, weil die Zellen Flüssigkeit abgeben (Abb. 7). Entsprechend stauen sich im Interzellulärraum mehr und mehr Abscheidungsprodukte. Weil diese grösstenteils
wasserlöslich sind, vermag die Interzellulärsubstanz durch Osmose von aussen Wasser aufzunehmen. So entsteht im Innern des Blastocysten ein Flüssigkeitstropfen, der bei gleich bleibender Grösse des Blastocysten einen Wanddruck auf die umschliessenden Blastomere ausübt. Die Furchung des Blastocysten erfolgt nicht überall gleich schnell. Deshalb kommt der Flüssigkeitstropfen nicht in der Mitte, sondern irgendwo am Rand zu liegen. Der Tropfen ist die erste Ansammlung von Entocoelwasser, dem Vorläufer des späteren Dottersackinhaltes. Als seine Umgrenzung zeigt die Wand des Blastocysten am 4. Tag
an ihrer schwächsten Stelle, wo der Druck der Interzellularsubstanz eine besonders intensive Dehnung hervorbringt, regelmässig eine einzige Schicht abgeplatteter Zel-len. Die winzigen Blastomere des gegenüberliegenden dicken Wandabschnitts fla-chen sich dagegen nicht ab, sondern behalten ungefähr gleiche Dimensionierung in alle Raumrichtungen. Die Zellen des dicken Wandabschnittes (Eidiskus) dürfen e-benso wie die dünnen Zellen des antidiskalen Pols des Eis als ortsgemässe, lokale Zellmodifikationen aufgefasst werden. Im Eidiskus sind die Zellen keiner stärkeren Deformation durch osmotischen Druck ausgesetzt. Sie bewahren die Fähigkeit zu ur-sprünglicher Eigenart. Hier lassen sich vermehrt Mitosen (Zellteilungen) nachweisen. Der Vergleich mit späteren Entwicklungsstadien ergab, dass Mitosen sich regelmäs-sig in Gebieten relativ geringer Zelldeformation finden.
V 6.2. Grenzgewebe und Binnengewebe Die Zellen eines Gewebes sind stets kinetisch durch Materialbewegungen miteinan-der in Verbindung. Die Zellen nehmen Nahrung aus den Zwischenzellspalten und aus ihren Nachbarzellen auf und ziehen sich durch diese Materialaufnahme gegen-seitig an. Andererseits stossen sie sich durch Abgabe von verbrauchten Stoffen ge-genseitig ab (Abb. 7). Das Wechselspiel zwischen Aufnahme und Abgabe ist die Voraussetzung dafür, dass die Zelle sich in bestimmter Weise anordnen und in Form halten.
Abb. 7 submikroskopische Stoffwechselbewegungen

Embryologie aus Sicht der Craniosacralen Osteopathie 24/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Zwei charakteristisch verschiedene Gewebe, die man schon sehr früh findet, sind das Grenzgewebe und das Binnengewebe.
• Das Grenzgewebe grenzt einerseits an Flüssigkeit und andererseits an Bin-nengewebe.
• Das Binnengewebe ist allseits von Grenzgewebe umschlossen. Entsprechend ihrer Lage haben die beiden Gewebearten unterschiedliche Bedeu-tung für das Wachstum. Die Grenzgewebe (Epithelien) bilden Zellmosaike entlang der Flüssigkeiten. Sie leis-ten erste Gestaltungsarbeit. Grenzgewebe entnimmt dem unterliegenden Binnenge-webe Nahrung und gibt Abbauprodukte in die freie Flüssigkeit ab. Abb. 5 zeigt ein di-ckes Epithel mit 2 verschiedenen Grenzschichten. Die innere Schicht wird im Flä-chenwachstum behindert (Pfeile mit Querstrich). Sie verdickt sich durch Nahrungs-aufnahme aus dem Stroma (Pfeilköpfe) Die äussere, schlechter ernährte Schicht wächst langsamer. Mit dem Verbrauch der Nahrungsstoffe ruft das wachsende Grenzgewebe Stoffwechselbewegungen in Richtung von seiner Basis zur freien O-berfläche und gleichzeitig Materialbewegungen entlang seiner Basis im angrenzen-den Binnengewebe hervor. Dies führt zu einem Fluss von Nahrungsstoffen in den In-terzellularspalten des Binnengewebes und dort zu feiner Kanalisierung (Vorläufer der späteren Blutgefässe). Das Konzentrationsgefälle von Nahrungsstoffen ist ein wichtiger dynamischer Faktor bei der Entstehung des Blutstroms. Merkmale von: Grenzgewebe Binnengewebe Abgabe der Abbauprodukte an umge-bende Flüssigkeit
Abbauprodukte werden zu Zwischenzell-substanz
Geschlossene Zellschicht, viele Zellen mit schmalen Interzellularspalten, kein Stau von Interzellularsubstanz
Binnengewebe ist von Grenzgewebe um-schlossen. Staut Abbauprodukte als Zwi-schenzellsubstanz in Zellzwischen-räumen.
Intensives Flächenwachstum entlang der freien Oberfläche
Volumenwachstum: weniger Zellen und damit weniger Wachstumsfermente
Binnengewebe wächst also langsamer als Grenzgewebe. Das Binnengewebe setzt dem mit ihm verbundenen Grenzgewebe Wachstumswiderstand entgegen.
V 6.2.1. Grenzgewebe Die entwicklungskinetischen Untersuchungen von E. Blechschmidt haben von alten deskriptiv-anatomischen Erkenntnissen rund ums Epithel fortgeführt zu einem kine-tisch-anatomischen Verständnis des Grenzgewebes. Seine Untersuchungen zeigten regelmässig, dass die sogenannten Epithelien in Wirklichkeit Diathelien, also Durch-lassschichten sind. Zur Zeit ihrer Entstehung sind sie nämlich stets Zwischenschich-ten zwischen freier Interzellularsubstanz (Flüssigkeitsräumen) einerseits und Binnen-gewebe mit gestauter Interzellularsubstanz andererseits (Abb. 9). An diesen Grenzen weisen die Diathelien zur Zeit ihrer Entstehung und frühen Entwicklung im Organis-mus regelmässig entlang der Kontaktfläche mit dem angrenzenden Stroma (Gewebe, Binnengewebe) ein charakteristisches Flächenwachstum auf, wie es beim Binnen-gewebe, das genetisch die gleichen Erbanlagen wie das Diathel besitzt, nicht zu be-obachten ist. Die von Blechschmidt regional vergleichenden Untersuchungen zeigen,

Embryologie aus Sicht der Craniosacralen Osteopathie 25/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
dass die Abscheidung von freier Interzellularsubstanz charakteristisch und wahr-scheinlich eine Vorbedingung für das Flächenwachstum ist. Wir unterscheiden dickes und dünnes Grenzgewebe. Regelmässig finden wir Grenz-gewebe dort dick, wo es im Flächenwachstum behindert ist, dagegen dort dünn, wo es im Flächenwachstum gefördert wird. Wo immer beim Erwachsenen ein Hautbezirk normalerweise relativ dick geworden ist, war in der Embryonalzeit das Grenzgewebe im Flächenwachstum behindert. Nachfolgende Beispiele zeugen davon. Die Oberhaut des wachsenden Handtellers ist im Wachstum behindert und schon beim Embryo dicker als am relativ grösseren Handrücken. Die charakteristische schwielige Verdickung des Handtellers für die Greiffunktion ist also schon in der Embryonalzeit sichtbar.
Ein typisches Beispiel für dicke und dünne Grenzge-webe ist die Epidermis (Oberhaut) im Bereich des embryonalen Kopfes. Dort finden wir das Epidermis genannte Epithel über dem schnell wachsenden Ge-hirn dünn (Pfeil in Abb. 8, dagegen im Bereich der Schlundfalten sehr dick (punktiert in Abb. 8). Während der weiteren Entwicklung nimmt mit der Verdickung des Epithels auch die Dicke des
Abb. 8
unterliegenden Bindegewebes zu. Mit der Verdickung wird der Nahrungsverbrauch gesteigert. Das Blutgefässsystem wird dichter. Die beim Erwachsenen auffallend starke Durchblutung des Gesichts wird schon embryonal eingeleitet.
Wo das intensive Flächenwachs-tum des Grenzgewebes in seiner Ausbreitung behindert wird, bilden die Zellen warzenartige Zapfen, die in das unterliegende Binnen-gewebe einwachsen
Abb. 9
Im Gegensatz zu dicken Epithelien wachsen dünne Epithelien oft nur einschichtig Sie werden vergleichsweise häufig durch unterliegende zellreiche schnell wachsende Organe wie z.B. Gehirn, Leber oder Herz in kurzer Zeit gedehnt und im Flächen-wachstum wenig behindert. Sie sind typische Diathelien, d. h. Schichten, die befähigt sind, Substanzen senkrecht zu ihrer freien Oberfläche hindurchwandern zu lassen.

Embryologie aus Sicht der Craniosacralen Osteopathie 26/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
V 6.2.2. Keilepithelien
Die Zellgrenzmembranen von Epithelien verbinden in der Regel die innere und äussere Oberfläche auf dem kürzesten Weg miteinander. Sie stehen also senk-recht zur Basis des Zellverbandes. Da der menschliche Körper fast nirgends plane Oberflächen aufweist, haben die Epithel-zellen daher fast immer Keilform. Grenz-gewebe sind also Keilepithelien (Abb. 10). Divergente Keilepithelien haben Zellgren-zen, die zur freien Oberfläche hin divergie-ren. Es sind nach aussen konvex gewölb-te Epithelien.
Abb. 10 Keilepithelien
Konvergente Keilepithelien haben Zellgrenzen, die zur freien Oberfläche hin konver-gieren. Die freie Oberfläche dieses Epithels ist kleiner als die Oberfläche zum Bin-nengewebe hin. Je nach Lage und Form der Keilepithelien sind die wachstumsfunktionellen Bedeu-tungen ihres Flächenwachstums verschieden. Nachfolgende Beispiele zeigen, wie spezifische Keilepithelien junge Organanlagen und/oder die Form der späteren Or-ganentwicklung hervorbringen. Ein Beispiel für konvergentes Keilepithel ist das junge Darmepithel (Abb. 11).
Die wachsenden Zellen üben einen gegenseitigen Wachstumsdruck aufeinander aus. Mit Hilfe des Wachstumsdruckes wird die Oberfläche des Epithels trotz Widerstand des Stromas grösser.
Das Darmlumen erweitert sich. Ein charakteristisch zur freien Oberfläche di-vergentes Keilepithel findet sich an den Enden der jungen Extremitätenanlagen (Abb. 12).
Abb. 11
Abb. 12

Embryologie aus Sicht der Craniosacralen Osteopathie 27/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Auch hier wächst mit dem Wachstum der Zellen der Wachstumsdruck. Morpholo-gisch erkennt man ihn an der lokalen Wachstumsbeschleunigung. Wieder kommt es zur Oberflächenvergrösserung – und zwar in Richtung des geringsten Widerstandes, wie es alle Wachstumsprozesse unter den jeweils verschiedenen Umständen tun. Die junge Extremitätenanlage wächst nicht mit allen ihren Zellen gleich schnell, son-dern an ihrer Wurzel langsam und an ihrem freien Ende intensiv. Das Wachstum am freien Ende nennt man appositionell. Eine dritte, besondere Art von Keilepithel findet sich am Boden der Neuralrinne.
Das konvergente Keilepithel arbeitet hier gegen besonders hohen Widerstand von Seiten des Stromas und gleichzeitig gegen Widerstand des seitlich angrenzenden Epi-thels. Die Basalmembran ist sehr dick. Die Dura entwickelt sich basal stark aus. Den Wachstumswiderstand vermögen die Zel-len durch eigene Wachstumsexpansion kaum zu überwinden. Sie bleiben auf en-gem Raum eingekeilt und enthalten wenig Flüssigkeit. Ihre Teilungsfähigkeit und da-mit ihr Flächenwachstum entlang des ventrikulären Flüssigkeitsraumes (Abb. 13, 1) erlöschen im zweiten Monat. Das emb-ryonale Rückenmark entwickelt sich nicht völlig eigenmächtig, sondern in engem funktionellem Zusammenhang mit dem
Bindegewebe seiner Umgebung. (Entwicklungs-funktionelle Untersuchungen am Nervensystem, Blechschmidt E., Zeitschrift für anat. Entwicklungsgeschichte, 1955, 119
(2):112-130). Dieser wachstumsfunktionelle Zusammenhang von neuralem Epithel und Bindegewebe wird im Kapitel „Gestaltungsfunktion des Gefässsystems“ und „Kranio-kaudale Krümmung“ noch eine spezielle Bedeutung bekommen.
V 6.2.3. Die Bedeutung der Flüssigkeitsräume Grenzgewebe liegt immer mit der einen Oberfläche zum Flüssigkeitsraum, mit der anderen zum Binnengewebe. Die Zellen sind alle orthogonal zu einem Lumen einge-stellt und lassen Teilchen senkrecht zu seiner Oberfläche permeieren, die des Bin-nengewebes auch parallel zum Grenzgewebe. Zellteilung erfolgt immer in unmittelba-rer Nähe zum Flüssigkeitsraum. Dort erfolgt also das Flächenwachstum – wie wir ge-sehen haben je nach Organanlage in spezifischer Wachstumsfunktion. Die Zellen aus der flüssigkeitsnahen Zellvermehrungszone (Abb. 14 a) wandern nun zur nah-rungsnahen Zellwachstumszone (Abb. 14 b). Hier erst erfolgt eine Volumenzunahme und die Körpergestaltung. Flächenwachstum ist die primäre Gestaltungsfunktion. Die Flüssigkeiten haben in diesem Zusammenhang einen wichtigen Anteil an der Kör-pergestaltung. Die Diathelzellen (in der deskriptiv-anatomischen Nomenklatur ent-sprechen sie den Epithelzellen) geben an der Grenze zum Flüssigkeitsraum hin ihre Abbaustoffe, vor allem Wasser, ab. Dadurch entlastet nehmen sie nun an der Kon-taktseite mit dem Stroma intensiv Nahrung auf, ohne wie die Zellen des wachsenden
Abb. 13

Embryologie aus Sicht der Craniosacralen Osteopathie 28/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Bindegewebes durch Stauung von Abbauproduk-ten im Inneren des Gewebes in Form von Interzel-lularsubstanz gehemmt zu werden. Formungs- und Faltungsvorgänge sind regelmässig Leistungen von Diathelien - nicht nur bei der Entstehung der Körperwand und damit der äusseren Körperform, sondern auch bei der Entwicklung der inneren Or-gane.
Abb. 14 Querschnitt eines 3.4 mm grossen Embryos. Punktiert: Flüssigkeitsraum bei 1 Neuralrohr, 2 So-mit, 3 Wolffscher Gang, 4 Leibeshöhle, 5 Darmrohr, 6 noch paarige Aorta dorsalis. Aussen: Flüssigkeit in der Chorionhöhle.
V 6.2.4. Binnengewebe Binnengewebe entsteht in Stoffwechselfeldern entlang von Grenzgewebe. Dort rü-cken die Zellen mehr und mehr auseinander, so dass ein netzförmiger Zellverband entsteht. In den Maschenlücken liegt viel flüssige Interzellularsubstanz. Die Interzel-lularsubstanz in Auflockerungsfeldern enthält wässerige Abbaustoffe. Die Flüssigkei-ten in den einzelnen Gewebevakuolen kommunizieren miteinander. Entlang der Ge-webevakuolen weisen die Zellen eine konkave Oberfläche auf. Der Flüssigkeitsdruck im Interstitium ist also grösser als jener des Zytoplasmas der Zellen. Die Zellen drif-ten divergent auseinander. So etwas ist im epithelialen Gewebe nicht möglich. Dort fehlen die geeigneten Interzellularräume. Die Flüssigkeit hat also immer eine wichtige Bedeutung bei der Gestaltung der Zell-verbände.
V 6.2.5. Die acht Stoffwechselfelder Wie oben erwähnt, gehören Lage-, Form- und Strukturentwicklung für die Entwick-lung eng zusammen. Die submikroskopischen Komponenten dieser Prozesse sind Bewegungen, die wir Stoffwechselbewegungen nennen. Die Stoffwechselbewegungen ihrerseits laufen in Stoffwechselfeldern geordnet ab. Erich Blechschmidt hat Stoffwechselfelder folgendermassen definiert: Das Stoffwechselfeld ist ein morphologisch abgrenzbarer Bereich mit räumlich ge-ordneten Stoffwechselbewegungen. Diese Bewegungen sind eine Grundeigenschaft des Entwicklungsgeschehens. Die Teilchenbewegungen in diesen Feldern erfolgt stets gegen Widerstand von Seiten ih-rer Umgebung und sind daher echte Arbeit im biophysikalischen Sinn (s. auch Kapitel

Embryologie aus Sicht der Craniosacralen Osteopathie 29/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Grenzgewebe). Die Entwicklung eines Menschen ist seit den ersten Entwicklungs-phasen, dynamisch und biologisch gesehen, eine individualspezifische Leistung. Blechschmidt hat acht Stoffwechselfelder beschrieben, die für verschiedene Körper-regionen charakteristisch sind:
1. Corrosionsfelder Sobald zwei Grenzgewebe eng miteinander in Kontakt sind, so dass kein ernährendes Binnengewebe mehr dazwischen liegt, erlischt die Nahrungszufuhr. Die mitein-ander in Kontakt stehenden Zellen gehen durch Nekrose zugrunde. Das ermöglicht den Austausch von Flüssigkeiten mit darüber liegendem Gewebe. Beispiele für solche Felder: Die beiden dorsalen Aortenäste verbinden sich zu einem gemeinsamen Stamm, während sich die Membran zwischen ihnen zurückbildet. Die Urnierkanälchen gewinnen durch Korrosion Anschluss an die Nierenkelche. Wo dies krankhafterweise nicht
stattfindet, entsteht eine sogenannte Cystenniere. Die embryonale Mundregion ist ebenfalls ein Korrosionsfeld. Hier drängen sich Ekto-derm und Entoderm ohne Binnengewebe eng aneinander. Es entsteht die Mundöff-nung.
2. Densationsfelder sind Verdichtungsfelder. Sie entstehen vorwiegend in der Tiefe der Binnengewebe. Wo biomechanisch ein Verlust an flüssiger Interzellularsub-stanz entsteht und die festen Partikel sich einander nähern, zeigt das Binnengewebe Verdichtungen. Die jungen Zellen werden in keiner bevorzugten Richtung auf Druck oder Zug
beansprucht. Sie sind daher kugelig und bilden die Anlage von Knorpelherden (Vor-knorpel). Der Arm ist ein gutes Beispiel für die Entstehung der Densation in der Ontogenese. Das Ektoderm bezieht die Nahrung aus dem unterliegenden Binnengewebe. Mit der Nahrungszufuhr entsteht hier im 2. Entwicklungsmonat ein dichtes Netz von Blutka-pillaren. Sie ernähren das Ektoderm ebenso wie das unmittelbar anliegende Stroma. Entsprechend dem osmotischen Druck in den Blutgefässen wird dem Binnengewebe Flüssigkeit abgesaugt. Dieses Densationsfeld ist das Entwicklungsareal des Armske-letts. Die Entwicklung der Luftröhre ist ein anderes anschauliches Beispiel. Das Epithel der Trachea ist auf der Rückseite dicker als auf der Vorderseite. Die an das Epithel an-grenzenden Zellen sind länger und tangential angeordnet; aus ihnen werden sich Trachealmuskeln und die Faserschicht entwickeln. Es wächst schneller als auf der Vorderseite. Hier werden die Zellen eher zusammengedrückt. Sie werden eher kuge-lig. Diese Zellen vermehren und verdichten sich zu Knorpelzellen. Das biokinetische Prinzip der Densationsfelder ist ausserdem für folgende Gebilde gültig: z.B. für die Rippen, die unter dem Wachstumsdruck der Herz- und Lebermas-se entstehen oder für das Nasenseptum.

Embryologie aus Sicht der Craniosacralen Osteopathie 30/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
3. Contusionsfelder entwickeln sich im Innern von Densationsfeldern, wenn eine biomechanische Stauchung stattfindet. Während ein Densationsfeld eine Zone ist, in der abgerundete Zellen verdichtet werden, ist ein Contusionsfeld eine Zone, in der die abgerundeten Zellen so aneinander gedrängt werden, dass sie sich tellerförmig
abplatten. Diese Tellerform ist typisch für junge Knorpelzellen, also Vorknorpelzellen. Ein Beispiel für ein Contusionsfeld ist die Anlage der Wirbelsäule (Abb. 15). Blechschmidt’s Untersuchungen haben ergeben, dass es das Rückenmark ist, das das Contusionsfeld für die Wirbelsäulenanlage initiiert. Die Seitenwand des Wirbel-kanals wird vom wachsenden Rückenmark und seinem flüssigkeitsreichen Bett zircu-lär gestrafft.
Das gestraffte Gewebe ist die sogenannt harte Rückenmarkshaut oder Dura spinalis, die zu-nächst ventral besonders kräftig ist. Ventral wird sie vom wachsenden Rückenmark gestrafft. Der Krümmungsradius nimmt zu, die Dura flacht sich ab. Mit der Abflachung werden die Zellen an der Aussenseite der Dura gestaucht (Abb. 15 konvergentes Pfeilpaar mit Querstrichen deutet das entstehende Contusionsfeld 1 an).
Der Pfeilkopf in Abb. 16 zeigt die Richtung an, die der Flüssig-keitsdruck der Interzellularsubstanz in der Arachnoidea (Spinnwebehaut) auf die dorsal und seitlich dünne Dura aus-übt. Konturierte Pfeile zeigen die Wachstumsbewegungen des jeweils jüngsten Teils der Dura. Die Dura begradigt sich zu-nehmend. Die Haltefunktion der ventral bereits gestrafften, zugfesten Dura ist mit konvergenten Doppelpfeilen beschrie-ben. Das konvergente Pfeilpaar mit Querstrichen umschreibt die Contusion der Zellen an der Aussenseite der Dura.
4. Distusionsfeld ist ebenfalls ein Stoffwechselfeld, wie es sich im Skelettsystem findet. Durch gestaute noch hinreichend grossmolekulare Abbau-produkte bekommen Knorpelzellen einen hohen osmoti-schen Druck, so dass Wasser aus der Umgebung ein-strömt. Die jungen Knorpelzellen quellen und zeigen damit ein sogenanntes Quellungswachstum. Damit üben sie eine
Stemmkörperfunktion in bevorzugter Richtung aus, z.B. bei der Fingeranlage parallel zur Längsachse.
Abb. 15
Abb. 16

Embryologie aus Sicht der Craniosacralen Osteopathie 31/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Im Hinblick auf die genannte Stemmkörperfunktion stellt das knorpelige Skelett und nicht etwa die Muskulatur den ersten aktiven Teil des Bewegungsapparates dar. Die Muskulatur gewinnt erst nach ihrem Dehnungswachstum die Kontraktionsfähigkeit.
5. Retensionsfeld entsteht dort, wo Binnengewebe durch einen Wachstumszug gestrafft wird. Das ist so zu verstehen, dass eine Ansammlung von inneren Gewebe-zellen, die zunächst undifferenziert waren, langsamer in ei-ne bestimmte Richtung wachsen als das umliegende Ge-webe. Der Zellverband leistet mit einem langsamen Wachs-
tum einen biomechanischen Zugwiderstand. Der Wachstumswiderstand führt zur Straffung des Gewebes. Als Merkmal davon werden die Zellen spindelförmig und die Zellkerne schlank. Was also schon entwicklungskinetisch gestrafft wurde, wird später als Halteapparat funktionieren. Auf diese Weise entstehen Sehnen, Bänder und Fas-zien. Weitere Beispiele für Retensionsfelder sind die bindegewebige Leitstruktur von Blutgefässen oder der zentrale Teil des Zwerchfells. Dort kommen das rasch wach-sende Herz und die sich schnell vergrössernde Leber einander so nahe, dass das Bindegewebe in dem Raum zwischen ihnen komprimiert und gleichzeitig von seinem Rand aus gestrafft wird.
6. Dilationsfeld. Man spricht von Di - lationsfeld, um zu betonen, dass es sich nicht um einen physikalischen Begriff handelt (rein mechanischer Zug oder Dehnung), wohl aber um einen biomechanischen Zug, also Zugwir-kungen als Leistung lebender Zellen und Zellverbände. In diesen Feldern werden Bindegewebszellen passiv gedehnt.
Sie geben ohne grossen Widerstand nach und werden dabei dünner. Solche Zellen entwickeln sich zu Muskelzellen. Ihre Dehnung ist entwicklungsdynamisch ein passi-ves Geschehen. Muskeln sind daher zuerst der passive Teil des Bewegungsappara-tes - im Gegensatz zum Knorpel, der mit seinem Quellungswachstum zuerst eine ak-tive Stemmkörperfunktion auf den Bewegungsapparat ausübt. Die Muskeln durchlau-fen also zuerst eine passive Entwicklungsdynamik, bevor sie die Fähigkeit zur akti-ven Verkürzung bekommen (s. auch Kapitel VII 1.4. Bildung der Somiten, Myotome).
7. Detraktionsfeld. Hier wird ein Zellverband unter Kompression auf einer harten Unterlage entlanggescho-ben. Dabei wird im Nahtgebiet Flüssigkeit ausgepresst. Als Folge davon verhärtet sich die Substanz. Solcherart durch biomechanische Gleitbewegungen verdichtete Zonen sind Entstehungsorte von Knochen. Jede Grundlage für Kno-chenbildung, ob Bindegewebe oder Knorpel oder bereits
verknöcherte Grundlage, die weiterer Knochenbildung dient, beginnt als Gewebever-dichtung mit Wasserverlust und nachfolgender Imprägnation mit schwer löslichen Calciumverbindungen. Es gibt eine Vielzahl von möglichen Formen von Knochenherden. Jede macht eine charakteristische kinetische Entwicklung durch, die immer von einer Ausdehnung un-ter dem Druck von Interzellularsubstanz begleitet ist. Ein Beispiel: Mit dem Längerwerden des Gesichts vergrössert sich der Abstand zwi-schen Gehirn und dem noch bindegewebigen Jochbogen. Er ist seitlich am Rand der

Embryologie aus Sicht der Craniosacralen Osteopathie 32/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Augenhöhle geknickt. Hier übt der Jochbogen mit der Abstandsvergrösserung zwi-schen ihm und dem Gehirn über das Bindegewebe der Schläfe einen Wachstumszug auf die gestraffte Hirnhaut aus. Durch diesen Zug wird die Dura wachstumsdyna-misch in besonderer Weise beansprucht. Mit der Zugbeanspruchung wird die äusse-re Schicht der Dura von der inneren Schicht lokal abgehoben. In der Zone der Abhe-bung kommt es zu einer Wasserverschiebung. Die Wasserverschiebung bedeutet ei-ne Entmischung mit lokaler Verfestigung zwischen den Durablättern. Das nunmehr verdichtete Gebiet ist das Entstehungsgebiet eines Knochenkerns, des knöchernen Stirnbeines.
8. Sogfeld ist eine Zone entlang von innerem oder äusserem Grenzgewebe, in der während des Wachstums biomechanisch ein Sog entsteht. Solche Zonen entwickeln sich, wenn sich das Grenzgewebe von dem langsamer wachsenden Binnengewebe abhebt. Dadurch kann Flüs-sigkeit einfliessen. Die einfliessende Flüssigkeit lockert den Zellverband des Binnengewebes auf. Wo die Menge der In-
terzellularsubstanz zunimmt, verschmelzen die Bläschen miteinander. In ein derartig aufgelockertes Gewebe können Epithelzellen aus der Nachbarschaft einsprossen, Stoffe aus der Flüssigkeit aufnehmen und sich auf diese Weise Platz für ihr Vor-wachsen schaffen. Solche Epithelsprossen kennen wir als Anlage von Drüsen. Die grössten Sogfelder stellen die Leber und die Lunge dar. Zu Beginn der Entwicklung handelt es sich bei diesen aufgelockerten Feldern im In-neren des Mesoderms um Vorstufen der Gefässbildung.
V 6.2.6. Kinetische Entwicklungstheorie und der Primäre Atemmechanismus
Folgende Gesetzmässigkeiten, wie Erich Blechschmidt sie für Entwicklungs- und Wachstumsbewegung und auch für Stoffwechselvorgänge in ihren Grundzügen be-schrieben hat, möchte ich hier nochmals hervorheben: • Die Grundfunktion der Organe ist die Gestaltungsfunktion. • Jeder Zellverband hat Gestaltungsfunktion. • Gestaltungsvorgänge sind Bewegungsabläufe. • Zellteilung erfolgt immer in unmittelbarer Nähe zum Flüssigkeitsraum. Dort erfolgt
also das Flächenwachstum. Flächenwachstum ist die primäre Gestaltungsfunkti-on. Das gilt auch für die Entwicklung des Neuralrohrs um den mit Liquor gefüllten Zentralkanal.
• Die Zellen eines Gewebes sind stets kinetisch durch Materialbewegungen mitein-ander verbunden. Die Zellen nehmen Nahrung aus den Zwischenzellspalten und aus ihren Nachbarzellen auf und ziehen sich durch diese Materialaufnahme ge-genseitig an. Andererseits stossen sie sich durch Abgabe von verbrauchten Stof-fen gegenseitig ab. Das Wechselspiel zwischen Aufnahme und Abgabe ist die Voraussetzung dafür, dass die Zellen sich in bestimmter Weise anordnen, eine bestimmte Form annehmen und in Form halten.
• Der Embryo wächst von Anfang an durch menschlich spezifische Stoffwechsel-vorgänge, die er gegen Widerstand, also durch aktive Arbeit leistet.

Embryologie aus Sicht der Craniosacralen Osteopathie 33/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
In den nächsten Kapiteln werden diese Gesetzmässigkeiten im Zusammenhang mit der eigentlichen Organogenese verständlicher werden. In der Organogenese von besonderer Bedeutung im Zusammenhang mit meiner Fragestellung ist die Entwick-lung des Nervensystems und der inneren Liquorräume (Ventrikelsystem). Sie haben für den Primären Atemmechanismus die Bedeutung des Motors. Ihnen ist daher das nächste Kapitel V gewidmet. Die Entwicklung des Nervensystems kann nur in engster, wechselseitiger Abhängig-keit mit derjenigen des Herz/Gefässsystems gesehen werden. Dieser Tatsache ist das Kapitel VII, 2. gewidmet. Im gleichen Kapitel entwickle ich auch die These: die wechselseitige Entwicklungsabhängigkeit von Gehirn- und Rückenmarksanlage zur Herz-/Gefässanlage bleibt als weiter bestehende inhärente Eigenbewegung des zentralen Nervensystems als Antriebsmotor des Primären Atemmechanismus beste-hen. Die übrigen Organe können leider im Rahmen dieser Arbeit, wenn überhaupt, nur kurz erwähnt werden.

Embryologie aus Sicht der Craniosacralen Osteopathie 34/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VI Die embryonale Entwicklung des Nervensystems
VI 1. Erste Leistungen des Nervensystems sind Wachstumsleistungen Erich Blechschmidt beschreibt die Entwicklung des Nervensystems als seine erste Leistung, d.h. als Wachstumsleistung. Diese frühe Tätigkeit, schon zur Zeit ihrer Ent-stehung, ist die Grundfunktion von Neuralrohr, Gehirn und Rückenmark in der emb-ryonalen Phase. Sie leitet als Wachstumsfunktion die späteren Leistungen des Ner-vensystems ein. Blechschmidt geht soweit, dass er die nachgeburtlichen Funktionen des Nervensystems im Vergleich zu den embryonalen komplexen und vielschichtigen Wachstumsfunktionen als nur noch eng spezialisierte Funktionen im Sinne von Rest-leistungen einordnet.
VI 2. Die Bildung des Neuralrohrs Die Entwicklung des Nervensystems nimmt seinen Anfang mit der Bildung der Neu-ralplatte. Über der Chorda (vgl. Kapitel VII, 1.3.) und dem paraxialen Mesoderm ver-dickt sich das Ektoderm zur Neuralplatte.
Das Neuralrohr entsteht aus der Neuralplatte. Das dicke Ektoderm bildet entlang der Neuralrinne viele Tochterzel-len. Diese wandern der Nah-rungsquelle in der Tiefe der Ektodermschicht entgegen. Hier wachsen sie zu neuen teilungsfähigen Zellen heran. Das wachsende Neurale-pithel zeigt entlang der basa-len Fläche des Ektoderms schnellere Oberflächenver-grösserung als im Kontakt mit der Amnionflüssigkeit. Dabei entsteht eine Halte-funktion Abb. 17, 2). Die Neuralrinne vertieft sich und schliesst sich zum Neural-rohr.
1) Flächenwachstum des Ektoderms . 2) Haltefunktion im Bereich der freien Oberfläche des Ektoderms (Kontaktfläche mit der Amnionflüssigkeit) 3) Haltefunktion im Bereich des Axialfortsatzes/Chordafortsatz.
Abb. 17

Embryologie aus Sicht der Craniosacralen Osteopathie 35/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VI 2.1. Wachstumsleistungen des Neuralrohrs Die Wand des Neuralrohrs bildet im ersten Monat einen epithelialen Zellverband zwi-schen der Neuralrohrflüssigkeit im Zentralkanal einerseits und seinem Gefässführen-den Bett andererseits. Das Neuralrohr ist ein Bildungsfaktor der primitiven Meningen und damit indirekt ein wichtiger Formator des Achsenskeletts. Die Umgebung des Rückenmarks ist wäh-rend seiner Entwicklung in ein dichtes Gefässnetz eingeschlossen, diese Region wird zur Pia. Peripher davon entwickelt sich in der gefässarmen Zone des Bindegewebes ein flüssigkeitsreicher Hof um das piabedeckte Rückenmark. Dieser Hof ist die Anla-ge der Arachnoidea. Das Volumen der Arachnoidea nimmt regelmässig zunächst ventral besonders zu. Dadurch hebt sie das Rückenmark von der Basis ab, so dass es bei Flexion des Rückens nicht gegen die feste Wirbelsäule gepresst wird. (Zeitschrift
für anatomische Entwicklungsgeschichte, 1955, 119 (2); 112 – 130) Eine weitere Folge der Arachnoideabil-dung ist die Abplattung der darüber liegenden Zellen, die sich zur Dura entwickeln, indem sie viel Flüssigkeit ausschwitzen. Die Dura wird ein straffes Gewebe (vgl. Ka-pitel V, 6.2.5. Stoffwechselfeld, Contusionsfeld). Die Begleitbindegewebe des wach-senden Rückenmarks sind dorsal, lateral und ventral verschieden differenziert (Vgl. Kapitel VII, 1.4. Bildung und Auflösung der Somatomere). Ventral werden sie in die Bildung des Achsenskeletts einbezogen. Nur aussen hat das Neuralrohr Kontakt mit den Nahrungsstoffen, die das Blut zu-führt. Die Zellkörper und Zellkerne liegen in der Tiefe, also nahe am Zentralkanal. Die Zellen lassen auf kürzestem Weg Fortsätze nach aussen zur nahrungsreichen Schicht wachsen. So stehen die Zellfortsätze also regelmässig senkrecht zur Ober-fläche des Neuralrohrs. Die Zellfortsätze sind an der Vergrösserung des Neuralroh-rumfanges unmittelbar beteiligt, sobald sie Nahrung aus den Blutgefässen aufneh-men. Sie leisten die Hauptwachstumsarbeit an der Aussenseite des Neuralrohrs. In der inneren Schicht, nahe dem Ventrikellumen, findet nur Zellteilung statt. Blechschmidt nimmt an, dass die Zellen bei der Mitose Flüssigkeit aus dem Zentral-kanal aufnehmen. Sobald der Umfang des Neuralrohrs zunimmt, wandern Zellen aus der tiefen Schicht peripherwärts und bilden zwischen weisser und schwarzer Zone die erste sogenannt graue Substanz. Das gilt für das Neuralrohr sowohl im Rücken- wie auch im Kopfge-biet. Nirgendwo sammelt sich Interzellularsubstanz an. Die Gliazellen dienen den Neuroblasten als Nahrungs- und Leitstrukturen. Sie wach-sen von der Kontaktseite mit dem Mesenchymbett her ins Neuralrohr ein. Das Neuralrohr ist ventral, wo es der mesodermalen Schicht anliegt, im Wachstum behindert (vgl. Abb. 8). Es wächst seitlich und dorsal schneller. Sein Wachstum findet hier weniger Widerstand. Die Entocystscheibe ist oval und nicht kreisrund. Von dieser Form wird auch die Ent-wicklung des Neuralrohrs mitbestimmt. So hat das Neuralrohr ein breiteres und ein schmaleres Ende. Sein Wachstum ist am Kopfende in alle Richtungen freier als im Abschnitt, wo Somiten, Wirbelsäulen- und Aortenanlage es umgeben.

Embryologie aus Sicht der Craniosacralen Osteopathie 36/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VI 2.2. Wachstumsleistungen von Gehirn und Rückenmark Gegen Ende des ersten Entwicklungsmonats zeigt das dickere Ende des Neuralrohrs in primitiver Weise bereits zahlreiche Formmerkmale des späteren Gehirns. Die Grenze zwischen Rückenmark und Gehirn ist noch unscharf. Das Rückenmark ist im Verhältnis zum Gehirn noch sehr klein. Legende zu Abb. 18 (26. Tag) A Haltefunktion des Bindegewebes und der Visceralbogengefässe in den Beugefal-ten (Viszeralbögen). Zwischen ihnen ist das Entoderm des Kopfdarms als Schlundta-sche ausgestülpt und Ektoderm als Halskerben eingezogen.
B Wachstumsdruck des Neuralrohrs C Wachstumsbewegung (Biegung) des Gehirns D Fortsetzung der Aorta 1) 1. – 3. Visceralbogen 2) Vorderhirn mit Augen-blase 3) Mittelhirn, bildet Schei-telbeuge 4) noch langes Hinterhirn mit Ohrblase 5) obere Cardinalvene 6) Einmündung in das Herz 7) Grosshirnzügel, gestraff-tes Bindegewebe in der Verlängerung der Aorta Das Kopfende weitet die
häutige Kopfwand an seinem breiten Ende und ascendiert gegenüber dem Einge-weidetrakt. Das Nahrung spendende Aortensystem bleibt im Vergleich zu dem Nah-rung aufnehmenden Neuralrohr kurz und wird so zum Halteapparat für Letzteres. Einmal im Nabel (Nabelring) verankert, übt das Gefässsystem mit seiner Gestal-tungsfunktion unmittelbar Einfluss auf das Wachstum des Neuralrohrs und des Ner-vensystems aus. Sobald das Neuralrohr durch zahlreiche Äste der Aorten an den grossen Gefäss-stämmen fester verankert ist, passt es sich während des weiteren Wachstums des Embryo durch eine zunehmende dorsokonvexe Krümmung den kurz bleibenden Aor-ten an. Das Herz entfernt sich mit zunehmender Krümmung vom Gehirn und des-zendiert im Verhältnis zum Gehirn. Es gibt das Hirn damit kranial mehr und mehr frei zu einer relativ selbständigen Entwicklung. Der Kopfteil des Neuralrohrs wird zuneh-mend mitosenreicher. Der Descensus des Herzens lässt dem Hirn immer mehr Raum. Die das wachsende Hirn versorgenden Blutgefässe spreizen sich von der Anlage der Schädelbasis aus peripherwärts. Dabei schwitzt das wachstumsleistende Gehirn Liquor in das umge-bende Mesenchym aus. Mit der Volumenzunahme der nun sogenannten Arachnoi-dea strafft sich das ihr peripher angrenzende Bindegewebe und bildet die frühembry-onale basale Dura (vgl. Kapitel V 6.2.5. Stoffwechselfelder, Contusionsfeld). Nahe
Abb. 18

Embryologie aus Sicht der Craniosacralen Osteopathie 37/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
der basalen Duraanlage bleiben die herznahen Abschnitte der Hirngefässe als kräfti-ge arterielle Gefässstämme des Gehirns kurz im Verhältnis zum antibasalen weitläu-fig vernetzten und dünnwandigen vor allem venösen Gefässplexus. Die Dura strafft sich zunächst in den kurz gebliebenen basalen Abschnitten, wo sie wahrscheinlich die Pulsstösse der besonders kräftigen Schlagadern des Gehirns auf-fangen muss. Das gestraffte Gewebe stellt den Hauptteil der desmalen Anlage des Schädels dar. Ein Teil der desmalen Schädelanlage setzt rechts und links vom Gehirn das Mesen-chymbett der paarigen Aortenanlage in einer geraden Verlängerung bis zur antibasa-len Konvexität der Gehirnanlage fort. Diese Fortsetzung bildet hier die sogenannten Grosshirnzügel (Abb. 18, 7) Sie schnüren das Neuralrohr an seinen Flanken über der Anlage der Mundspalte von rechts nach links ein. An dieser taillenförmigen Einschnü-rung grenzt sich allmählich das Diencephalon gegen die Grosshirnhemisphären ab. Die medialen Seiten der Hemisphären gehen mit den einwachsenden Blutgefässen eine enge Verbindung ein – die Plexus choroidei stülpen sich in die Ventrikel ein. Dorsal davon bildet das Mittelhirn die Scheitelbeuge. Der mit dem Gesicht in Verbin-dung stehende Hirnabschnitt zwischen Mittelhirn und Rückenmark erscheint als Hin-terhirn. Es wird zum eigentlichen Gesichtshirn.
Die grossen Hirnnerven N. trigemi-nus, facialis, glossopharyngeus und vagus sind in den Visceralbögen 1,2,3 und 4 verankert. Die Visceral-bögen sind beim Menschen die ers-ten oberflächenvergrössernden Teile der Gesichtsregion. Wegen dieser Verankerung vermag das Hirn hier im Laufe des weiteren Längenwachs-tums nur teilweise vom Gesicht und von der Schädelbasis abzurücken. Der lokal besonders fest verankerte Gehirnabschnitt entwickelt sich zur Brücke (Pons). Nur vor und hinter der Brücke wölbt sich das Gehirn noch längere Zeit stark antibasalwärts vor, so dass es hier zur Ausbildung der Scheitel- und Nackenbeuge kommt (Abb. 21) Damit ist eine erste Gliederung des Gehirns beschrieben, die noch keine funktionelle Differenzierung aufzeigt, sondern die durch integrierte Wachs-tumsvorgänge hervorgebrachte
wachstumsfunktionelle Bildung beschreibt. Das Gehirn ist im 2. Monat immer noch ein weitlumiges Hohlorgan, das sich in Fal-tung befindet und dabei an mehreren Stellen knickt. Das Lumen, also die Ventrike-lanlage des noch deutlich rohrförmigen Gehirns ist örtlich verschieden weit. Die Dura wird durch das Expansionswachstum des Gehirns gestrafft. Sie setzt dieser Entwicklung zunehmenden Widerstand entgegen, indem sie eine relativ zum Gehirn-volumen kleine Oberfläche anstrebt und sich ihrer Gestalt, der Kugelform, nähert.
Abb. 19
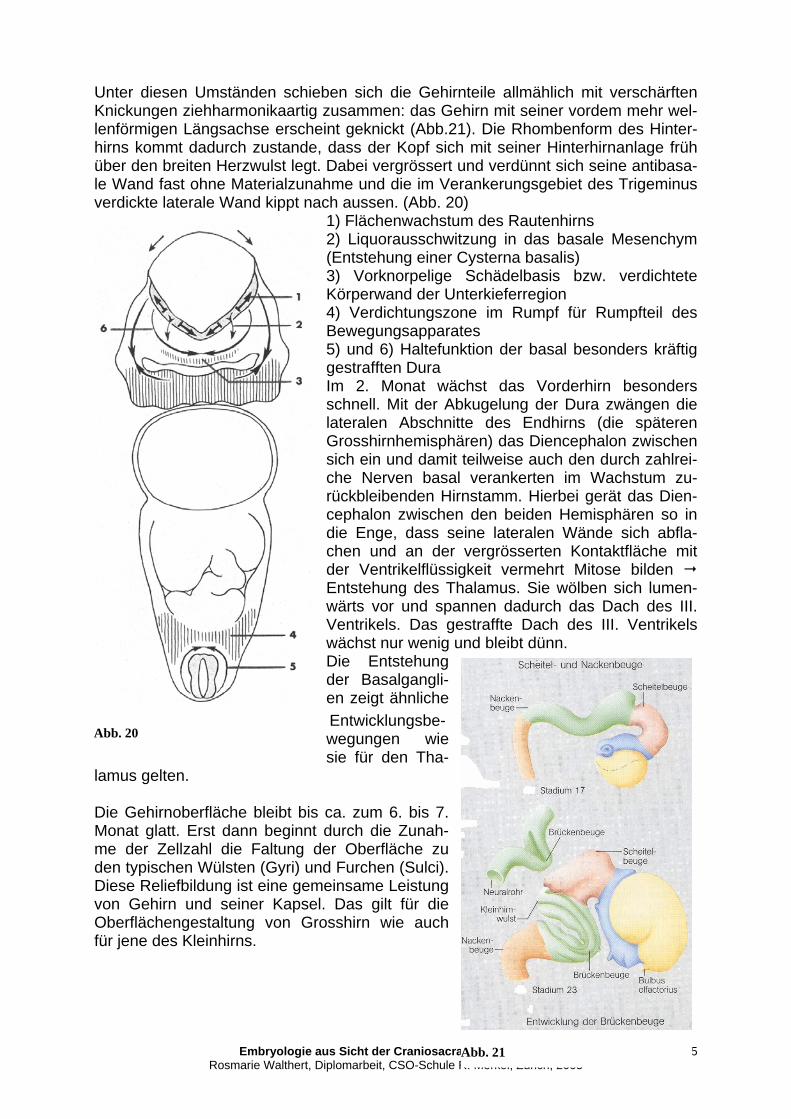
Embryologie aus Sicht der Craniosacralen Osteopathie 38/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Unter diesen Umständen schieben sich die Gehirnteile allmählich mit verschärften Knickungen ziehharmonikaartig zusammen: das Gehirn mit seiner vordem mehr wel-lenförmigen Längsachse erscheint geknickt (Abb.21). Die Rhombenform des Hinter-hirns kommt dadurch zustande, dass der Kopf sich mit seiner Hinterhirnanlage früh über den breiten Herzwulst legt. Dabei vergrössert und verdünnt sich seine antibasa-le Wand fast ohne Materialzunahme und die im Verankerungsgebiet des Trigeminus verdickte laterale Wand kippt nach aussen. (Abb. 20)
1) Flächenwachstum des Rautenhirns 2) Liquorausschwitzung in das basale Mesenchym (Entstehung einer Cysterna basalis) 3) Vorknorpelige Schädelbasis bzw. verdichtete Körperwand der Unterkieferregion 4) Verdichtungszone im Rumpf für Rumpfteil des Bewegungsapparates 5) und 6) Haltefunktion der basal besonders kräftig gestrafften Dura Im 2. Monat wächst das Vorderhirn besonders schnell. Mit der Abkugelung der Dura zwängen die lateralen Abschnitte des Endhirns (die späteren Grosshirnhemisphären) das Diencephalon zwischen sich ein und damit teilweise auch den durch zahlrei-che Nerven basal verankerten im Wachstum zu-rückbleibenden Hirnstamm. Hierbei gerät das Dien-cephalon zwischen den beiden Hemisphären so in die Enge, dass seine lateralen Wände sich abfla-chen und an der vergrösserten Kontaktfläche mit der Ventrikelflüssigkeit vermehrt Mitose bilden Entstehung des Thalamus. Sie wölben sich lumen-wärts vor und spannen dadurch das Dach des III. Ventrikels. Das gestraffte Dach des III. Ventrikels wächst nur wenig und bleibt dünn. Die Entstehung der Basalgangli-en zeigt ähnliche Entwicklungsbe-wegungen wie sie für den Tha-
lamus gelten. Die Gehirnoberfläche bleibt bis ca. zum 6. bis 7. Monat glatt. Erst dann beginnt durch die Zunah-me der Zellzahl die Faltung der Oberfläche zu den typischen Wülsten (Gyri) und Furchen (Sulci). Diese Reliefbildung ist eine gemeinsame Leistung von Gehirn und seiner Kapsel. Das gilt für die Oberflächengestaltung von Grosshirn wie auch für jene des Kleinhirns.
Abb. 20
Abb. 21

Embryologie aus Sicht der Craniosacralen Osteopathie 39/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VI 2.3. Gliederung des Nervensystems Faktoren vom dorsalen Ektoderm und solche von der ventralen Chorda dorsalis aus-gehend bestimmen die weitere Entwicklung des Neuralrohrs und induzieren eine un-terschiedliche Differenzierung der Neuronen zu motorischer Funktion ventral (Grund-platte) und dorsal mit sensibler Funktion (Flügelplatte). Vor der Entwicklung der Gehirnbläschen ist im Neuralrohr des Kopfes eine metamere Gliederung zu erkennen, die der Gliederung des Rumpfes in Somitensegmente ent-spricht. Diese Segmente des Neuralrohrs werden als Neuromere bezeichnet und spiegeln die primäre Gliederung der Kopfanlage wider, während mit der Ausbildung der Gehirnbläschen bereits die funktionelle Ausgestaltung beginnt. Die drei primären Hirnbläschen sind Zentren für die zentralen Sinnesorgane Riechen, Sehen und Hören. Aus dem Prosenzephalon stülpen sich nach lateral die Augenblä-schen aus, die im Oberflächenektoderm die Augenplakoden induzieren. Im Ektoderm des Stirnfortsatzes induziert das Prosenzephalon die Riechplakoden. Das zweite Bläschen entspricht dem späteren Mesenzephalon (Mittelhirn). Das dritte Gehirnblä-schen ist das Rhombenzephalon (Rautenhirn). Ihm sind beiderseits die Ohrbläschen zugeordnet, die sich durch Einstülpung der Ohrplakoden aus dem Ektoderm entwi-ckeln.
Abb. 22
Durch das Auswachsen der Endhirnbläschen wird das ursprüngliche Prosenzephalon als Dienzephalon (Zwischenhirn) zwischen die Endhirnhemisphären eingeschlossen. Die Endhirnbläschen werden zum I. und II. Seitenventrikel, das unpaare Vorderhirn-bläschen zum III. Ventrikel im Dienzephalon. Die Lamina terminalis wird als vordere

Embryologie aus Sicht der Craniosacralen Osteopathie 40/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Begrenzung des Prosenzephalons dem Telenzephalon zugerechnet. Die Endhirnblä-schen stellen am Anfang nur ein dünnes Häutchen dar. Aber schon von der 5. Wo-che an beginnen sie mit der Entwicklung der Grosshirnrinde. Telezenphalon: Die Endhirnhemisphären gehen am Foramen interventriculare vom Zwischenhirn ab und überwachsen schliesslich den gesamten Hirnstamm. Dienzephalon: Es ist primär ein Sehhirn, aus dem sich die Augenbläschen ausstül-pen. Mesenzephalon: Hier entwickelt sich die Vierhügelplatte. In den oberen Vierhügeln wird optische Information, in den unteren Hör- und Gleichgewichtsinformation zu Hal-tungs- und Blickbewegungen verarbeitet. Das Mittelhirnbläschen verengt sich zum Aquaeductus cerebri. Rhombenzephalon:
a) Myelenzephalon (hinterer Abschnitt): Es kann als verlängertes Rückenmark aufgefasst werden. Es enthält als Schlundhirn die Kerne für die Schlundbo-gennerven des ersten bis fünften Schlundbogens.
b) Metenzephalon (vorderer, cranialer Abschnitt):Es ist der letzte Hirnabschnitt oberhalb des Rückenmarks. Es enthält die zentrale Repräsentation des Hör- und Gleichgewichtsorgans. Bei der Einknickung des Rautenhirns zur Brü-ckenbeuge werden die Rautenlippen quergestellt und bilden einen Wulst, der sich in den IV. Ventrikel hinein und über dem Dach der Rautengrube aus-dehnt. Die Kleinhirnwülste entstehen in den oberen Lippen der Rautengrube. Das Kleinhirn dient der Feinabstimmung von Haltungs- und Stellreflexen.
Abb. 24
Abb. 23

Embryologie aus Sicht der Craniosacralen Osteopathie 41/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII Der Primäre Atemmechanismus und seine Entstehung in der embryonalen Entwicklungsphase
Der Primäre Atemmechanismus findet seine Anfänge im Embryo auf mindestens zwei verschiedenen Ebenen, nämlich auf einer energetischen und auf einer bewe-gungsmässigen. Immer wieder spannend dabei sind die Ausführungen von Erich Blechschmidt. Für meinen Wissensstand wird er mit seinen submikroskopischen Forschungen den grobstofflichen Verhältnissen mehr als gerecht. Aber auch den feinstofflichen, ja so-gar den energetischen Überlegungen kommt er mit seinen Ausführungen erstaunlich nahe. Manchmal kann man sich fragen, was er von traditionellen Medizinkulturen aus dem fernen Osten gewusst haben mochte.
VII 1. Gestaltende Energie oder „breath of life“ Sutherland hat vom Primären Atemmechanismus auch als „Breath of life“ – Lebens-atem - gesprochen. Ausserdem hat er der Fluktuation der zerebrospinalen Flüssigkeit eine energetische Qualität zugeordnet, die Potency oder Intelligency (Anfangsbuch-stabe mit einem grossen I geschrieben). Diese energetische Ebene habe ich im Werdeprozess der axialen Organanlagen gesucht – im Organisationszentrum Primi-tivrinne und -knoten. Interessant in diesem Zusammenhang ist Erich Blechschmidt’s Betrachtungsweise des Axialfortsatzes. Seine Forschungsergebnisse an fast lebensfrischen Embryonen stellen nicht nur die Wachstumsentwicklung von Primitivstreifen und Axialfort-satz in lage- und bewegungsmässigen Zusammenhang. Sie sprechen ausserdem dem Axialforsatz eine ganz spezifische Eigenschaft zu, nämlich die Eigenschaft des ruhenden Pols in einer extrem komplexen und dichten Entwicklungsphase. Die Vorherrschaft des Gehirns lässt sich schon nach den ersten beiden Wochen er-kennen. Schon hier ist die Entocystscheibe am cranialen Ende breiter als am cauda-len Pol. Hier wirkt der Haftstil einengend und wachstumsbremsend, während am an-deren Kopfende schnelles Wachstum gegen geringen Widerstand stattfindet. In der Kopfregion wölbt sich das Ektoderm in den Fruchtwasserraum vor und bildet eine Expansionskuppe. Im Rumpfteil der Entocystscheibe entsteht im Gegensatz zur Ex-pansionskuppe eine Senke – die Impansionssenke. Die Expansionskuppe geht mit einem scharfen Rand in die Impansionssenke über. Der wachsende Umbördelungs-rand überrollt mehr und mehr die Impansionssenke, so dass eine fingerförmige Ein-stülpung entsteht, der sogenannte Axialfortsatz. Während sich der wachsende Um-bördelungsrand immer weiter über die Impansionssenke schiebt, findet im Inneren des Axialfortsatzes kaum ein Wachstum statt. Blechschmidt betrachtet deshalb die Spitze des Axialfortsatzes als das Zentrum, gleichsam den Nullpunkt der Ent-wicklungsbewegung der ganzen Keimscheibe. Sehr bald verschwindet das Lumen des Axialfortsatzes. Was bleibt, ist die Chorda.

Embryologie aus Sicht der Craniosacralen Osteopathie 42/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 1.1. Anfänge des energetischen Aspektes Die Anfänge des energetischen Aspektes können in der Anlage der ventralen Mittel-linie des Embryos, in der Chorda dorsalis gesucht werden. Ab dem 15. Tag ist am hinteren Pol des Keimlings der Primitivstreifen als verdicktes Ektoderm sichtbar. Der Streifen wächst bis zum vorderen Drittel der Keimscheibe vor. Auf dem Streifen entsteht die Primitivrinne, durch die sich am 16./17. Tag Zell-material einrollt und so die Mesodermschicht bildet. Wenn der Primitivstreifen seinen Endpunkt zwischen der Mitte und dem vorderen Drittel der Keimscheibe erreicht hat, verdickt sich seine Spitze zum Primitivknoten, durch den sich das Material für den Chordafortsatz einschiebt. Am 16./17. Tag beginnt in der Mitte der Keimscheibe durch Zellbewegungen und Zellteilungen die Entwicklung des Embryonalkörpers. Zellen wandern nach lateral der Keimscheibe, ordnen sich als paraxiales und intermediäres Mesoderm und verbinden sich am Rande der Keimscheibe in Form der dünnen Seitenplatte mit dem Mesoderm der Hüllen (parietale Mesodermschicht mit Amnion und viszerale Mesodermschicht mit dem Dottersack). Im Inneren der zweiblättrigen Keimscheibe gibt es also jetzt ei-ne dritte Schicht: das Mesoderm. Zellen, die sich durch den Primitivknoten ein-schieben, wandern als Kopffortsatz auf die Prächordalplatte zu.
Abb. 25 Aufsicht auf die Keimscheibe mit Primitivrinne und Primitivknoten
Heute weiss man, dass jede Schicht (Entoderm, Mesoderm, Ektoderm) einen be-stimmten Beitrag zu ganz verschiedenen Geweben und Organen leisten kann. Jede Keimschicht wird sich mit der benachbarten Schicht vereinigen und ver-binden. Sie werden sich gegenseitig durchdringen, um zu wachsen und auch, um die verschiedenen Teile des menschlichen Körpers mitzubilden. Das Ektoderm liefert die Anlagen für alles, was später mit Informationsprozessen zu tun hat (Nervensystem, Sinnesorgane). Es überzieht den Körper und bildet die Epi-dermis (auch Haare, Nägel, Hautdrüsen, Zahnschmelz).

Embryologie aus Sicht der Craniosacralen Osteopathie 43/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Das Mesoderm stellt das Material für die inneren und äusseren Bewegungsprozesse (Zirkulation, Muskulatur, Bewegungsorgane usw.). Es bildet als Mesenchym embryo-nales Bindegewebe und leistet seinen Beitrag zu verschiedenen Typen von Binde-gewebe: so z.B. Blut, Knorpel-, Knochen- und Muskelgewebe, Aufbau der Blutgefäs-se, der Körperhöhlen (Perikard, Pleura, Peritoneum), der Nieren und der Gonaden, Muskel- und Bindegewebsschichten des Verdauungssystems. Thymus. Das Entoderm stellt das Material für die Stoffwechselorgane. Es liefert die Ausklei-dung eines Grossteils des Verdauungskanals und der mit ihm vergesellschafteten Drüsen (Leber, Gallenblase, Gallenwege, Pankreas) sowie die innere Schicht des Atmungssystems (Tracheobronchialsystem), der Allantois und das innere Blatt der Kloaken- und Rachenmembran.
Abb. 26 Querschnitt durch die Keimscheibe in Höhe der Primitivrinne. Die Ektodermzellen wandern nach innen.
VII 1.2. Primitivknoten als Organisationszentrum Der Hensen-Knoten, der Ort der Kopffortsatzeinstülpung verschiebt sich kontinuier-lich nach hinten, so dass sich die Primitivrinne verkürzt und der Kopffortsatz oder Chordakanal, also die spätere Chorda dorsalis, sich verlängert.
Der zurückwan-dernde Primitiv-knoten ist ein Or-
ganisations-zentrum, das in den Keimblättern die Ausbildung der Achsenorga-ne des Embryos induziert. Durch Invagination in die Primitivgrube wird das Zellma-
Abb. 27

Embryologie aus Sicht der Craniosacralen Osteopathie 44/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
terial im Primitivstreifen als Chorda vor dem Knoten niedergelegt. Sich zurückzie-hend induziert es lateral die Organisation des Mesoderms in Somiten und im dar-über liegenden Ektoderm die Neuralplatte und anschliessend zwei aufeinander zu-wachsende Neuralfalten, die sich zu einem Rohr, dem Neuralrohr zusammenschlies-sen. Der Neuralrohrschluss beginnt kranial und folgt wie die Somitenbildung der Wanderung des Primitivknotens nach kaudal.
VII 1.3. Die ventrale Mittellinie oder Chorda dorsalis Sie wird auch Notochord (Urwirbelsäule) oder pirmäre primitive Mittellinie genannt. Sie ist die erste in der Mitte verlaufende Struktur.
2. Medianschnitt. Vor dem Primitivknoten ver-schmilzt der Boden des Axi-alkanals mit dem Entoderm und löst sich auf. C) Das Material des Chordafortsat-zes wird in das Entoderm eingeschaltet und bildet an dieser Stelle das Dach des Dottersackes. Die Primitiv-grube öffnet sich vorüber-gehend über den Axialkanal in den Dottersack hinein (Pfeil). Der Canalis neutralis verbindet vorübergehend den späteren Zentralkanal mit dem späteren Darmrohr. Kranial schliesst sich an den Chordafortsatz ein verdick-ter Entoderm-bezirk an, der als Praechor-dalplatte be-zeichnet wird. Auch im Be-reich der späteren Kloa-kenmembran bleiben Ento-derm und Ektoderm in direk-tem Kontakt. 3. Querschnitte durch die Keimscheibe. a) Im Bereich des Primitiv-streifens gehen Ektoderm und Mesoderm ineinander über. Die flache Entoderm-schicht hat sich etwas abge-löst. b) Die Primitivgrube im Primitivknoten setzt sich
nach vorn in den Axialkanal fort. c) Der Axialkanal verläuft als kleiner epithelialer
Abb. 28
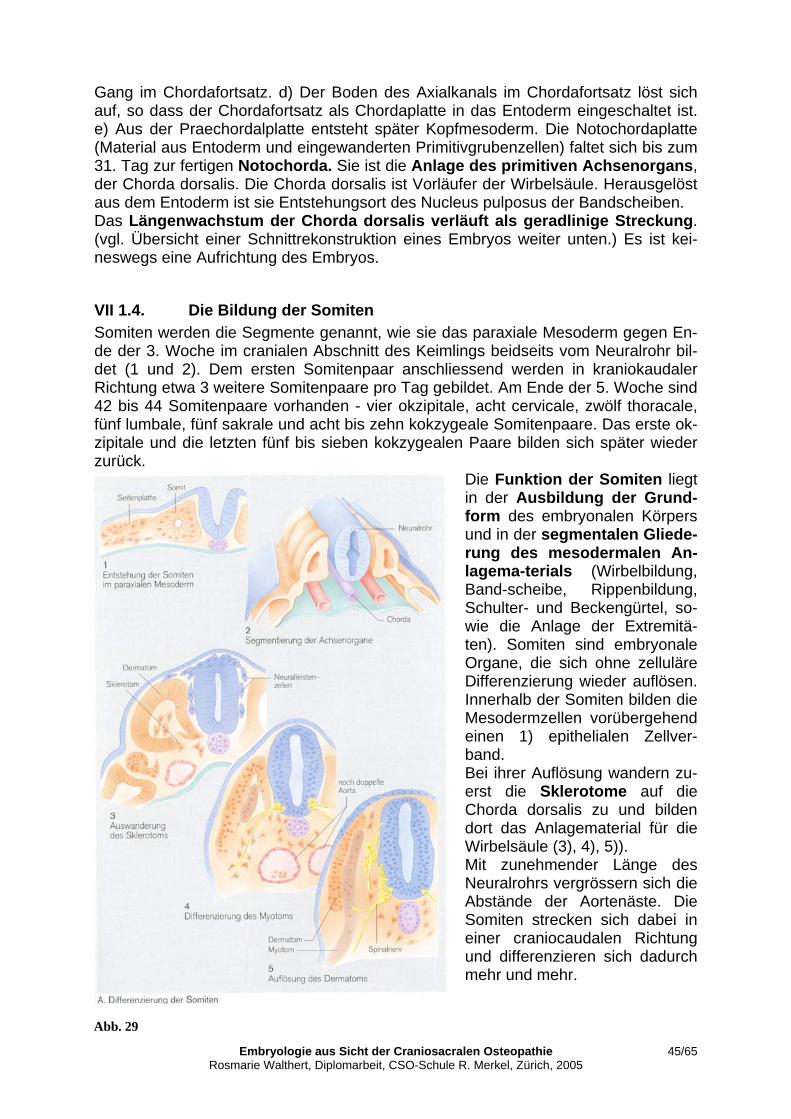
Embryologie aus Sicht der Craniosacralen Osteopathie 45/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Gang im Chordafortsatz. d) Der Boden des Axialkanals im Chordafortsatz löst sich auf, so dass der Chordafortsatz als Chordaplatte in das Entoderm eingeschaltet ist. e) Aus der Praechordalplatte entsteht später Kopfmesoderm. Die Notochordaplatte (Material aus Entoderm und eingewanderten Primitivgrubenzellen) faltet sich bis zum 31. Tag zur fertigen Notochorda. Sie ist die Anlage des primitiven Achsenorgans, der Chorda dorsalis. Die Chorda dorsalis ist Vorläufer der Wirbelsäule. Herausgelöst aus dem Entoderm ist sie Entstehungsort des Nucleus pulposus der Bandscheiben. Das Längenwachstum der Chorda dorsalis verläuft als geradlinige Streckung. (vgl. Übersicht einer Schnittrekonstruktion eines Embryos weiter unten.) Es ist kei-neswegs eine Aufrichtung des Embryos.
VII 1.4. Die Bildung der Somiten Somiten werden die Segmente genannt, wie sie das paraxiale Mesoderm gegen En-de der 3. Woche im cranialen Abschnitt des Keimlings beidseits vom Neuralrohr bil-det (1 und 2). Dem ersten Somitenpaar anschliessend werden in kraniokaudaler Richtung etwa 3 weitere Somitenpaare pro Tag gebildet. Am Ende der 5. Woche sind 42 bis 44 Somitenpaare vorhanden - vier okzipitale, acht cervicale, zwölf thoracale, fünf lumbale, fünf sakrale und acht bis zehn kokzygeale Somitenpaare. Das erste ok-zipitale und die letzten fünf bis sieben kokzygealen Paare bilden sich später wieder zurück.
Die Funktion der Somiten liegt in der Ausbildung der Grund-form des embryonalen Körpers und in der segmentalen Gliede-rung des mesodermalen An-lagema-terials (Wirbelbildung, Band-scheibe, Rippenbildung, Schulter- und Beckengürtel, so-wie die Anlage der Extremitä-ten). Somiten sind embryonale Organe, die sich ohne zelluläre Differenzierung wieder auflösen. Innerhalb der Somiten bilden die Mesodermzellen vorübergehend einen 1) epithelialen Zellver-band. Bei ihrer Auflösung wandern zu-erst die Sklerotome auf die Chorda dorsalis zu und bilden dort das Anlagematerial für die Wirbelsäule (3), 4), 5)). Mit zunehmender Länge des Neuralrohrs vergrössern sich die Abstände der Aortenäste. Die Somiten strecken sich dabei in einer craniocaudalen Richtung und differenzieren sich dadurch mehr und mehr.
Abb. 29
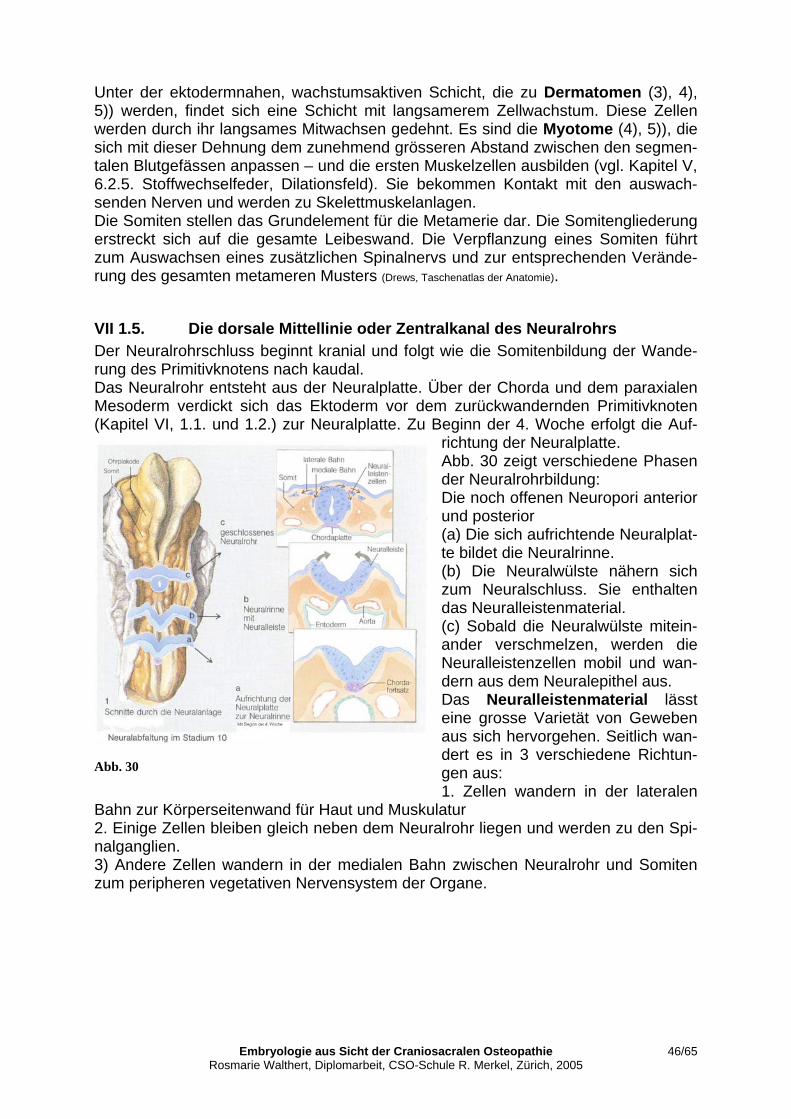
Embryologie aus Sicht der Craniosacralen Osteopathie 46/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Unter der ektodermnahen, wachstumsaktiven Schicht, die zu Dermatomen (3), 4), 5)) werden, findet sich eine Schicht mit langsamerem Zellwachstum. Diese Zellen werden durch ihr langsames Mitwachsen gedehnt. Es sind die Myotome (4), 5)), die sich mit dieser Dehnung dem zunehmend grösseren Abstand zwischen den segmen-talen Blutgefässen anpassen – und die ersten Muskelzellen ausbilden (vgl. Kapitel V, 6.2.5. Stoffwechselfeder, Dilationsfeld). Sie bekommen Kontakt mit den auswach-senden Nerven und werden zu Skelettmuskelanlagen. Die Somiten stellen das Grundelement für die Metamerie dar. Die Somitengliederung erstreckt sich auf die gesamte Leibeswand. Die Verpflanzung eines Somiten führt zum Auswachsen eines zusätzlichen Spinalnervs und zur entsprechenden Verände-rung des gesamten metameren Musters (Drews, Taschenatlas der Anatomie).
VII 1.5. Die dorsale Mittellinie oder Zentralkanal des Neuralrohrs Der Neuralrohrschluss beginnt kranial und folgt wie die Somitenbildung der Wande-rung des Primitivknotens nach kaudal. Das Neuralrohr entsteht aus der Neuralplatte. Über der Chorda und dem paraxialen Mesoderm verdickt sich das Ektoderm vor dem zurückwandernden Primitivknoten (Kapitel VI, 1.1. und 1.2.) zur Neuralplatte. Zu Beginn der 4. Woche erfolgt die Auf-
richtung der Neuralplatte. Abb. 30 zeigt verschiedene Phasen der Neuralrohrbildung: Die noch offenen Neuropori anterior und posterior (a) Die sich aufrichtende Neuralplat-te bildet die Neuralrinne. (b) Die Neuralwülste nähern sich zum Neuralschluss. Sie enthalten das Neuralleistenmaterial. (c) Sobald die Neuralwülste mitein-ander verschmelzen, werden die Neuralleistenzellen mobil und wan-dern aus dem Neuralepithel aus. Das Neuralleistenmaterial lässt eine grosse Varietät von Geweben aus sich hervorgehen. Seitlich wan-dert es in 3 verschiedene Richtun-gen aus: 1. Zellen wandern in der lateralen
Bahn zur Körperseitenwand für Haut und Muskulatur 2. Einige Zellen bleiben gleich neben dem Neuralrohr liegen und werden zu den Spi-nalganglien. 3) Andere Zellen wandern in der medialen Bahn zwischen Neuralrohr und Somiten zum peripheren vegetativen Nervensystem der Organe.
Abb. 30

Embryologie aus Sicht der Craniosacralen Osteopathie 47/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 1.6. Abschliessende Gedanken zum energetischen Aspekt Zusammenfassend möchte ich für das Organisationszentrum Primitivknoten den zeit-lichen Rahmen hervorheben. Innerhalb von wenigen Tagen, nämlich vom 16. bis zum 19. Tag induzieren Primitivstreifen und der in der Längsachse der Keimscheibe zurückwandernde Chordafortsatz mit Axialkanal und dem nachwandernden Primitiv-knoten gewaltige Schritte in der Entwicklung. Innerhalb von nur 3 Tagen hat das zu-rückwandernde Organisationszentrum Primitivknoten die Entwicklung von Chorda dorsalis, Somiten und Neuralrohr eingeleitet. Hervorheben möchte ich nochmals den ebenfalls von cranial nach caudal mit dem Chordafortsatz mitwandernden Axialfort-satz, der vorübergehend die beiden Flüssigkeitsräume Amnion (dorsal der Keim-scheibe liegend) und Dottersack/Darmbläschen (ventral der Keimscheibe liegend) miteinander verbindet. Amnion- und Dottersackflüssigkeiten stehen vorübergehend (ca. 1 Tag lang) im Austausch miteinander.
Abb. 30 a Von der zweidimensionalen Scheibe zum dreidimensionalen Körper

Embryologie aus Sicht der Craniosacralen Osteopathie 48/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Damit werden so viele grundsätzliche Entwicklungsbewegungen induziert, dass die-ser Prozess mir als einer der bedeutendsten Entwicklungsschritte in der embryonalen Phase überhaupt erscheint. Auf den Impuls der einwandernden Zellen beginnen, wie eben gesagt, gleich auf drei Ebenen Wachstumsbewegungen (Entwicklung der Chorda dorsalis, der Somiten und des Neuralrohrs). Gleichzeitig wächst eine neue Gewebeschicht (das Mesoderm) zwischen die beiden schon bestehenden Schichten Ektoderm und Entoderm. In der Ausdehnung der drei erwähnten Anlagen (Chorda, Somiten und Neuralrohr) in der ganzen Rumpflänge des Embryos bis in die Kopfre-gion liegt ein gewaltiges Potenzial. Entlang ihres gesamten Verlaufs organisieren sich die weitere Entwicklung der Organe und sogar die Gestaltwerdung des Embry-os. Der Lauf des Chordafortsatzes zeichnet das Muster der Mittellinie für das wer-dende Zentralnervensystem, die Grundgestalt des menschlichen Körpers, für Skelett-Kopf- und Gesichtsbildung. Und durch die Verschmelzung des Axialkanals mit dem Entoderm hat es Anteil am späteren Darmrohr. Chordafortsatz und Axialkanal wirken tief in die Keimscheibe ein. Wie ein wei-cher Malstift prägen sie ein Zeichnungsmuster, in dessen Verlauf die Entwick-lung zur Dreidimensionalität angelegt wird. Damit beginnt die räumliche Entwick-lung des Embryos. Er bekommt seinen Körper. Vielleicht kann der Lebensatem - „Breath of Life“, Sutherland - in Zusammenhang mit diesem embryonalen Entwicklungsschritt gesehen werden? Das biodynamische Modell der Craniosacralen Osteopathie (Begründer sind un-abhängig voneinander Jealous, James, S. (DO) und Sills, Franklyn) Jealous hebt besonders die ventrale Mittellinie, die Chorda dorsalis hervor. Sills unterscheidet eine ventrale von einer posterioren Mittellinie. Er ordnet ihnen un-terschiedliche Funktionen zu, die ein Leben lang zusammenwirken. Die primäre oder ventrale Mittellinie ist eine Orientierungsachse für den Bereich der Zellen und Gewebe. Zeitlebens sorgt die Mittellinie dafür, dass die Organisation der menschlichen Gestalt aufrechterhalten bleibt. Der Chordafortsatz reicht bis zur Regi-on der Hypophyse (sella thurcica ossis sphenoidalis). Ihr energetisches Prinzip reicht aber bis ins Os ethmoidale. Ihre Natur entspricht dem Luftigen. Eine weitere Mittellinie liegt posterior. Es ist die Orientierungsachse für die Flüssig-keitsebene. Sie verläuft im liquorgefüllten Zentralkanal bis in den 3. Ventrikel. Ihre Natur entspricht dem Flüssigen. Die beiden Mittellinien sind miteinander verbunden und halten gemeinsam die Orga-nisationsfunktion von Flüssigkeit, Zellen und Gewebestruktur aufrecht. Die Begriffe Primitivrinne, Chorda dorsalis, Axialfortsatz, und Neuralrohr aus der Embryologie veranlassen mich, etwas auszuholen. Fernöstliche Medizintraditionen Franklyn Sills hat die Arbeit von Dr. Stone im Buch „The Polarity Process“ beschrie-ben. Er war selber Instruktor für Polarity-Therapie bevor er sich der Craniosacralen Biodynamik zugewendet hat. Dr. Randolph Stone, Doktor der Osteopathie, der Chiropraktik und Naturheilkundiger und Begründer der Polarity Therapie, hatte sich viel mit fernöstlichen Medizintraditio-nen und Kulturen befasst. In der indischen Tradition, der ayurvedischen Medizin, ist er mit dem System der Nadis in Berührung gekommen.

Embryologie aus Sicht der Craniosacralen Osteopathie 49/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Nadi bedeutet Röhre oder Ader. Die Nadis entsprechen in etwa den Nerven. Sie ha-ben jedoch nicht nur physische, sondern auch feinstoffliche Bedeutung. Gehirn und Wirbelsäule sind für einen Yogi sehr wichtig. Ein feinstofflicher Strang führt angefangen beim After in der ganzen Länge des Rückenmarkkanals bis hoch zum Schädeldach. Der Sanskritname dieses Stranges ist susumna. In seinem Ver-lauf sind die 7 grössten Chakras (Energieräder) eingeordnet. Auf beiden Seiten die-ses zentralen Stranges führt je ein weiterer „Nervenstrang“ (Nadi), der in Sanskrit ida bzw. pingala genannt wird. Diese beiden Nadis reichen von den beiden Nasenlö-chern via Schädelbasis bis zum Steissbein. Ida und pingala sind Begriffe aus der Yogaphilosophie, die in etwa dem parasympathischen und sympathischen Nerven-system entsprechen. Es gibt insgesamt rund 72000 Nadis im menschlichen Körper. Yogis beschäftigen sich in Meditation und Atemführung viel mit den nadis und kon-zentrieren sich auf sie. Besonders wichtig ist susumna, denn zuunterst „schläft“ Kun-dalini sakti, die schöpferische Energie, die göttliche Natur. Das Ziel des Yoga ist es, Sakti zu erwecken. Sakti soll durch die Wirbelsäule aufsteigen - bis zum Sitz von Si-va. Siva ist reines Bewusstsein und hat seinen Sitz im Schädeldach. Sakti soll zu Si-va aufsteigen und sich mit ihm vereinigen. In dieser Vereinigung von Natur und Be-wusstsein wird der Mensch frei von Sorgen, Verstrickungen und Problemen im ge-sundheitlichen, sozialen und psychischen Bereich. Den Zustand von Freiheit erreicht er durch Reinheit von Körper und Geist, durch rich-tiges Üben und richtige Atmung und durch die Gnade Gottes. Mit Atmung ist auch die innere Atmung, die Zellatmung und die feinstoffliche Ebene der Atmung, die Lebens-energie gemeint ist (Yogi Dhirananda (S. K. Ghosh), Yogamrita, die Essenz des Yoga). Es gibt weitere alte Konzepte von menschlichen Energiefeldern, die sich an der Mit-telachse orientieren. So ist das Meridiansystem aus der traditionell chinesischen Me-dizin ein Beispiel, das ich hier nicht näher bespreche. Das tibetische ist dem indi-schen System sehr ähnlich. Dr. W. G. Sutherland hatte, wie schon erwähnt im Zusammenhang mit dem primären Atemmechanismus vom „Breath of life“ gesprochen. Ich gehe davon aus, dass er mit diesem Begriff Bewusstsein oder Energie gemeint haben könnte, wie sie oben mit den Mittellinien oder den Energiekanälen aus östlichen Traditionen beschrieben sind. Der Gedanke von Energiefluss und Lebenskraft ist dem Westen nicht fremd. Auch wir kennen in unserer christlichen Kultur Begriffe wie Seele (Inneres des menschli-chen Individuums) oder „Heiliger Geist“ (allgemein göttliche oder universelle Ebene). In der modernen, industrialisierten Gesellschaft sprechen wir aber eher in einem in-tellektualisierenden, analysierenden Sinn davon und geben der Ebene der erfahrba-ren Empfindung keine ernsthafte Bedeutung. Der „Heilige Geist“ kann für den Gläu-bigen Energiefluss und Lebenskraft sein. Wir sprechen von Lebenskraft und Energiefluss, ohne nach Instrumenten zu suchen, wie wir sie verinnerlichen und in unseren Körpern empfindend erlebbar machen könnten. Von dieser Warte aus wird Gesundheit zum Konsumgut, das von aussen abhängt. Man kann es an Fachpersonen in Medizin oder im Wellnessbereich delegie-ren. So suchen wir Heilwerden im Aussen. Der Gedanke, Heilwerden wirklich im In-neren entstehen zu lassen, bleibt uns fremd.

Embryologie aus Sicht der Craniosacralen Osteopathie 50/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Überblick einer Schnittrekonstruktion der inneren Organe eines 4.2 mm grossen Embryos im Stadium 13
Nervensystem: Die Anlage des Gehirns gliedert sich in das Prosenzephalon mit den Augenbläschen (1), das Mesenzephalon (2) und das Rhombenzephalon (3). Die Schlundbogennerven N. trigeminus (4), N. facialis (5), N. glossopharyngeus, (6) Ohr-bläschen, (7) und N. vagus (8) wachsen aus, ebenso wie die Spinalnerven, Spinal-ganglienleiste (9) . Herz und Gefässe: von der Herzschleife ist die Myokardoberfläche dargestellt. Der Vorhofabschnitt (a), die Kammer (b) und der Bulbus (c) sind durch Furchen gegen-einander abgegrenzt. Vom Venensystem sind sichtbar die V. cardinalis sup. (10), der gemeinsame Stamm der Kardinalvenen (11), die V. cardinalis inf. (12), die V. umbili-calis und der Plexus der Dottervenen (22). Arteriensystem: Aorta mit Intersegmental-arterien (13) und A. umbilicalis (18). Darmrohr: Unterhalb der Schlundtaschen sind die Lungenknospe (20) und unter dem Herzen das Netzwerk der Leberzellbälkchen sichtbar (21), die in die Gefäss-bahn der Dotter- und Nabelvenen (22) hineinsprossen. Chorda dorsalis: (15) liegt ventral vom Neuralrohr

Embryologie aus Sicht der Craniosacralen Osteopathie 51/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 2. Gestaltende Bewegung Im Primären Atemmechanismus sind zwei verschiedene Bewegungsaspekte ange-legt. Submikroskopische Stoffwechselbewegungen Der eine Aspekt liegt in seiner späteren Funktion, der Förderung von submikroskopi-schen Stoffwechselbewegungen. Diese Funktion wird meiner Meinung nach über die lange Phase der Organogenese des Embryos aufgebaut. Ich erinnere an den Grundsatz der Kinetischen Entwicklungstheorie, der besagt, dass die submikroskopischen Elemente der Entwicklungsbewegung Stoffwechselbe-wegungen sind. Die Entwicklungsbewegung des Nervensystems als Wachstumsleistung scheint mir grundlegend zu sein für diese spätere vernetzte Aufgabe. Sie kann aber nur in Ab-hängigkeit zur Entwicklung der übrigen Organe gesehen werden. Physiologische Bewegungsrichtung oder inhärente Bewegung des PAM Der andere Aspekt ist die physiologische Bewegungsrichtung oder inhärente Bewe-gung, die der Primäre Atemmechanismus (PAM) später über die Anheftungsstellen der Reziproken Spannungsmembran auf die Schädelknochen ausüben wird (vgl. Ka-pitel IV). Erich Blechschmidt’s Kinetische Entwicklungstheorie gibt jeder Entwicklungsbewe-gung eine funktionelle Bedeutung: Die Grundfunktion der Organe in der embryonalen Phase ist die Gestaltungsfunktion. Das Nervensystem kann den Mechanismus des Primären Atems nicht alleine ausfüh-ren, sondern nur im Zusammenwirken mit dem umgebenden Flüssigkeitsraum inner-halb der bindegewebigen und knöchernen „Hüllen“. Die Bewegung wird über die Flüssigkeiten auf die cranialen/spinale Meningen und die knöchernen Strukturen des beweglichen Schädels und des Sakrums übertragen. Die Bewegung setzt sich im gesamten Körper fort. Über Austrittsstellen von Nerven und Gefässen durch Öffnun-gen im Schädelbereich und in den Foramina der Wirbelsäule bestehen Verbindungen zum Bindegewebe des ganzen Körpers. Tatsächlich bestehen bereits embryonal ausgeprägte, wachstumsfunktionelle Einflüsse zwischen ektodermaler Hirn-/Rückenmarkanlage und mesodermalem Gefässsystem. Das embryonale Rücken-mark entwickelt sich alles andere als eigenmächtig. Das Wachstum geschieht in en-gem funktionellem Zusammenhang mit dem Bindegewebe seiner Umgebung. Immer wieder hat E. Blechschmidt darauf hingewiesen, dass das Nervensystem (Ektoderm) schneller wächst als das Gefässsystem (Mesoderm) und dass das Gefässsysstem das Nervensystem bevorzugt versorgt. Dabei hat das Mesoderm mit seinem Gefäss-system nicht nur eine ernährende Funktion. Es dient dem werdenden Neuralrohr schon sehr früh auch als Halteapparat. Die entsprechende Entwicklungsgebärde für diese inhärente Bewegung kann in der Krümmungsphase des Embryos gesehen werden. Es ist die Zeit, in der der Embryo die Anlage des Nervensystems entwickelt, seine Körpergrundgestalt annimmt und al-le Organsysteme anlegt. Die Krümmungsphase übt die spätere Flexionsphase oder Inspirationsphase des Primären Atemmechanismus vor.

Embryologie aus Sicht der Craniosacralen Osteopathie 52/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Für die Betrachtung der Bewegungsebene möchte ich mit der Bildung von Herz und Kreislauf beginnen. Die direkte entwicklungskinetische Beziehung des Gehirns zum Herzen ist für meine Überlegungen von grosser Bedeutung. Das Nervensystem wird vom Gefässsystem in seinem Wachstum gehemmt.
VII 2.1. Blutbildung und Herzkreislauf Die Entwicklung des Herzens, formuliert Blechschmidt, steht im Dienste der Blutver-sorgung des jungen Gehirns. Mit zunehmender Vergrösserung des Herzens im Dienste der Blutversorgung des Gehirns entwickelt sich noch im 1. Monat als Hilfe des Herzens, gleichsam als Vorfilter, die Leber. Die Differenzierung der Blutgefässe beginnt etwa Mitte der 3. Woche im Mesoderm vor der Praechordalplatte. Dort und auch im Bereich des Haftstils werden sie als Blut-inseln sichtbar. Die Blutinseln bestehen aus Zellnestern, die sich aussen zu Endo-thelzellen (Angioblasten) und innen zu den Blutzellen differenzieren. Durch Aus-sprossen der Endothelzellen verbinden sich die Blutinseln miteinander. Eine so klei-ne embryonale Anlage ist rasch mit Flüssigkeit durchströmt.
Abb. 31
Entwicklungsphasen des Herz-Kreislaufsystems (nach Rohen): 13.-15. Tag Blutinseln entstehen auf dem Dottersack, im Haftstil und im Chorion 17. Tag Erste Gefässe bilden sich aus 20. Tag Primitive Herzschläuche entstehen 21. Tag Die Herzanlage bekommt Verbindung mit den Gefässen 22. Tag Das Herz beginnt zu schlagen

Embryologie aus Sicht der Craniosacralen Osteopathie 53/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Die Herzanlage entsteht am Kopfende in einem Spaltraum zwischen Ekto- und En-toderm (19. -21. Tag). In diese Perikardhöhle lagern sich mesenchymale Gefässin-seln zusammen und bilden so den Endokardschlauch. Die frühe Herzanlage (spätere Ventrikelregion) befindet sich eigentlich ausserhalb der Keimscheibe. Perikard und Endokardschlauch bewegen sich aber zunehmend nach caudal. Mit der Abfaltung des Neuralrohres im Kopfgebiet verlagert sich die Herzanlage unter die Gehirnanla-ge. Die Herzanlage macht insgesamt eine beträchtliche Einwärts- und Abwärtswan-derung von aussen nach innen, also vom Umkreis der Keimscheibe zur Mitte des Embryos. Das bald schon schlagende Herz ist zuerst Brücke zwischen den organi-schen Funktionsprozessen an der Peripherie (Chorion, Plazenta) und dem sich bil-denden Embryonalkörper. Das Gefässsytem aus Dottersack und Chorionzotten (spä-tere Plazenta) setzt sich in den embryonalen Körper fort.
VII 2.2. Die Gestaltungsfunktion des Gefässsystems Am seitlichen Rand der Embryonalanlage, nahe dem Übergang des Ektoderms in das Amnion, entstehen im Mesoderm die ersten Venen in einer Kanalisierungszone. Die Venen führen darin Blut aus den Haftstielgefässen zum Herzen. Von dort gelangt das sauerstoffreiche Blut vor allem zur Gehirnanlage. Aus dem Kopfbereich fliesst das Blut seitlich am Neuralrohr entlang wieder zum Haftstiel zurück. Dieser Weg ist entwicklungsdynamisch vorbereitet. Denn sobald sich das Neuralrohr schliesst, ge-winnt das Binnengewebe an seinen Flanken Raum. Diese Zone bleibt jedoch nicht leer, sondern wird von strömender Interzellularflüssigkeit erfüllt. Es entsteht die paa-rige Aortenanlage (vgl. Übersicht Schnittrekonstruktion weiter oben). Die genannte Entwicklung ist eine Folge davon, dass schon vor der Bildung des eigentlichen Blut-kreislaufs nahrungsreiche Interzellularsubstanz in einem Konzentrationsgefälle in Richtung Gehirnanlage fliesst. Die Gefässbildungen sind also biodynamisch folge-richtige Differenzierungen. Die Vorform für die Gefässbildungen sind feine kanalförmige Interzellularsub-stanzstrassen im Binnengewebe. Diese Kanalisierungen finden sich regelmässig dort, wo räumlich Gelegenheit und raum-zeitlich (physikalisch) unmittelbare Veran-lassung für ihre Bildung besteht, d. h. wo ein Stoffwechselgefälle wirksam ist. Auch die zunächst kapilläre Aortenanlage richtet sich nach der prioritären Blutvertei-lung im Embryo. Ihre Verzweigungen gelten nämlich ebenfalls dem hauptsächlichen Nahrungsverbraucher, dem Neuralrohr. Hier findet man in regelmässigen Abständen nacheinander, also metamer, die ersten Rückenäste der Aorten entstehen. Sie unter-teilen das Mesoderm in einzelne Segmente. Durch diese Unterteilung entstehen die sogenannten Somiten oder Körperwandorgane. (Vgl. Kap. VII 1.4.) Mit dem Wachstum der segmentalen Aortenäste beidseits des Neuralrohrs und der Unterteilung des Mesoderms in Somiten entstehen Einkerbungen des benachbarten Ektoderms. Die Einkerbungen sind eine Folge davon, dass am Ende der Gefässe soviel Nahrung aus ihnen entnommen wird, dass sie selbst zusammen mit ihrem Gewebsbett im Wachstum zurückbleiben. Blechschmidt hat regional vergleichende Untersuchungen gemacht, die bestätigen, dass alle grösseren Gefässe ähnliche Gestaltungsfunktionen haben. Sie wachsen al-le relativ langsamer als die Zellverbände ihrer Versorgungsgebiete. Der Segmentation des Gefässsystems folgt die Segmentation des Nervensystems. Auch sie ist kinetisch gesehen eine Folge der Somitenbildung (vgl. Kapitel VII, 1.4.).

Embryologie aus Sicht der Craniosacralen Osteopathie 54/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Durch das Flächenwachstum der Somiten ist die Bildung der Spinalnerven vorberei-tet. Sobald sich ein Somit auf das Neuralrohr „stützt“ und sich gleichzeitig mit seinem dorsalen Dermatomanteil nach aussen vorwölbt, entsteht eine Schröpffunktion. Folge davon ist das Auswachsen der ventralen Wurzel senkrecht zur Oberfläche des Neu-ralrohrs. Ungefähr gleichzeitig entstehen die Spinalganglien. Das Neuralrohr wächst dorsal besonders schnell, während sein Bett seitlich und ventral kurz bleibt (Anlage der Dura). Durch das dorsal expansive Wachstum des Neuralrohrs werden die den Somiten anliegenden Teile des Neuralrohrs in Form der Spinalganglien abgestreift. So ist die metamere Bildung von dorsaler und ventraler Wurzel der Spinalnerven in entwicklungsdynamischer Abhängigkeit zu den „Ursegmenten“ Somiten entstanden.
Wie in Kapitel „Erste Leistungen des Ner-vensystems“ beschrieben, wird die späte-re efferente Leistung eines Neuriten mor-phologisch bereits hier durch karyofugal abfliessendes Zytoplasma aus dem Zell-kern abgeleitet. Blechschmidt nimmt an, dass für das dafür notwendige Strö-mungsgefälle die Wachstumsdehnung der Muskelanlage im Bereich der Somiten massgebend ist. Abb. 32 zeigt die Entste-hung einer dorsalen und ventralen Wurzel im Wachstumsfluss (Fluxion). Die ver-schiedenen Ernährungsbedingungen von afferenten und efferenten Fortsätzen zeichnen die Richtungen der späteren Er-regungsleitung. 1) langsames Wachstum, daher Wider-stand 2) Neurit. Als Vorbereitung der späteren afferenten Leistung der hinteren Wurzel hat Blechschmidt eine karyopetale Fluxi-onsrichtung beschrieben (geschwänzter Pfeil). 3) Neuroganglion,
4) Dendrit, 5) Neurit, 6) Ausscheidung von Liquor cerebrospinalis, 7) Ausscheidung von Liquor arachnoidalis.
Abb. 32

Embryologie aus Sicht der Craniosacralen Osteopathie 55/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Abb. 33 zeigt die Kontaktzonen zwischen Nerven und Blutgefässen. Durch den As-census des Neuralrohrs liegen die Nerven jeweils am Unterrand der Gefässe. Wäh-rend das Neuralrohr (punktiert) nach oben weiter wächst, streift es die Spinalnerven gleichermassen am Gefässsystem (Vene grau und Arterie schwarz) ab.
Der Wachstumswiderstand der Aortenanla-ge führt dazu, dass das Neuralrohr sich an seinem frei beweglichen Ende (im Kopf-gebiet) über den Herzwulst krümmt. Mit dieser Krümmung entstehen Beugefalten - das frühembryonale Gesicht. Es verbreitert sich über dem Herzwulst quer. Die Beu-gefalten bilden quere Bögen, die das Kopfdarmlumen ventral umgreifen (Visceralbö-gen oder Schlundbögen).
Abb. 35
Mit der zunehmenden Krümmung des Embryo in der Kopfregion werden die Visce-ralbögen mehr und mehr verbreitert und das Gewebe in ihrem Inneren dadurch ge-strafft und zirkulär zum Kopfdarm ausgerichtet (Abb. 34/35). Das so ausgerichtete Gewebe wird zur Leitstruktur für grosse Gefässe, die zwischen der kurzen ventralen Aorta und den längeren dorsalen Aorten Kurzschlüsse herstellen. Diese Visceralbogenarterien wölben jeweils die Körperwand nach aussen und nach innen vor. Die Wand wird dadurch bogenförmig dick, bleibt aber zwischen den Bögen dünn. Im Schnitt entsteht ein Bild wie Abb. 34. Die dünnen Zonen erscheinen aussen als Kerben, innen dagegen als Taschen (Schlundtaschen).
Abb. 33
Abb. 34

Embryologie aus Sicht der Craniosacralen Osteopathie 56/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 2.3. Die Ausgestaltung der Körpergrundgestalt Bevor ich die Abfaltungen bespreche, wie sie für die Annahme der Körpergrundge-stalt (nach Seidel) wichtig sind, möchte ich die Begriffe Flexion/Extension und Inhala-tion/Exhalation hervorheben. Einerseits drücken Flexion/Extension Bewegungsrichtungen des Körpers aus. Dazu gehören auch die Krümmung von Nacken und Wirbelsäule nach vorne oder hinten. Die kraniokaudale Krümmungsphase des Embryos wird im Folgenden besondere Bedeutung bekommen. Andererseits gelten dieselben Begriffe aber auch für die Bewegungsphasen des Pri-mären Atemmechanismus. Für das Konzept des Primären Atemmechanismus gelten für Flexion/Extension ausserdem synonym die Begriffe Inhalation/Exhalation. Die unpaaren Knochen machen in der Flexionsphase des PAM eine Bewegung ent-lang einer Mittellinie um eine transversale Achse, in der Extensionsphase im umge-kehrten Sinn (Abb. 3 Kap. IV 6). Die paaren Knochen machen in der Flexions- oder Inspirationsphase eine Aussenrotation von der Mittellinie weg, während der Extensi-ons- oder Exhalationsphase kehren sie zurück, machen also eine Innenrotation zur Mittellinie hin. Während der Entstehung der Körpergrundgestalt machen die Gewebeschichten der Keimscheibe zwei verschiedene Bewegungen: Zuerst kommt es zu einer lateralen Abfaltung, der unmittelbar eine kraniokaudale Krümmung folgt.

Embryologie aus Sicht der Craniosacralen Osteopathie 57/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 2.3.1. Die laterale Abfaltung Mit einer massiven Entwicklung von Körpersubstanz durch das intraembryonale Me-senchym, Bildung der Somiten und Faltung des Neuralrohres hebt sich der Keim von der Unterlage ab und wölbt sich zunehmend in die Amnionhöhle hinein vor, die sich inzwischen ebenfalls stark vergrössert hat. Die Keimscheibe gewinnt an Tiefe, wird räumlich.
Abb. 36
a) Die Somiten beginnen sich aufzulösen und Mesenchym zu bilden (dünne Pfei-
le). Die Keimscheibe, bei der sich die Neuralrohrbildung vollzieht (dicke Pfei-le), wird voluminöser. Übergang in eine räumliche Struktur (ca. 22. Tag)
b) Der Embryonalkörper beginnt sich vom Dottersack abzuheben (Pfeil) und ein intraembryonales Coelom (Leibeshöhle) zu entwickeln (23. Tag). Die Körper-höhlen geben Raum für die Entwicklung der inneren Organe. Dottersack und Entoderm beginnen mit der stufenweisen Faltung des Darmrohrs.
c) Der Embryonalkörepr ist bis auf den Nabelstrang völlig verselbständigt. Die „Körpergrundgestalt“ (nach Seidel) ist konstituiert (24. Tag). Vom Oberfläche-nektoderm umhüllt ruht er in seiner Fruchtblase
Das Amnion folgt dieser Körperbildung und formt vorne eine Kopffalte, an beiden Seiten entsprechende Seitenfalten und hinten eine Schwanzfalte aus, so dass nur ventral die Verbindung mit dem Dottersack und dem Trophoblasten erhalten bleibt.

Embryologie aus Sicht der Craniosacralen Osteopathie 58/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Hier hatte sich schon frühzeitig das sogenannt „extraembryonale“ Mesenchym zum Haftstil verdichtet. Jetzt bildet sich in diesem Bereich der vom Amnionepithel überzo-gene Nabelstrang. Als zweischichtige Keimscheibe bleibt der Embryo gerade. Auch mit der Abfaltung der Seitenwände und dem Vorwölben in die Amnionhöhle bleibt der Keimling mehr oder weniger gestreckt. Es wird vermutet, dass die beobachtete Hyperextension bei den Präparaten möglicherweise deshalb entstanden ist, weil die Spannungsverhält-nisse mit Eröffnen des Darmbläschens sich verändert haben.
VII 2.3.2. Die kraniokaudale Krümmung Die Abfaltungsbewegung geht am 23. Tag direkt in eine starke ventrale Krümmung der Embryonalachse über. Die treibende Kraft für diese Krümmung des Embryos ist das schnelle Wachstum der Neuralanlage. Das Längenwachstum ist dorsal einseitig, die Krümmung folgt sogleich. Blechschmidt weist nach, dass die Flügelplatte das Neuralrohrs viel rascher neue Zellen bildet als die Grundplatte. Durch die Volumen-zunahme dieser Zellen ergibt sich nicht nur eine exzentrische Wachstumsrichtung in die Länge, sondern auch eine fortschreitende Krümmung des Neuralrohrs und damit des Embryos. (Zeitschrift für Anatomie und Entwicklungsgeschichte, Bd. 120 S. 150–172 (1957)).
Die Kopffalte mit den Hirnbläschen über-wächst die Mundbucht und den Herzschlauch. Die Aortenanlage bin-det den Embryo in Beugehaltung zurück. Die Herzanlage wan-dert von ausserhalb der Keimscheibe vor dem Kopfende nach vorne unten in die Brusthöhle ein. Mit dem Eintreten des Herzens in die Brusthöhle wird der Hals schlanker und das frühe Breitgesicht sicht-bar. Vorher ist es völlig verdeckt vom Herz-wulst.
Was mag es bedeuten, dass die Herzanlage gleichsam von aussen, nämlich von ausserhalb der Keimscheibe sich unter die Gehirnanlage in den Hals schiebt und weiter in den Brustraum? Sie wird sogar zum ersten funktionstüchtigen Organ, wenn sie bereits am 22. Tag mit rhythmischen Schlägen für eine geordnete Strömungsrich-tung in den Gefässen sorgt. Es stellt sich die Frage, wieso das Herz als einzige An-lage von aussen kommt? Zu Beginn ist sie tatsächlich zu einem grossen Teil ausser-
Abb. 37

Embryologie aus Sicht der Craniosacralen Osteopathie 59/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
halb des eigentlichen Embryos im Einsatz. Aber es ist doch sehr speziell, dass gera-de das Herz von ausserhalb der Keimscheibe einwandert oder eingeholt wird, wo doch alle anderen Organe aus den eigentlichen Keimschichten selbst entstehen. Zu-dem steht das Einsetzen des rhythmischen Herzschlags in engem zeitlichem Rah-men zum Anfang der kraniokaudalen Krümmungsphase.
VII 2.3.3. Embryonales Üben des Primären Atemmechanismus (PAM) In der ganzen embryonalen Entwicklung gibt es nur diese eine einzige Geste des Einrollens. Mit Beginn der 4. Woche (1.5 mm) bis ungefähr zum 42. Tag bleibt der Embryo (10mm) in ausgeprägter Beugehaltung. Ich erlaube mir die Aussage: Der Embryo übt während seiner Krümmungsphase die Flexionsphase (Inspirationsphase) des primären Atemmechanismus vor, indem er mit dem wachsenden Neuralrohr die Herzanlage von ausserhalb der eigentlichen Keimscheibe umfasst, einholt, „einatmet“, verinnerlicht. Gleichzeitig atmet er die übri-gen inneren Organe ein, indem er sie, die Körperwände und Extremitätenanlagen mit Nervenzellen und –fortsätzen durchsetzt. Während dieser Geste der Ver-Beugung baut der Embryo schlussendlich mit seinem Nervensystem Kontakt zu jedem Zellsystem des ganzen Körpers auf.
Das Nervensystem bestreitet diese Ent-wicklungsarbeit bei der Ausbildung seiner Organgestalt samt Zentren und Bahnen nicht allein, sondern in gemeinsamer Zu-sammenarbeit mit den Organen in der näheren und weiteren Umgebung. In die-sem Wirkungszusam-menhang begrenzen die Gewebe, in die das Nervensystem eingeschlossen ist, durch Erzeugung ständig wachsender Widerstände all-mählich mehr und mehr den Fortgang der Entwicklung und determinieren so langsam die Ent-
Abb. 37 a 10 mm Mitte des 2. Monats

Embryologie aus Sicht der Craniosacralen Osteopathie 60/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
wicklungsmöglichkeiten, das Potenzial des ganzen Embryos gegenüber den Einwir-kungen (Reizen) denen der wachsende Organismus ausgesetzt ist (Blechschmidt). So finden die Wachstumsbewegungen des Neuralrohrs in Länge und widderhornarti-gen Rückwärtsbewegung (im Bereich der Grosshirnhemisphären) ein Ende am wei-testmöglichen „Bewegungsausschlag“ ihrer Entwicklungsmöglichkeit. Diese Phase entspricht der späteren Inspirationsphase des Respiratorischen Atemmechanismus. Dort am Umkehrpunkt, Ruhepunkt (Fulkrum?) erfährt die Entwicklungsbewegung des Nervensystems die Vorform der gegenläufigen Widerstandsbewegung – der Retrak-tion (Exspirationsphase) als einer Kraft von aussen – der Entwicklungsbewegung des mesodermalen Herz-/Gefässsystems. Eigentlich erfährt die Wachstumsbewegung nicht erst am Schluss der Beugungspha-se einen Widerstand, sondern fortlaufend. Das Neuralrohr generiert den Bewegungs-impuls für sein Längenwachstum, seine Expansionsphase. Die Wachstumsrichtung nimmt dabei die Richtung einer Beugung des Neuralrohrs und entspricht der PAM-Flexionsphase. Hingegen wirkt das Gefässsystem immer wieder wachstumshem-mend auf das Neuralrohr ein, bis es für die nötige Sauerstoffversorgung nachge-wachsen ist. Diese Wartephase auf Sauerstoff- und Nahrungsnachschub ist die Ex-tensionsphase der Hirnanlage und entspricht der Extensionsphase des PAM, wäh-rend welcher der zentrale Flüssigkeitsraum, die späteren Ventrikel „kontrahiert“ (Sut-herland) wird. Vielleicht ist es auch ein Nachgeben des Bewegungsimpulses im Sin-ne von Entspannen und Warten? (Anm. der Verfasserin). Der Embryo übt mit seinem Neuralrohr also eine „Entwicklungs-Gebärde“ (Walthert). Innerhalb dieser Gebärde alterniert der neuralrohreigene Bewegungsimpuls mit dem Hemmungswiderstand der Herz-/Gefässanlage. So wird die spätere alternierende in-härente Bewegung embryonal aufgebaut. Nach meiner Erfahrung ist die Flexionsbe-wegung stets deutlicher palpierbar als die schwächere Extensionsphase. Abgeschlossen wird die Entstehungsphase des Primären Atemmechanismus erst mit Ausgestaltung des Gehirns in der Fetalperiode. Für die vorliegende Arbeit beschrän-ke ich mich aber auf die beiden ersten Lebensmonate des Embryos.
VII 2.3.4. Embryonales Üben der sekundären Lungenatmung Dann, am 42. Tag beginnt der Embryo, sich mit der Ausbildung der knorpeligen Wir-belsäulenanlage aufzurichten. Dabei funktioniert das Knorpelskelett mit seinem Stemmkörperwachstum zunächst als relativ aktiver Teil des Bewegungsapparates gegenüber der Muskulatur. Die Muskulatur wird, wie überall im Körper, auch hier erst im Verlauf ihres Dehnungswachstums kontraktil. (Lokalisation der Muskulatur im Dila-tionsfeld, Stoffwechselfelder, vgl. Kap. V 6.2.5.) Das Zwerchfell ist lumbal bereits fest verankert. Diese Verankerung übt eine Halte-funktion auf die Zwerchfellkuppe und die Organe aus. Mit zunehmender Aufrichtung des Embryos deszendieren die Organe von Brust und Hals im Verhältnis zum Kopf.

Embryologie aus Sicht der Craniosacralen Osteopathie 61/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Abb. 38 Embryonen von links nach rechts: 4.2 mm; 10 mm; 29 mm
Mit dem Descensus des Zwerchfells erweitert sich der Thorax im Zusammenhang mit dem Wachstum der Rippen. Die ursprüngliche Leibeshöhle vergrössert sich hier. Es entsteht Raum für die Entwicklung der Lunge. Das Entoderm stülpt sich in dieses Sogfeld (vgl. Kap. V 6.2.5.) der Anlage des Respirationstraktes vor. Wie später beim Atmen die Luft eingesogen wird, so wird jetzt schon das Luftgefäss (Lungenflügel) in seinem Wachstum eingesaugt. Wachstum in Sogform bereitet die spätere Funktion vor. Während der Aufrichtungsphase des Körpers übt das embryonale Diaphragma mit seiner lumbalen Verankerung und dem Descensus in Bezug zum Kopf die spätere Einatmung in die Lungen vor. Es erfährt ein Dehnwachstum in die Kuppelform. Spä-ter wird es - sich kontrahierend - die Höhe der Kuppel abflachen, die unteren Rippen anheben und den Thorax in Einatemstellung führen. Weil die Lunge dieser Bewe-gung folgen muss (Vakuum zwischen den Pleurablättern), wird Luft einströmen.

Embryologie aus Sicht der Craniosacralen Osteopathie 62/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
VII 3. Parallelen und Unterschiede von Primärem und Sekundärem Atemme-chanismus. Beide Atemmechanismen kennen die Inspirationsphase gefolgt von einer Exspirati-onsphase. Primärer und sekundärer Atemmechanismus entwickeln sich aber unter-schiedlich. Das Nervensystem wächst aktiv in Inspirationsphase und reagiert passiv auf Wider-standskräfte/Zugkräfte (Exspirationsphase) von aussen, vom nachwachsenden Ge-fässsystem. Das Organ Lunge hingegen wird passiv in Inspiration ausgedehnt / eingesogen. Die Exspirationsphase übt es mit seinen Retraktionskräften aktiv aus.
VII 3.1. Wechselwirkungen der Gewebeschichten Ektoderm und Mesoderm sind die beiden Keimschichten, aus welchen die Strukturen mit Motorfunktion für den PAM hervorgehen. Ein kurzer Seitenblick auf die wechsel-seitig abhängigen Wachstumsbewegungen von Ektoderm und Mesoderm sei an die-ser Stelle erlaubt:
• Während der Krümmungsphase gilt: Das sich rasch entwickelnde ekto-dermale Neuralrohr wird in seinem Wachstum gehemmt durch die langsa-mer wachsende mesodermale Herz-/Gefässanlage.
• Die Aufrichtung des Embryos erfolgt nach dem Descensus des Herzens – erst mit ausreichender Stemmkörperfunktion des Knorpelskeletts - das Mesoderm übernimmt aktiv die Aufrichtung des embryonalen Körpers.
Für den Primären Atemmechanismus lässt sich daher sagen, dass der Embryo in der körperlichen Flexionsstellung alle Organanlagen einatmet - das Herz von aussen - begleitet von allen anderen, deren Anlage sich im Körperinneren bildet. Wenn er alle Organe eingeatmet hat, kann er während der körperlichen Aufrichtungsphase des Knorpelskeletts wieder ausatmen (Extension). Dies entspricht zeitlich der ersten Ü-bungsphase der sekundären Lungenatmung (Descensus des Diaphragmas).
VII 3.2. Relevanz für meine praktische Arbeit. Schon Sutherland hatte mit der mentalen Kontrolle und Lenkung des Kranialen Re-spiratorischen Mechanismus experimentiert (Hartmann, Das grosse Sutherland-Kompendium, Die Schä-
delsphäre III – 31). Selbst praktiziere ich Yoga. Yoga bietet wunderbare Instrumente über Atem-, Körper- und Meditationsübungen, um auf der energetischen Ebene mit sich selber zu arbei-ten (vgl. Kapitel VII, 1.6.). Entsprechend bevorzuge ich, andere Menschen auf dieser Ebene aus dem Fundus dieser Instrumente für die Praxis im Alltag zu unterrichten. Vermutlich liegt es daran, dass sich mir die energetische Ebene nicht so leicht er-schliesst. Wenn ich craniosacrale Osteopathie mit Menschen praktiziere, ist mir bis heute die Bewegungsebene stets deutlicher im Bewusstsein als die energetische. Damit meine ich die Ebene der Strukturen mit subtilsten Bewegungen und Körperbewegungen von kleinster bis grösserer Amplitude. Wenn es um den Bauchbereich, Diaphragmaarbeit und Arbeit in Brustraum, Hals und Kopfbereich geht, ist die Bewegungsebene in meiner Wahrnehmung vordergründig. Da sind für mich Ausdehnen und Zusammen-

Embryologie aus Sicht der Craniosacralen Osteopathie 63/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
ziehen der Flüssigkeitsebene, die longitudinale Ausrichtung der Meningen und natür-lich die physiologischen Bewegungen der Schädelknochen und die Bewegungen in Oberkörper und oberen Extremitäten in ihrer Normalität oder Funktionsstörung gut wahrnehmbar. Die energetische Qualität tritt im Bereich von Lumbalwirbelsäule und Beckenraum öf-ter in mein Bewusstsein. Ich kann mir bisher nicht erklären, warum es gerade diese Körperbereiche sind, die sich mir auf der energetischen Ebene leichter erschliessen und warum sie mir in anderen Körperbereichen weniger zugänglich ist. Die sekundäre Lungenatmungsbewegung ist überall deutlich. Und ich instruiere sie immer häufiger ganz gezielt. Fast regelmässig kann ich die Ebene des PAM leichter palpieren, wenn der/die KlientIn seine/ihre Aufmerksamkeit auf die sekundäre At-mung lenkt. Abschliessend muss ich eingestehen, dass dieser Arbeit viel praktische Tätigkeit fol-gen muss, um das hier zusammengetragene Wissen und die daraus hergeleiteten Theorien mehr in die Praxis zu integrieren.
Abb. 39

Embryologie aus Sicht der Craniosacralen Osteopathie 64/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Bildnachweis: Abb. 1 Hartmann, Das grosse Sutherland-Kompendium Abb. 2 Liem, Torsten, Kraniale Osteopathie Abb. 3 Liem, Torsten, Kraniale Osteopathie Abb. 4 Drews, Ulrich, Taschenatlas der Embryologie Abb. 5 Drews, Ulrich, Taschenatlas der Embryologie Abb. 6 Jaap van der Wal, Seminarunterlagen Abb. 7 - 13 Blechschmidt, Erich, Anatomie und Ontogenese des Menschen Abb. 14 Blechschmidt, Erich, Vom Ei zum Embryo Abb. 15 - 18 Blechschmidt, Erich, Anatomie und Ontogenese des Menschen Abb. 19 Drews, Ulrich, Taschenatlas der Embryologie Abb. 20 Blechschmidt, Erich, Die Entwicklung des menschlichen Nerven-
systems Abb. 21 Drews, Ulrich, Taschenatlas der Embryologie Abb. 22 Nach Sills Franklyn, Craniosacral Biodynamics Abb. 23, 24 Rohen, J. W., Funktionelle Embryologie Abb. 25 - 27 Rohen, J. W., Funktionelle Embryologie Abb. 28 – 30 Drews, Ulrich, Taschenatlas der Embryologie Abb. 30 a Blechschmidt, Erich, Der menschliche Embryo Abb. 31 Rohen, J. W., Morphologie des menschlichen Organismus Abb. 32 Blechschmidt, Erich, Die Entwicklung des menschlichen Nerven-
systems Abb. 33 – 35 Blechschmidt, Erich, Anatomie und Ontogenese des Menschen Abb. 36 Rohen, J. W., Funktionelle Embryologie Abb. 37 Sadler, T. W., Medizinische Embryologie Abb. 37 a Blechschmidt, Erich, Der menschliche Embryo Abb. 38 Blechschmidt, Erich, Der menschliche Embryo Abb. 39 Drews, Ulrich, Taschenatlas der Embryologie (nach Blechschmidt) Übersicht Schnittrekonstr.
Drews, Ulrich, Taschenatlas der Embryologie (nach Blechschmidt)

Embryologie aus Sicht der Craniosacralen Osteopathie 65/65 Rosmarie Walthert, Diplomarbeit, CSO-Schule R. Merkel, Zürich, 2005
Literaturangabe:
Blechschmidt, Erich; Anatomie und Ontogenese des Menschen; FE Medienverlag Kisslegg, 1978
Blechschmidt, Erich; Embryologie heute; Hippokrates, März 1974, S. 3 - 21 Blechschmidt, Erich, Die Frühentwicklung des Menschen, Hogrefe, 1966 Blechschmidt, Erich, Die Entwicklung des menschlichen Nervensystems, Hogrefe Blechschmidt, Erich; Die Stoffwechselfelder des menschlichen Eis; Zeitschrift Ge-
burtshilfe Perinatologie, März 1968, S. 143 - 155 Blechschmidt Erich; Der menschliche Embryo, 2. unveränderte Auflage, Schattau-
er, 1963 Blechschmidt Erich; Die praenatalen Organsysteme des Menschen; Hippokrates
Stuttgart, 1973 Blechschmidt Erich; Die vorgeburtlichen Entwicklungsstadien des Menschen; S.
Karger, 1961 Blechschmidt Erich; Wie beginnt das menschliche Leben; 7. erweiterte Auflage,
Christiana Verlag, 2002 Damasio, Antonio R., Descartes’ Irrtum, Fühlen, Denken und das menschliche Ge-
hirn, 2. Auflage, Ullstein - List Drews, Ulrich, Taschenatlas der Embryologie, Thieme, 1993 Hartmann, Christian, Das grosse Sutherland-Kompendium, Jolandos, 2004 Liem Torsten, Kraniosakrale Osteopathie, Hippokrates, 3. Auflage 2001 Liem Torsten, Praxis der Kraniosakralen Osteopathie, Hippokrates, 2000 Magoun Harald I., Osteopathie in der Schädelsphäre, Edition spirales, 3. Ausgabe
1976, deutsche Übersetzung 2000 Magoun Harald I., Osteopathy in the cranial field, The osteopathic cranial
association, Original Edition 1951, second printing 1997 Merkel Rudolf, Skript Nr. 1 – 6, Schule für Craniosacrale Osteopathie, Zürich O’Rahilly, Ronan / Müller, Fabiola; Embryologie und Teratologie des Menschen;
Huber, 1999 Paoletti Serge, Faszien, Urban und Fischer, 2001 Rohen Johannes W. / Lütjen-Drecoll Elke; Funktionelle Embryologie; 2. Auflage
Schattauer 2004 Rohen, Johannes W., Morphologie des menschlichen Organismus, 2.
überarbeitete Auflage, Verlag Freies Geistesleben, 2002 Sadler W. Thomas; Medizinische Embryologie, übersetzt von Ulrich Drews, 10.
korrigierte Auflage Thieme, 2003 Sills Franklyn, Craniosacral Biodynamics, vol. I, North Atlantic Books 2001 Sills Franklyn, Craniosacral Biodynamics, vol. II, North Atlantic Books 2004 Sills Franklyn, Energie-Arbeit, Goldmann Upledger, John E./ Vredevoogt, Jon D., Lehrbuch der CranioSacralen Therapie, 4.,
neu bearbeitete Auflage, Karl F. Haug Verlag, 2000 Van der Wal, Jaap, www.embryo.nl, [email protected]