2015 lter asm_poster
TRANSCRIPT
1995 2000 2005 2010
0.5
1.0
1.5
2.0
2.5
Year
BP
:PP ●
●
●
●
●●
●
●
●●
●
●
●
●●
●
●●
●
2000 2005 2010
1015
2025
3035
4045
Year
DO
C g
m
−2
●●
●
● ●
●
●
●●
●
●●
●
●
●●
●
●
2000 2005 2010
1015
2025
3035
4045
Year
DO
C g
m
−2
●
●
●
● ●
●
●
●
●●
●
●
●
● ●
●
●●
Lake FryxellW
est Lobe Bonney
2004 2006 2008 2010
0.00
0.05
0.10
0.15
Year
BP
:PP
●
●●
●
●
●
●
●
●
1995 2000 2005 2010
0.05
0.10
0.15
0.20
0.25
0.30
Year
BP
:PP
●
●
●
● ●
●●
●
● ● ●
●
●
●
● ●
●
●
●
●
2004 2006 2008 2010
5052
5456
5860
62
Year
DO
C g
m
−2
●
●
●
●
●
●
●
●
●
North Palm
er Region
02
46
Year
PP
g C
m
−2 d
−1
2004 2006 2008 2010
●
●
●
●
●●
● ●
●
●
●●
●
●
●
●
●
●
●
●
010
2030
4050
60
Year
PP
mg
C
m−2
d−1
1995 2000 2005 2010
●
●●
●
●● ●
●
●
●●
●●
● ●
●
●
●
●
●●
05
1015
2025
3035
Year
PP
mg
C
m−2
d−1
1995 2000 2005 2010
Assessing microbial ecosystem function across two polar extremes: The Palmer (PAL) and McMurdo Dry Valley (MCM) LTERs
*1Je� S. Bowman, 2Trista J. Vick-Majors, 3Rachael Morgan-Kiss, 4Christina Takacs-Vesbach, 1Hugh W. Ducklow, 2John C. Priscu
1Lamont-Doherty Earth Observatory, 2Montana State University, 3Miami University, 4University of New Mexico*[email protected] | www.polarmicrobes.org
2015 LTER ASM, Estes Park, CO
Our hypothesis is that polar desert lakes are di�erent from the coastal Antarctic marine ecosystem. As obvi-ous as the answer seems - one need only to consider the size of members of the top trophic levels to appre-ciate the huge ecological di�erences between these environments - making this comparison allows us to identify both common and unique ecological features of these sites associated with fundamental processes that might otherwise be overlooked. The 20+ year record of key ecosystem parameters at PAL and MCM provides a further opportunity to explore how these ecosystems respond to common events, such as the unusually warm austral summers in 2001-2002 and 2008-2009. To make our comparison we considered:
1. Records of bacterial production (BP), primary pro-duction (PP), and dissolved organic carbon (DOC).2. Recent observations of microbial community structure.3. Metabolic inference-based predictions of microbial metabolic potential.
Function
Trop
hic
Leve
l
PAL MCM
Pelagibacter Actinobacteria
Whale/seal Rotifer/tardigrade
The di�erence in the size of top predators at PAL (humpback whale, left) and MCM (rotifer, right) underscores major di�erences in ecosys-tem function. Such obvious di�erences, however, may mask function-al similarities that appear as we move toward basal trophic levels (left). Actinobacteria and Pelagibacter, the dominant bacteria at MCM and PAL respectively, for example, are both oligotrophic specialists with large functional overlaps.
1 m
ELB WLB FRX NPAL SPAL
Site/Region
BP
:PP
0.00
10.
010.
11
10
1 10 100 1000 10000
PP mg C m−2 day−1
BP
mg
C m
−2 d
ay−1
110
100
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●●
●●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●●
●
●● ●
●●
●
●
●
●
●
●●
●
●
●
●
● ●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
● ●
●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●
●
●
●●●●
●●●
●●
●
●
●
●
●
●
●
●● ●
● ●●●
●
●
●●
●
●●
●
●
● ●
●
●
●●
●●●●●
●
●
●
●
●
●
●
●●
●
●
●● ●
●●●
●●
●
●
●
●
●
●●
●
●
●●●
●
●●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
● ●●
●● ●●
●
●
●
●●
●
●
●●
●
●●●
●
●●
●
●●●
●●●
●
●
●
●
●●
●●●●
●
●●
●
●
●
●●●●
●●●●
● ●●
● ●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●●
●●
●●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●
● ●
●●
●
●●
●
●●
●●●
●
●
●●
●
●
●●●●●●●
●
●
● ●●●
●
●
●●
●●
●
●●
●●
●
●
●
●
●●
●●
●●
●●
●
●
●
●●
●●●
●●●●●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●
●●
●
● ●
●
● ●●
●●
●●
●
●●●
●
●
●●
●●
●
●
●●
●●●
●
●●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●●
●
PP mg C m−3 day−1
0.00
10.
010.
11
0.01 0.1 1 10
BP
mg
C m
−3 d
ay−1
FryxellEast Lobe BonneyWest Lobe BonneyNorth PalmerSouth Palmer1:101:5
+
+++ +
The ratio of BP to PP provides insight into the functioning of the microbial loop. Values in the global pelagic marine environment tend to center around 1:10 (grey dotted lines). At a ratio of 1:5 (black dotted lines) PP is thought to pro-vide insu�cient C to support both BP and respiration. Autochthonously �xed carbon must be subsidized by a source outside the photic zone. The three MCM lakes included in this investigation have BP:PP ratios exceeding PAL, despite their lower DOC concentrations. Lake Fryxell has extreme values of BP:PP suggesting a large subsidy from DOC-rich water below the photic zone.
The relationship between BP and PP
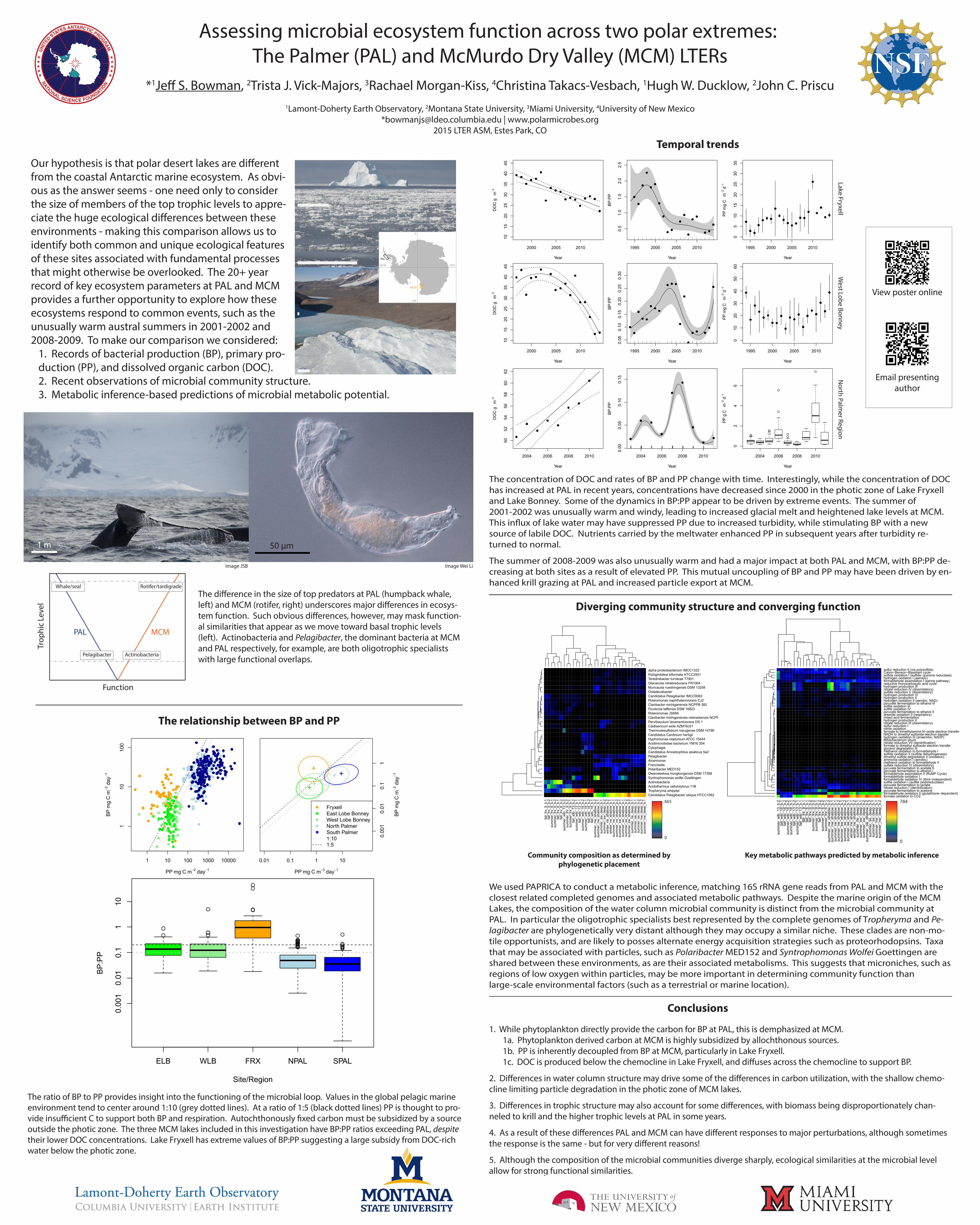
Temporal trends
The concentration of DOC and rates of BP and PP change with time. Interestingly, while the concentration of DOC has increased at PAL in recent years, concentrations have decreased since 2000 in the photic zone of Lake Fryxell and Lake Bonney. Some of the dynamics in BP:PP appear to be driven by extreme events. The summer of 2001-2002 was unusually warm and windy, leading to increased glacial melt and heightened lake levels at MCM. This in�ux of lake water may have suppressed PP due to increased turbidity, while stimulating BP with a new source of labile DOC. Nutrients carried by the meltwater enhanced PP in subsequent years after turbidity re-turned to normal.
The summer of 2008-2009 was also unusually warm and had a major impact at both PAL and MCM, with BP:PP de-creasing at both sites as a result of elevated PP. This mutual uncoupling of BP and PP may have been driven by en-hanced krill grazing at PAL and increased particle export at MCM.
Diverging community structure and converging function
fall_
frx_9
_b.1
fall_
frx_9
_b.2
sum
mer
_frx
_6_b
.1su
mm
er_f
rx_6
_b.2
fall_
frx_6
_b.1
fall_
frx_6
_b.2
sum
mer
_wlb
_13_
b.2
sum
mer
_frx
_9_b
.2su
mm
er_f
rx_9
_b.1
fall_
wlb
_13_
b.2
sum
mer
_wlb
_13_
b.1
fall_
wlb
_13_
b.1
sum
mer
_wlb
_18_
b.2
sum
mer
_wlb
_18_
b.1
fall_
wlb
_18_
b.2
fall_
wlb
_18_
b.1
sum
mer
_nw
_sha
llow
_b.2
sum
mer
_nw
_sha
llow
_b.1
sum
mer
_sw
_dee
p_b.
1su
mm
er_s
w_d
eep_
b.2
win
ter_
ne_s
hallo
w_b
.2w
inte
r_ne
_sha
llow
_b.1
sum
mer
_ne_
shal
low
_b.1
sum
mer
_ne_
shal
low
_b.2
sum
mer
_sw
_sha
llow
_b.1
sum
mer
_se_
shal
low
_b.1
sum
mer
_se_
shal
low
_b.2
sum
mer
_sw
_sha
llow
_b.2
sum
mer
_ne_
deep
_b.1
sum
mer
_se_
deep
_b.1
sum
mer
_se_
deep
_b.2
sum
mer
_nw
_dee
p_b.
2su
mm
er_n
w_d
eep_
b.1
Candidatus Pelagibacter ubique HTCC1062Tropheryma whippleiAcidothermus cellulolyticus 11BActinobacteriaSyntrophomonas wolfei GoettingenOwenweeksia hongkongensis DSM 17368Polaribacter MED152FrancisellaAlcanivoraxPelagibacterCandidatus Amoebophilus asiaticus 5a2CytophagiaAcidimicrobidae bacterium YM16 304Hyphomonas neptunium ATCC 15444Candidatus Cardinium hertigiiThermodesulfobium narugense DSM 14796Caldisericum exile AZM16c01Parvibaculum lavamentivorans DS 1Clavibacter michiganensis nebraskensis NCPPB 2581Polaromonas JS666Fluviicola taffensis DSM 16823Clavibacter michiganensis NCPPB 382Polaromonas naphthalenivorans CJ2Candidatus Pelagibacter IMCC9063OctadecabacterMuricauda ruestringensis DSM 13258Glaciecola nitratireducens FR1064Teredinibacter turnerae T7901Robiginitalea biformata HTCC2501alpha proteobacterium IMCC1322
665
0
sum
mer
_wlb
_13_
b.2
fall_
wlb
_18_
b.1
fall_
wlb
_18_
b.2
sum
mer
_wlb
_18_
b.2
sum
mer
_wlb
_18_
b.1
fall_
wlb
_13_
b.2
sum
mer
_wlb
_13_
b.1
fall_
wlb
_13_
b.1
fall_
frx_9
_b.1
fall_
frx_6
_b.1
fall_
frx_6
_b.2
sum
mer
_frx
_9_b
.2su
mm
er_f
rx_9
_b.1
fall_
frx_9
_b.2
sum
mer
_frx
_6_b
.2su
mm
er_f
rx_6
_b.1
sum
mer
_nw
_sha
llow
_b.2
sum
mer
_nw
_sha
llow
_b.1
sum
mer
_ne_
shal
low
_b.1
sum
mer
_ne_
shal
low
_b.2
sum
mer
_sw
_sha
llow
_b.1
sum
mer
_se_
shal
low
_b.2
sum
mer
_se_
shal
low
_b.1
win
ter_
ne_s
hallo
w_b
.1w
inte
r_ne
_sha
llow
_b.2
sum
mer
_ne_
deep
_b.1
sum
mer
_sw
_dee
p_b.
1su
mm
er_s
w_d
eep_
b.2
sum
mer
_sw
_sha
llow
_b.2
sum
mer
_se_
deep
_b.2
sum
mer
_se_
deep
_b.1
sum
mer
_nw
_dee
p_b.
1su
mm
er_n
w_d
eep_
b.2
formate oxidation to CO2
pyruvate fermentation to acetone
pyruvate fermentation to lactate
formaldehyde oxidation I
pyruvate fermentation to ethanol Ipyruvate fermentation to acetate II
methanol oxidation to formaldehyde II
methanol oxidation to formaldehyde Iglycerol degradation IIIformate to dimethyl sulfoxide electron transfer
Bifidobacterium shunt
NADH to dimethyl sulfoxide electron transfer
nitrite oxidationsulfur reduction I
hydrogen production Vmixed acid fermentation
pyruvate fermentation to ethanol IIsulfite oxidation IVsulfite oxidation IIIpyruvate fermentation to ethanol III
hydrogen production IIhydrogen production VI
hydrogen production IIIreductive monocarboxylic acid cycle
784
0
long
lat
180
0
90 W 90 E
PAL
MCM
We used PAPRICA to conduct a metabolic inference, matching 16S rRNA gene reads from PAL and MCM with the closest related completed genomes and associated metabolic pathways. Despite the marine origin of the MCM Lakes, the composition of the water column microbial community is distinct from the microbial community at PAL. In particular the oligotrophic specialists best represented by the complete genomes of Tropheryma and Pe-lagibacter are phylogenetically very distant although they may occupy a similar niche. These clades are non-mo-tile opportunists, and are likely to posses alternate energy acquisition strategies such as proteorhodopsins. Taxa that may be associated with particles, such as Polaribacter MED152 and Syntrophomonas Wolfei Goettingen are shared between these environments, as are their associated metabolisms. This suggests that microniches, such as regions of low oxygen within particles, may be more important in determining community function than large-scale environmental factors (such as a terrestrial or marine location).
Community composition as determined by phylogenetic placement
Key metabolic pathways predicted by metabolic inference
1. While phytoplankton directly provide the carbon for BP at PAL, this is demphasized at MCM. 1a. Phytoplankton derived carbon at MCM is highly subsidized by allochthonous sources. 1b. PP is inherently decoupled from BP at MCM, particularly in Lake Fryxell. 1c. DOC is produced below the chemocline in Lake Fryxell, and di�uses across the chemocline to support BP.
2. Di�erences in water column structure may drive some of the di�erences in carbon utilization, with the shallow chemo-cline limiting particle degradation in the photic zone of MCM lakes.
3. Di�erences in trophic structure may also account for some di�erences, with biomass being disproportionately chan-neled to krill and the higher trophic levels at PAL in some years.
4. As a result of these di�erences PAL and MCM can have di�erent responses to major perturbations, although sometimes the response is the same - but for very di�erent reasons!
5. Although the composition of the microbial communities diverge sharply, ecological similarities at the microbial level allow for strong functional similarities.
Conclusions
View poster online
Email presentingauthor
50 µm
Image Wei LiImage JSB