2 prova - fisio
TRANSCRIPT

FisiologiaFisiologia da dorA dor é um mecanismo de proteção corporal. Ela ocorre sempre que quaisquer tecidos estejam sendo lesados e provoca reação do indivíduo no sentido de remover o estímulo doloroso. Mesmo atividades simples tais como se assentar sobre o ísquio por longo período de tempo podem causar destruição tecidual, devido à perda do fluxo sanguíneo para a pele, no local em que ela fica comprimida pelo peso corporal. Quando a pele está dolorida devido à isquemia, a pessoa desvia seu peso daquela região para outra, de forma inconsciente. Entretanto, pessoas que tenham perdido o sentido da dor, tais como as que sofreram lesão da medula espinhal, são incapazes de percebê-la e não executam o movimento necessário, o que leva rapidamente à ulceração nas áreas de pressão.Os receptores da dor, na pele e nos outros tecidos, são terminações nervosas livres. Eles estão amplamente distribuídos nas camadas superficiais da pele e também de certos tecidos internos, tais como periósteo, paredes arteriais, superfícies articulares e na foice e tentório da calota craniana. A maior parte dos outros tecidos profundos tem inervação pobre de terminações sensíveis à dor; entretanto, uma lesão tecidual difusa e ampla pode, por somação, causar um tipo de dor contínua, crônica e lenta nessas áreas.Receptores mecanonociceptores: é excitado devido à intensa pressão sobre a pele.Receptores termonociceptores: estímulos térmicos de alta intensidadeReceptores polimodais: estímulos térmicos, químicos e mecânicos.Dor aguda (rápida): ocorre dentro de um tempo da ordem de 0,1 segundo após a aplicação do estímulo doloroso. Esse tipo de dor é sentido quando uma agulha é espetada na pele, ou quando a pele é cortada por uma lâmina, ou, ainda, quando a pele é submetida a um choque elétrico. Esse tipo de dor (rápida, em pontada) não é sentido na maior parte dos tecidoscorporais profundos. A dor do tipo rápida é transmitida pelas fibras dolorosas do tipo A delta.A delta --> mielinizada, possui limiar mais baixo que a fibra C, possui maior velocidade de condução e modula a primeira fase da dor.O feixe neoespinotalâmico possui neurônios de segunda ordem que são excitados por fibras de dor rápida do tipo A (delta), que transmitem estímulos nociceptivos mecânicos e térmicos. Algumas fibras deste feixe terminam nas áreas reticulares do tronco cerebral, porém muitas seguem até o tálamo. A partir daí, os sinais são transmitidos a outras áreas basais do encéfalo e ao córtex sensorial somático. Este sistema está envolvido com a discriminação, avaliação e rápida resposta à dor.Dor crônica (lenta): começa após 1 ou mais segundos depois da aplicação do estímulo, aumentando, então, lentamente por muitos segundos ou, às vezes, minutos. Esse tipo de dor está comumente associado à destruição tecidual. Ela pode tornar-se cruciante e levar a um sofrimento prolongado e insuportável. Esse tipo de dor pode ocorrer tanto na pele como em qualquer tecido ou órgão profundo. A dor lenta provém da estimulação das fibras do tipo C.fibra C -- > amielínica ou pouco mielinizada, maior diâmetro, velocidade de condução lenta e é responsável pela dor difusa.O feixe paleoespinotalâmico transmite a dor principalmente por fibras periféricas do tipo C (dor lenta). Poucas fibras chegam até o tálamo, porém a maioria termina em múltiplas áreas do bulbo, ponte e mesencéfalo. O sistema paleoespinotalâmico possui ligações com áreas que determinam aspectos motivacionais e afetivos que influenciam na percepção dolorosa. Ao contrário da maior parte dos outros receptores sensoriais do organismo, a adaptação dos receptores de dor, quando ocorre, é mínima. A excitação das fibras sensíveis à dor se torna progressivamente maior à medida que o estímulo doloroso continua. Esse aumento da sensibilidade dos receptores de dor é chamado de hiperalgesia. Esse fenômeno é importante porque possibilita que a pessoa permaneça sempre alerta ao estímulo lesivo, que provoca a dor durante todo o tempo em que ele persiste. Essa sensibilidade é ocasionada porque quando o tecido é lesado libera varias substâncias químicas (bradicinina, substância P, prostaglandinas, K+, H+) que desencadeiam a resposta inflamatória. Com isso mastócitos liberam histamina, que

ativa diretamente os nociceptores. Isso ocasiona a diminuição do limiar do potencial de ação fazendo com que um simples toque ocasione a dor no tecido, o que normalmente não ocasionaria. Os feixes nervosos ascendentes da dor são os neoespinotalâmico e o paleoespinotalâmico. A transmissão dos estímulos nociceptivos até a medula espinhal é feita pelos nervos periféricos. No tronco e nos membros, é feita através dos nervos espinhais; nas vísceras pelos nervos simpáticos, parassimpáticos e esplâncnicos. Na região da cabeça é transmitida principalmente pelo nervo trigêmeo.As células que sinalizam impulsos viscerais, também transportam informação de receptores cutâneos, fazendo com que o cérebro identifique de forma errônea a fonte de dor (dor referida). Existem várias teorias que tentam explicar a dor referida: • A possibilidade de uma única célula nervosa fazer sinapse com várias outras e vice-versa gera um fenômeno conhecido como convergência ou divergência sináptica. •Cada metâmero recebe inervação oriunda do segmento espinhal correspondente. Com o desenvolvimento do organismo, esta disposição metamérica se modifica e o resultado disto é que na vida adulta a pele e o músculo de um membro, por exemplo, acabam ficando distantes da víscera à qual estavam associados, mas são inervados pela mesma raiz nervosa, daí a dor do infarto ser referida ao braço e à mão.A dor fantasma é uma sensação dolorosa referente ao membro (ou parte dele) perdido que pode se apresentar de diversas formas tais como ardor, aperto, compressão ou até mesmo uma dor intensa e frequente. Mas é claro que esse tipo de dor não é causado pelo estímulo de nociceptores na área em que a dor é sentida, pois eles não estão mais presentes. Quando ocorre a perda da inervação sensorial de uma região ou a amputação de um membro as informações sensoriais periféricas se tornam inteiramente ausentes, fazendo com que neurônios no sistema nervoso central que até então recebiam informações daquela parte do corpo se tornem anormalmente hiperativos. Importantes evidências de que o crescimento pós-lesão e o novo padrão de conexões estabelecidas por neurônios no cérebro de amputados podem ser possível causa do problema.Modulação da dor - Algesia – Vias descendentes da dorFoi descrito um sistema modulador da dor envolvendo a substância cinzenta periaquedutal. O grau em que uma pessoa reage à dor é altamente variável. Isto resulta, em parte, da capacidade do próprio cérebro em suprimir a entrada de sinais de dor ao sistema nervoso ativando um sistema de controle da dor, chamado sistema de analgesia. Este sistema é constituído por três componentes principais: • um complexo inibitório da dor localizado nas pontas dorsais da medula espinhal, • núcleo magno da rafe localizado na ponte, • área cinzenta periaquedutal e periventricular do mesencéfalo. Os principais inibidores da transmissão nociceptiva são os peptídeos opióides, a serotonina e a noradrenalina. Os opióides são liberados pela ACP para atuar no núcleo magno da rafe, lócus coerulus e subcoerulus, desempenhando um papel relevante no sistema modulador descendente. Muitas fibras nervosas que se originam nos núcleos periventriculares e na área cinzenta periaquedutal secretam encefalina nas suas terminações nervosas que alcançam o núcleo magno de rafe. Deste núcleo partem fibras que se dirigem para as pontas dorsais da medula espinhal, onde secretam serotonina e estimulam os neurônios locais a secretarem encefalina. Acredita-se que a encefalina cause inibição pré-sináptica nas fibras da dor quando estas fazem sinapse nas pontas dorsais. A inibição pré-sináptica é provavelmente realizada pelo bloqueio de canais de cálcio nos terminais, provocando a diminuição da entrada de cálcio e, consequentemente, a quantidade de neurotransmissor liberado diminui. A encefalina também age no neurônio de segunda ordem impedindo a abertura dos canais de Ca++ ativados pelo glutamato. O bloqueio parece durar períodos prolongados porque, após a ativação deste sistema, a analgesia dura vários minutos até horas.Sempre que uma informação dolorosa trafega pela via espinotalâmica, aferências chegam até a substância cinzenta periaquedutal, podendo ativar a via de analgesia endógena. Porém, normalmente, essa ativação não é suficiente para impedir a passagem do estímulo nociceptivo para o neurônio de segunda ordem, pois, se assim o fizesse não sentiríamos dor.

Por outro lado, a substância cinzenta periaquedutal recebe muitas aferências excitatórias provenientes principalmente do hipotálamo e, sendo o hipotálamo a via de saída das eferências do sistema límbico, é possível entender o porquê da grande influência (positiva ou negativa) que o estado emocional do indivíduo exerce sobre a intensidade da dor percebida. Por exemplo, soldados feridos na guerra que continuam a batalha e relatam não sentir dor até que o combate termine; atletas contundidos que continuam a prova e percebem o ferido somente após o término da competição. Opostamente, sensibilidade à dor aumentada em muitas pessoas deprimidas e ansiosas.A ação pré-sináptica, nos terminais aferentes se dá pelo impedimento da liberação de transmissores excitatórios. A ação pós-sináptica produz potencial pós-sináptico inibitório. ReflexosA musculatura esquelética tem a capacidade de realizar fundamentalmentetrês tipos de movimentos que são:x Movimento reflexo: movimentos mais simples, dependentes de informações sensoriais e muito pouco do movimento voluntário. Um bom exemplo é o reflexo de retirada pela dor (arco reflexo).x Movimento rítmico/autônomo: são de ordem um pouco mais complexa, dependem de atos voluntários para o início e término do movimento. Um exemplo é a marcha e a mastigação.x Movimento voluntário: são os movimentos mais complexos, tem uma numerosa diversificação e são altamente dependentes do aprendizado. A escrita é um bom exemplo deste controle. Esses reflexos podem ser do tipo:- somático: aferente é a superfície e o órgão efetor são os músculos esqueléticos.- visceral: aferente é a vísceras e o efetor são as próprias vísceras ou outras.- viscero-somático: aferente é a víscera e o efetor é o somático. Ex: dor no apêndice.- somato-visceral: aferente é o soma e o efetor é a víscera. Ex: estimulação na pele pode causar aumento da frequência cardíaca. Os reflexos da medula espinhal são respostas motoras estereotipadas para tipos específicos de estímulos, como o estiramento do musculo. O circuito neuronal que conduz essas respostas motoras é chamado de arco reflexo. O arco reflexo inclui os receptores sensoriais, os neurônios aferentes sensoriais, que conduzem informação para a medula espinhal; os interneurônios na medula espinhal; e os motoneurônios, que direcionam o músculo a se contrair ou a se relaxar. Existem dois tipos de motoneurônios: motoneurônios α e motoneurônios γ. Os motoneurônios α inervam fibras musculares esqueléticas extrafusais. Os potenciais de ação, nos motoneurônios α levam a potenciais de ação nas fibras musculares extrafusais que inervam, resultando em contração. Os motoneurônios γ inervam fibras musculares intrafusais especializadas, um componente dos fusos musculares. A função geral do fuso muscular é a de detectar o comprimento do músculo; a função dos motoneurônios γ que os inervam é ajustar a sensibilidade do fuso muscular (permitindo a resposta, se as fibras extrafusais se contraem e se encurtam). As fibras extrafusais compreendem a maioria das fibras dos músculos esqueléticos e são usadas para gerar força. As fibras intrafusais são fibras especializadas, encapsuladas em bainha formando fusos em paralelo às fibras extrafusais.Reflexos somáticos • Postura {reflexo miotático ou de estiramento {reflexo miotático inverso
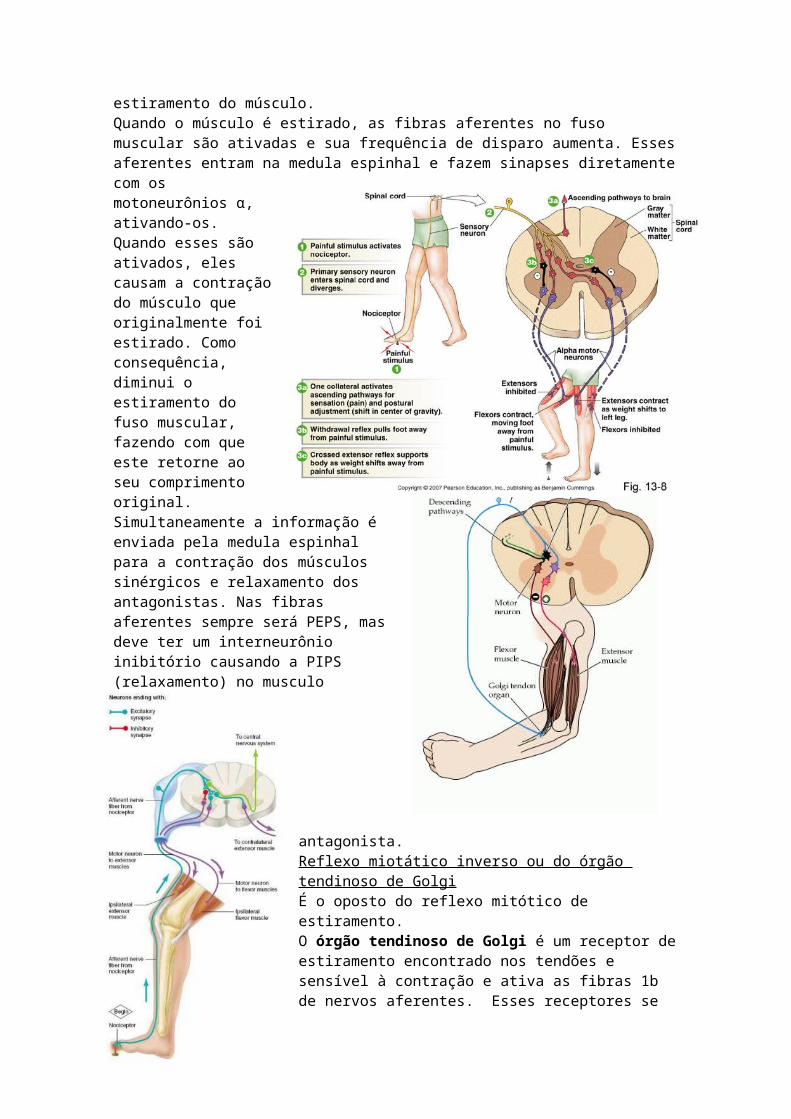
• Defesa {reflexo de retirada ou flexão {reflexo de extensão cruzadaReflexo miotático ou de estiramentoO resultado é que o arco reflexo é uma contração induzida por estiramento do músculo.Quando o músculo é estirado, as fibras aferentes no fuso muscular são ativadas e sua

frequência de disparo aumenta. Esses aferentes entram na medula espinhal e fazem sinapses diretamente com os motoneurônios α, ativando-os. Quando esses são ativados, eles causam a contração do músculo que originalmente foi estirado. Como consequência, diminui o estiramento do fuso muscular, fazendo com que este retorne ao seu comprimento original. Simultaneamente a informação é enviada pela medula espinhal para a contração dos músculos sinérgicos e relaxamento dos antagonistas. Nas fibras aferentes sempre será PEPS, mas deve ter um interneurônio inibitório causando a PIPS (relaxamento) no musculo antagonista.Reflexo miotático inverso ou do órgão tendinoso de GolgiÉ o oposto do reflexo mitótico de estiramento. O órgão tendinoso de Golgi é um receptor de estiramento encontrado nos tendões e sensível à contração e ativa as fibras 1b de nervos aferentes. Esses receptores se encontram em serie com as fibras musculares extrafusais.Quando o musculo se contrai, as fibras musculares extrafusais se encurtam, ativando os órgãos tendinosos de Golgi acoplados a elas. Em resposta, as fibras aferentes do grupo 1b que fazem sinapse com interneurônios inibitórios na medula espinhal, são ativados.
Esses interneurônios inibitórios fazem sinapses com os motoneurônios α, inibindo a descarga dos neurônios α, produzindo o relaxamento do musculo (antes contraído). Como o músculo homônimo relaxa, o reflexo provoca o relaxamento dos músculos sinérgicos e a contração dos músculos antagonistas.Reflexo de retirada ou flexãoÉ um reflexo polissináptico que ocorre em resposta a estímulos dolorosos e nocivos. As fibras aferentes somatossensoriais e as de dor iniciam o reflexo flexor, que causa a retirada da parte afetada do corpo do estimulo doloroso ou nocivo. Esse reflexo produz flexão no lado do estimulo e extensão do lado oposto. Quando um membro toca um estimulo doloroso (fogão

quente), as fibras aferentes do reflexo flexor são ativadas. Essas fibras fazem sinapses com múltiplos interneurônios na medula espinhal. Do lado do estimulo doloroso provoca a contração dos músculos flexores e o relaxamento dos músculos extensores (retirada da mão do fogão quente).Reflexo de extensao cruzadaO membro contralateral sofrerá o oposto. Assim se houver o estimulo do lado esquerdo, o braço e a perna esquedos serao flexionados (retirados) e o braço e perna direitos se estenderão para manter o equilíbrio. Fisiologia endócrina O sistema endócrino, em conjunto com o sistema nervoso é responsável pela homeostasia. O crescimento, desenvolvimento, reprodução, pressão sanguínea, concentraçao de íons e de outras substâncias no sangue são controlados pelo sistema endócrino. O hormônio é uma substância química classificada como peptídeo, esteróide ou amina. Os hormônios são secretados para a circulação em pequenas quantidades e levados até os tecidos-alvo, onde produzem respostas fisiológicas. Síntese hormonalSíntese de hormônios peptídicos e protéicosNo núcleo da célula o DNA é transcrito em RNAm, que é traduzido em um pré-hormônio. No RE o pré-hormônio é convertido em pró-hormônio. O pró-hormônio é acondicionado em vesículas secretoras. Nas vesículas secretoras, enzimas clivam as sequências peptídicas, produzindo o hormônio final. Esse hormônio é armazenado nas vesículas secretoras até a célula endócrina ser estimulada.Síntese de hormônios esteróidesSão sintetizados e secretados pelo córtex supra-renal, pelas gônadas, corpo lúteo e pela placenta. Todos esses hormônios derivam do colesterol, que é modificado pela remoção ou adição de cadeias laterais, hidroxilação ou aromatização do núcleo esteroide. Síntese dos hormônios amínicos São catecolaminas (epinefrina, norepinefrina e dopamina) e os hormônios tireoidianos. Esses hormônios são derivados da tirosina. Regulação da secreção hormonalPara que haja a homeostasia, a secreção dos hormônios precisa ser regulada. Essa regulação pode ocorrer por mecanismos neuronais (nervos simpáticos pré-ganglionares fazem sinapse na medula supra-renal e, quando estimulados, causam secreção das catecolaminas para circulação) ou por mecanismos de feedback (o elemento da resposta ao hormônio retroalimenta, direta ou indiretamente – pode ser positivo ou negativo).Feedback negativoNos sistemas endócrinos, o feedback negativo faz com que alguma característica da ação hormonal, direta ou indiretamente, iniba a secreção posterior do hormônio.O feedback de alça longa significa que o feedback do hormônio regula todo eixo hipotalâmico-hipofisário. O feedback de alça curta faz com que o hormônio da hipófise anterior regule a
secreção do hormônio hipotalâmico. O feedback de alça ultracurta o hormônio hipotalâmico inibe sua própria secreção.O resultado efetivo de qualquer feedback negativo é que, quando os níveis hormonais são considerados adequados ou altos, a secreção posterior do hormônio é inibida. Quando os níveis hormonais são considerados inadequados ou baixos, a secreção do hormônio é estimulada.Feedback positivoO feedback positivo alguma característica da ação hormonal provoca mais secreção do hormônio. Feedback positivo é autocrescente conduzindo um evento explosivo.Regulação de receptores hormonaisPara responder, um tecido-alvo deve conter receptores específicos que reconheçam o hormônio. Tais receptores são acoplados a mecanismos celulares que produzem a resposta fisiológica. A sensibilidade do tecido-alvo pode ser alterada por duas maneiras: alterando o numero de receptores ou alterando a afinidade dos receptores para um hormônio. A regulação para menos diminui o numero de receptores ou sua afinidade pelo hormônio. A regulação para mais aumenta o numero de receptores ou a afinidade pelo hormônio.Mecanismo de ação hormonal e segundos mensageirosAs ações hormonais sobre as células-alvo começam quando o hormônio se liga a um receptor de membrana, formando o complexo hormônio-receptor. Em muitos sistemas, esse complexo está acoplado a proteínas efetoras. Essas proteínas são enzimas (Adenil ciclase ou fosfolipase C). Quando essas proteínas são ativadas é produzido um segundo mensageiro, AMPc ou IP que amplifica o sinal inicial do hormônio e orquestra as ações fisiológicas.Proteína GSão proteínas ligadas a membrana que acoplam os receptores de hormônios às enzimas efetoras. Assim, servem como “interruptores moleculares” que decidem se pode ou não ocorrer a ação hormonal. Quando a GTP se liga a proteína ela é ativada; quando a GDP está ligada é inativada. Isso significa que quando o GTP se liga à essa proteína estimula a adenilil ciclase (enzima efetora).- Mecanismo adenilil ciclase (AMPc)O mecanismo envolve a ligação de um hormônio a um receptor acoplado à proteína G ou Gi, e, então, há ativação ou inibição da adenilil ciclase, levando ao aumento ou diminuição intracelular de AMPc. Em seguida, esse segundo mensageiro amplifica o sinal do hormônio, produzindo as respostas fisiológicas.• hormônio se liga ao seu receptor na membrana celular, fazendo com que o GDP seja liberado e substituído pela GTP;• O GTP migra e se liga e ativa a adenilil cliclase, essa catalisa a conversão do ATP em AMPc (segundo mensageiro);• Esse AMPc por diversas etapas ativa a proteína cinase A, que fosforila proteínas intracelulares. Essas proteínas fosforiladas executarão as ações fisiológicas.• Esse AMPc é degradado a AMP (metabólito inativo) pela enzima fosfodiesterase, quando há término da ação do segundo mensageiro.- Mecanismo da fosfolipase CEsse mecanismo envolve a ligação do hormônio ao receptor e o acoplamento por meio da fosfolipase C. Os níveis intracelulares de IP3 e de Ca2+ são aumentados, produzindo as repostas finais.• O hormônio se liga ao receptor que libera a GDP e é substituído pelo GTP.• O complexo com GTP ativa a fosfolipase C. Essa fosfolipase C ativa a liberação de diacilglicerol e IP3 do PIP2 (fosfolipídio de membrana). O IP3 gerado provoca liberação de Ca2+ das reservas intracelulares no reticulo endoplasmático, resultando no aumento da concentração intracelular de Ca2+;• O Ca2+ e diacilglicerol ativam a proteína cinase C, que fosforila proteínas provocando as

respostas fisiológicas.Relações hipotalâmico-hipofisáriasO hipotálamo e a hipófise funcionam de modo coordenado, comandando a maioria dos sistemas endócrinos. A hipófise posterior é chamada de neuro-hipófise. A hipófise anterior é chamada de adeno-hipófise. O hipotálamo controla a hipófise por mecanismos neurais e hormonais.- Relação entre hipotálamo e hipófise posterior (neuro-hipófise) A hipófise posterior deriva do tecido neural e secreta dois hormônios peptídicos, o hormônio anti-diurético (ADH) e ocitocina. A hipófise posterior é uma coleção de axônios cujos corpos celulares estão localizados no hipotálamo. Uma vez sintetizados nos corpos celulares, os hormônios são transportados ao longo dos axônios em vesículas neurossecretoras e armazenados na terminação da hipófise posterior. Quando o corpo é estimulado, as vesículas são liberadas dos nervos terminais por exocitose e esse hormônio entra em capilares fenestrados. O sangue venoso da hipófise posterior entra na circulação sistêmica, que leva os hormônios até seus tecidos-alvo. A relação entre o hipotálamo e a hipófise posterior é direta.- Relação entre hipotálamo e hipófise anterior (adeno-hipófise)A hipófise anterior é uma coleção de células endócrinas. Ela secretada seis hormônios: hormônio estimulante da tireóide (TSH), hormônio folículo-estimulante (FSH), hormônio luteinizante (LH), hormônio do crescimento, prolactina e hormônio adrenocorticotrópico (ACTH). O hipotálamo e a hipófise anterior estão ligados diretamente pelos vasos sanguíneos porta hipotalâmico-hipofisários. Isso permite que os hormônios hipotalâmicos sejam levados à hipófise anterior de modo direto e em altas concentrações, e os hormônios hipotalâmicos não aprecem na circulação sistêmica em altas concentrações.Fisiologia Reprodutora femininaAs gônadas femininas são os ovários, juntamente com o útero e as tubas uterinas, compõe o sistema reprodutor feminino. Os ovários possuem função de oogênese e secreção de hormônios esteroides sexuais femininos (progesterona e estrogênio). Os hormônios esteroides possuem funções parácrina e endócrina. Localmente, esses hormônios atuam sustentando o desenvolvimento do óvulo. Sistemicamente, esses hormônios atuam no útero, mamas e osso. A unidade funcional dos ovários é o folículo ovariano, composto por uma célula germinativa circundada por células endócrinas. Quando totalmente desenvolvido este fornece nutrientes para o oócito em desenvolvimento, libera o oócito no tempo apropriado (ovulação), prepara a vagina e as tubas uterinas para auxiliarem na fertilização do óvulo por um espermatozoide, prepara o revestimento do útero para a implantação do zigoto e matem a produção de hormônios esteróides até que até que a placenta assuma esse papel.OogêneseHá o crescimento dos folículos ovarianos, o qual apenas um folículo maduro domina sobre todos os outros, ocorrendo regressão ou atresia dos outros. Após isso ocorre a ovulação (folículo se rompe e libera oócito). Após a penetração nas tubas uterinas começa a segunda divisão meiótica, que só é terminada após a fertilização. Os elementos residuais do folículo formam o corpo lúteo. Este se compõe, de células da granulosa, células da teca, capilares e fibroblastos. O corpo lúteo sintetiza e secreta hormônios esteroides, que são necessários para implantação e manutenção do zigoto. Se houver fecundação, o corpo lúteo secretará hormônios ate que a placenta assuma esse papel. Se não houver fecundação, ele regride sendo substituído por uma cicatriz chamada corpus albicans.Síntese e secreção de estrogênio e progesteronaOs hormônios esteroides ovarianos, progesterona e 17β-estradiol, são sintetizados nos folículos ovarianos pelas células da granulosa e células da teca.As células da teca sintetizam e secretam progesterona e também sintetizam testosterona, esse hormônio se difunde das células da teca para as células da granulosa que contêm aromatase. Nas células da granulosa, a testosterona é convertida em 17β-estradiol. O LH estimula a colesterol desmolase nas células da teca e o FSH estimula a aromatase das células da

granulosa.GnRHA função ovariana na mulher é controlada pela atividade pulsátil do eixo hipotalâmico-hipofisário. O GnRH é levado à adeno-hipófise em altas concentrações através dos vasos sanguíneos, onde estimula a secreção pulsátil de FSH E LH.FSH E LHFSH --> as células da granulosa são as únicas que possuem receptores para esse hormônio. Ele estimula o crescimento das células da granulosa e estimulam a síntese do estradiol. Consequentemente haverá mais células, mais estradiol, mais células.LH --> a ovulação se inicia pelo LH. Antes da ovulação as concentrações sanguíneas de LH aumentam e induzem a ruptura do folículo dominante, liberando o oócito. Estimula também a formação do corpo lúteo e mantém a produção do hormônio esteróide.Controle hormonal pode ser do tipo positivo ou negativo dependendo da fase do ciclo menstrualFolicular: o FSH e LH estimulam a síntese e secreção de estradiol pelas células foliculares. Assim esse aumento de estradiol atua como inibidor sobre as células da adeno-hipofise e hipotálamo, inibindo a secreção adicional de FSH, LH e GnRH respectivamente.Meio do ciclo: os níveis de estradiol aumentam como resultado da proliferação das células foliculares. Quando atingido o nível critico desse hormônio, o estradiol passa a ter efeito feedback positivo na adeno-hipofise, ou seja, causa secreção adicional de FSH e LH. Isso desencadeia a ovulação.Lútea: ocorre secreção de progesterona, que causa feedback negativo sobre a adeno-hipofise, inibindo secreção de FSH e LH. Progesterona e estrógeno Útero: estrogênio provoca proliferação e crescimento celular e contratilidade; a progesterona aumenta a atividade secretora e diminui a contratilidade. Tubas uterinas: estrogênio estimula a atividade ciliar e a contratilidade; a progesterona aumenta a atividade secretora e diminui a contratilidade.Vagina: estrogênio estimula proliferação das células epiteliais; a progesterona a diferenciação, mas inibe a proliferação das células epiteliais.Ciclo menstrualFase proliferativa: durante esse período há o desenvolvimento do folículo. No inicio dessa fase, os receptores para FSH e LH aumentam nas células da teca e da granulosa, estimulando a síntese de estradiol. Os altos níveis de estradiol causam a proliferação do revestimento endometrial do útero e inibem a secreção de FSH e LH.O estrogênio é secretado no folículo e induz a formação de númeroscada vez maiores de receptores de FSH pelas células dagranulosa; esse processo estabelece um efeito de feedback positivoao tornar as células da granulosa mais sensíveis do que nuncaao FSH da adeno-hipófiseOvulação: ocorre no 14 dia. Ela ocorre devido ao aumento de secreção de estradiol, ao final da fase folicular, que há feedback positivo sobre a secreção de FSH E LH. O surto de FSH e LH causa a ovulação. Na ovulação o muco vaginal aumenta em quantidade, ficando aquoso e mais penetrável ao esperma. Os níveis de estradiol diminuem logo após a ovulação, mas aumentarão durante a fase lútea.Fase secretora: começa após a ovulação e termina com o inicio da menstruação. Durante essa fase o corpo lúteo se desenvolve e secreta estradiol e progesterona. Os altos níveis de progesterona estimula a atividade secretora do endométrio e aumenta a vascularização, ou seja, prepara o endométrio para o ovulo fertilizado. O muco torna-se menos abundante e mais espesso. Se não ocorrer a fertilização o corpo lúteo regride, com isso os níveis sanguíneos de estradiol e progesterona caem abruptamente.Menstruação: a regressão do corpo lúteo e a perda abrupta de estradiol e progesterona

causam a eliminação do endométrio e de sangue. Durante esse tempo, os folículos primordiais para o próximo ciclo estão sendo recrutados e começando a se desenvolver.Fisiologia Reprodutora Masculina Os testículos possuem duas funções: espermatogênese e secreção de testosterona. Parte dos testículos é composto por túbulos seminíferos que consistem nas espermatogônias (células tronco), espermatócitos (células em processo de se tornarem espermatozoides) e células de Sertoli (sustentam os espermatozoides em desenvolvimento). Células de Sertoli - possuem três funções importantes: fornecem nutrientes aos espermatozoides que estão se diferenciando; formam junções comunicantes entre si, criando uma barreira entre os testículos e a corrente sanguínea (barreira hematotesticular); e secretam solução aquosa no lúmen dos túbulos seminíferos, que ajudam a transportar os espermatozoides dos túbulos para o epidídimo. Células de Leydig – possui função de síntese e secreção de testosterona. A testosterona possui efeitos locais (parácrinos) que sustentam a espermatogênese nas células de Sertoli, e efeitos endócrinos em outros órgãos-alvo (músculo esquelético e próstata).Síntese e secreção de testosteronaA testosterona é o principal androgênio, é sintetizada e secretada pelas células de Leydig. As vias são parecidas com as do córtex supra-renal, com diferenças importantes: nos testiculos, faltam enzimas 21β-hidroxilase e 11β-hidroxilase e por isso não podem sintetizar os glicocorticóides ou mineralocorticóides; e os testículos possuem enzima 17β-hidroxiesteróide desidrogenase (converte androstenediona em testosterona). Em alguns tecidos-alvo, a diidrotestosterona é o adrogênio ativo. Portanto nesses tecidos, a testosterona é convertida em diidrotestoterona pela enzima 5α-redutase. A maior parte de testosterona são ligadas à globulina ou albumina. Já as que não estão ligadas, são biologicamente ativas. As proteínas plasmáticas atuam como reservatório do hormônio circulante. A síntese dessas proteínas são estimuladas pelos estrogênios e inibida pelos androgênios.Regulação dos testículos GnRHÉ secretado pelos neurônios hipotalâmicos. O GnRH é secretado no sistema porta hipotalâmico-hipofisário, levado em altas concentraçoes diretamente até a adeno-hipófise. Durante o período reprodutivo, a secreção de GnRH é pulsátil e comanda um padrão secretor pulsátil semelhante na adeno-hipófise.FSH e LHSão hormônios liberados na adeno-hipófise que estimulam os testículos a executarem suas funções espermatogênicas e endócrinas. O FSH estimula a espermatogênese e a função das células de Sertoli. O LH estimula as células de Leydig a sintetizar testosterona, por aumentar a atividade da colesterol desmolase.A testosterona, secretada pelas células de Leydig possui efeitos endócrinos ou parácrinos. Nos testículos a testosterona se difunde das células de Leydig para as células de Sertoli, onde reforça a ação espermatogênica do FSH. Fora dos testículos, ela é levada pela circulação aos tescidos-alvo.Feedback negativoO eixo hipotalâmico-hipofisário é controlado por feedback negativo. A primeira via, a própria testosterona forma feedback com hipotálamo e com adeno-hipófise, inibindo secreção de GnRH e LH. No hipotálamo ela diminui a frequência e amplitude dos pulsos de GnRH. Na segunda via, as células de Sertoli secretam inibina. Essa é uma glicoproteína que inibe a secreção de FSH pela adeno-hipófise.Ações dos andrógenos Testosterona: é responsável pela diferenciação fetal do sistema genital interno (epidídimo, canal deferente e vesículas seminais). Na puberdade ela aumenta a massa muscular, promove o crescimento acelerado de pelos pubianos, fechamento das placas epifisárias, crescimento do pênis e vesículas seminais, mudança de voz, espermatogênese e libido.

Diidrotestosterona: é responsável pela diferenciação fetal da genitália externa (pênis, saco escrotal e próstata), pela distribuição dos pelos corporais, atividade das glândulas sebáceas e pelo crescimento da próstata.Inibidores da 5α-redutase: bloqueiam a conversão da testosterona em diidrotestosterona, bloqueando a produção de androgênios ativos em alguns tecidos-alvo. Como o crescimento da próstata e o padrão masculino de calvície dependem da diidrotestosterona, os inibidores da 5α-redutase podem ser usados no tratamento da hipertrofia prostática benigna e na perda de cabelos em homens.Mecanismo de ação: começa pela ligação da testosterona ou diidrotestosterona a uma proteína receptora de androgênio nas células dos tecidos-alvo. O complexo androgênio-receptor desloca-se para o nucleo, iniciando a transcrição gênica. São gerados novos RNAms e traduzidos em novas proteínas, responsáveis pela várias ações fisiológicas dos andrôgênios.Glândula adrenalA medula supra-renal é uma zona mais interna da glândula. Sua origem é neuroectodérmica e secreta catecolaminas, epinefrina e norepinefrina.O córtex supra-renal, se localiza na zona mais externa da glândula e possui três camadas distintas. Essas partes secretam hormônios esteróides adrenocorticais. O precursor para todos os esteróides adrenocorticais é o colesterol. Este é fornecido ao córtex supra-renal pela circulação. O colesterol circula ligado a lipoproteínas de baixa densidade. Há receptores para essas lipoproteínas nas membranas das células adrenocorticais, quando há essa ligação o colesterol se transfere para a célula por endocitose. Nas células, esse colesterol é esterificado e armazenado em vesículas citoplasmáticas, até que haja necessidade de síntese dos hormônios esteróides.Zona reticular (mais interna): pequenas quantidades de cortisol e hormônios sexuais (adrogênio supra-renais - DHEA e androstenediona). Esses compostos possuem atividade androgênica fraca, mas nos testículos são convertidos em testosterona. Há formação desses androgênios pela presença da enzima 17,20-liase. Nos homens, os androgênios supra-renais têm pouco significado; os testículos produzem sua própria testosterona. Contudo, nas mulheres, o córtex supra-renal é a principal fonte de compostos androgênicos.Zona fascicular (intermediária): sintetiza e secreta glicocorticóides (cortisol e corticosterona). Portanto, essa zona possui todas as enzimas necessárias para converter o colesterol em cortisol. O cortisol não é o único esteróide com atividade glicocorticóide, a corticosterona também é. Assim, o cortisol não é, necessário para manter a vida, desde que a corticosterona esteja sendo sintetizada. Se nenhuma das duas for sintetizada pode levar o indivíduo à morte.Zona glomerular (externa): sintetiza e secreta mineralocorticóides (aldosterona). A etapas para converter o colesterol em corticosterona são idênticas às da zona fascicular e a adição da aldosterona sintase à zona glomerular converte a corticosterona em aldosterona. A 11-desoxicorticosterona (DOC) e corticosterona também possuem atividade minaralocorticóide.Regulação da secreção dos esteróides adrenocorticaisA síntese e secreção dos hormônios esteróides pelo córtex supra-renal dependem da estimulação da colesterol desmolase pelo ACTH. Na ausência de ACTH, cessa a biossíntese dos hormônios esteróides adrenocorticais.A secreção de cortisol é de natureza pulsátil e possui padrão diurno. Ocorre em média 10 surtos secretores durante um período de 24 horas. As intensidade mais baixas de secreção ocorrem durante a noite e logo após adormecer, e as mais altas ocorrem imediatamente e antes do despertar. O principal surto secretório antes do despertar é responsável por metade da secreção diária total do hormônio. A liberação de ACTH também apresenta padrão diurno.O CRH é secretado pelo hipotálamo e atua sobre as células corticotróficas da adeno-hipófise, provocando a secreção de ACTH. Este atua nas células do córtex supra-renal, estiumulando a síntese e secreção dos hormônios adrenocorticais. O CRH segue para a hipófise pelo sangue porta hipotalâmico-hipofisário. Na adeno-hipófise atua sobre as células corticotróficas por adenilil ciclase/AMPc, provocando secreção de ACTH na corrente sanguínea. Após ligação do

ACTH aos receptores das células no córtex supra-renal há estimulação da transferência do colesterol armazenado para as mitocôndrias, ativação da colesterol desmolase. Outros efeitos é a estimulação da transcrição de genes aumentando receptores de ACTH. Níveis elevados de ACTH incluem hipertrofia e hiperplasia das células do córtex supra-renal.O “relógio interno” que induz o padrão diurno pode ser modificado, alternando-se o ciclo sono-vigília. O padrão diurno é abolido pelo coma, cegueira ou exposição à luz ou à escuridão.Feedback negativoÉ exercido pelo cortisol que pode inibir diretamente a secreção de CRH pelo hipotálamo, inibe a secreção de CRH através de neurônios que fazem sinapses no hipotálamo, inibindo a ação do CRH na adeno-hipófise resultando na inibição do ACTH.Regulação da aldosteronaA principal regulação da aldosterona não ocorre pelo ACTH, mas por variações do volume do LEC através do sistema renina-angiotensina-II aldosterona e por variações dos níveis séricos de potássio (K+).Ações dos glicocorticóidesEstimulação da glicogênese: promove a gliconeogênese e o armazenamento do glicogênio. Afetam o metabolismo proteico, lipídico e dos carboidratos, aumentando a síntese de glicose, ou seja, possuem efeitos catabólicos e diabetogênicos. O cortisol diminui a utilização de glicose pelos tecidos e a sensibilidade à insulina no tecido adiposo. São essenciais para a sobrevivência durante o jejum, por estimularem essas vias gliconeogênicas. Efeitos anti-inflamatórios: induz a síntese de lipocortina, inibindo a síntese do percussor das prostaglandinas e leucotrienos (mediadores da resposta inflamatória), inibe produção de interleucina-2 e proliferação de linfócitos T; e inibe liberação de histamina e serotonina.Supressão da resposta imune: inibe a produção de interleucina-2 e proliferação de linfócitos T. Podem ser administrados para suprimir a resposta imune e impedir a rejeição de órgãos transplantados. Manutenção da reatividade vascular às catecolaminas: é necessário para manutenção da pressão sanguínea normal e desempenha papel permissivo nas arteríolas por receptores adrenérgicos. Portanto, é necessário para a resposta vasoconstritora das arteríolas às catecolaminas.Inibição da formação de osso: diminui a síntese de colágeno, por diminuir a produção de osteoblastos e absorção intestinal de Ca2+.Aumento da taxa de filtração glomerular: o cortisol aumenta o TFG por causar vasodilatação das arteríolas, aumentando o fluxo sanguíneo renal.Efeitos sobre o SNC: são encontrados receptores de glicocorticóides no sistema límbico. O cortisol diminui o sono, aumenta o sono de ondas lentas e aumenta o tempo de vigília.Ações dos mineralocorticóidesResumidamente a aldosterona exerce três ações sobre a porção terminal do túbulo distal e os ductos coletores do rim: aumenta a reabsorção de Na+, a secreção de K+ e a secreção de H+. Assim, quando os níveis de aldosterona estão elevados, a reabsorção de Na+, e a sereção de K+ e H+ estão aumentadas. Essas alterações no transporte resultam em expansão do volume do LEC e hipertensão, hipocalemia e alcalose metabólica.
Ações dos androgênos supra-renaisOs DHEA e androstenediona são convertidos em testosterona nos testículos. Nas mulheres, esses androgênicos são os principais, sendo responsáveis pelo desenvolvimento dos pelos pubianos e axilares e pela libido.Em síndrome adrenogenital, na qual ocorre síntese aumentada de androgênios supra-renais, os altos níveis de DHEA e de androstenediona levam à masculinização em mulheres, ao desenvolvimento precoce dos pelos pubianos e axilares e à supressão das gônadas nos homens e mulheres.

Eixo hipotálamo-hipófise-tireóideOs hormônios tireoidianos são sintetizados e secretados pelas células epiteliais da tireóide. Produzem efeito sobre praticamente todos os sistemas do corpo. Os dois hormônios tireoideianos ativos são a triiodotironina (T3) e a tetraiodotironina (T4). Embora T3 seja mais ativo que o T4, quase toda a liberação hormonal é de T4. Esse problema de secretar a forma menos ativa é resolvido nos tecidos-alvo, que convertem T4 em T3. O T3 reverso não possui atividade biologicamente.Síntese de hormônios tireoidianosSão sintetizados pelas células epiteliais foliculares da tireóide. As células têm membrana basal que faceia o sangue e uma membrana apical que faceia o lúmem folicular. O material no lúmem é o colóide, que é composto por hormônios tireoidianos recém-sintetizados, ligados à tireoglobulina. Quando a tireóide é estimulada, esse colóide é absorvido por endocitose pelas células foliculares.Os hormônios tireoidianos contém grande quantidade de iodo, que deve ser adequadamente suprido na alimentação, os hormônios são armazenados extracelularmente no lúmen folicular, até que a tireóide seja estimulada a secretar e embora o T4 seja o principal produto secretado, não é a forma mais ativa.1. A tireoglobulina (glicoproteína) contém grandes quantidade de tirosina, é sintetizada no retículo endoplasmático rugoso e no aparelho de golgi das células foliculares. A seguir é incorporada por vesículas secretoras e liberada para o lúmen folicular. Depois as tirosinas da tireoglobulina serão iodadas, formando os precursores dos hormônios tireoidianos.2. Ocorre o co-transporte de Na+ e I-. Esse transporte ocorre ativamente do sangue para as células foliculares contra o gradiente eletroquímico. Essa bomba é regurada pelos níveis de I- no organismo. Quando há deficiência alimentar de iodo, esse transporte aumenta sua atividade, tentando compensar deficiência. Se a deficiência for grave, não haverá compensação e a síntese dos hormônios tireoidianos diminuirá.3. Quando o I- é bombeado para a célula, ele atravessa em direção à membrana apical, onde é oxidado a I2 pela enzima peroxidase tireóidea. Essa enzima catalisa essa etapa de oxidação, organização do I2 na tireoglobulina e na reações de acoplamento. 4. Na membrana apical, junto ao lúmen do folículo o I2 se combina com as tirosinas da tireoglobulina formando monoiodotirosina (MIT) e diiodotirosina (DIT). 5. Reação de acoplamento. Pode ocorrer acoplamento entre MIT e DIT formando (T3) ou duas DIT formando (T4).6. Quando a tireóide é estimulada a tireoglobulina iodada é endocitosada pelas células epiteliais foliculares. A membrana apical emite pseudópodos que engolfam uma porção do colóide e o absorvem para a célula. 7. As gotículas de tireoglobulina se fundem com membranas lisossômicas. Após isso hidrolisam as ligações peptídicas, liberando T4 e T3, que são transportados para os capilares, para serem levados à circulação sistêmica. Os MIT e DIT que permaneceram são reciclados na síntese de nova tireoglobulina.8. Desiodação de MIT e DIT. São desiodadas na célula folicular. O I- gerado é reciclado para o reservatório intracelular e adicionado ao I- transportado pela bomba. Já as moléculas de tirosina são incorporadas à síntese da nova tireoglobulina.
Os hormônios tireoidianos circulam na corrente sanguínea ligados a proteínas plasmáticas ou em forma livre. Como somente os hormônios tireoidianos livres são fisiologicamente ativos, o papel da globulina ligante de tiroxina (TGB) é formar e manter grande reservatório de hormônios tireoidianos circulantes, que podem ser liberados e adicionados ao hormônio livre.Ativação do T4 nos tecidos-alvo: através da enzima 5’-iodinase converte o T4 em T3 ou T3 reverso (inativo).

Regulação da secreção de hormônios tireoidianosO principal controle da síntese e secreção dos hormônios é pelo eixo hipotalâmico-hipofisário. O hormônio liberador de tireotropina (TRH) é secretado pelo hipotálamo e atua nas células tireotróficas da adeno-hipófise, provocando secreção do hormônio estimulante da tireóide (TSH). Esse atua na tireóide, aumentando a síntese e secreção dos hormônios tireoidianos.TRH: é secretado pelo hipotálamo que atua estimulando a transcrição do gene de TSH e a secreão desse hormônio (pode estimular a secreção de prolactina também).TSH: é secretado pela adeno-hipófise em resposta ao TRH. Seu papel é regular o crescimento da tireóide e secreção dos hormônios tireoidianos.A secreção de TSH é regulada pelo TRH secretado pelo hipotálamo e pelos hormônios tireoidianos que inibem a secreção de TSH por diminuir receptores de TRH nas células tireotróficas, diminuindo a sensibilidade ao estímulo pelo TRH. Esse mecanismo é mediado pelo T3 livre.Na tireóide: o TSH se liga ao receptor de membrana, acoplado à adenilil ciclase por meio da proteína G. Esta gera AMPc, que funciona como segundo mensageiro. Isso permite que haja o aumento da síntese e secreção de hormônios tireoidianos e quando TSH está elevado por período prolongado há hipertrofia e hiperplasia das células foliculares e fluxo tireoidiano aumentado.Ações dos hormônios tireoidianosEsses hormônios atuam sinergicamente com o hormônio do crescimento e somatomedinas, promovendo a formação de osso; aumentam a taxa de metabolismo basal, a produção de calor e o consumo de oxigênio; e alteram os sistemas cardiovascular e respiratório, aumentando o fluxo sanguíneo e a oferta de oxigênio aos tecidos.Após a produção de T3 nas células-alvo, este penetra ao núcleo e se liga ao receptor nuclear, estimulando a transcrição de DNA. Os RNAms recém transcritos são traduzidos e novas proteínas são sintetizadas. Muitas proteínas são sintetizadas sob o comando dos hormônios tireoidianos, como a NA+ K+ ATPase, proteínas de transporte, receptores adrenérgicos, enzimas lisossômicas, enzimas proteolíticas e proteínas estruturais. A enzima produzida dependerá do tecido-alvo.Taxa de metabolismo basal (TMB): aumento do consumo de oxigênio e o aumento resultante da TMB e da temperatura corporal. Esses hormônios aumentam o consumo de oxigênio em todos os tecidos, exceto no cérebro, nas gônadas e no baço. O hormônio estimula a síntese e o aumento da ativdade da Na+ e K+ ATPase. Isso explica a grande porcentagem do consumo total de oxigênio e a produção de calor no corpo.Metabolismo: o consumo aumentado de oxigênio depende da disponibilidade aumentada de substratos para o metabolismo oxidativo. Os hormônios aumentam a absorção da glicose pelo trato gastrointestinal e potencializam os efeitos de outros hormônios (catecolaminas, glucagon e hormônio do crescimento), aumentam a síntese e degradação de proteínas com efeito catabólico. Cardiovascular e respiratório: aumentado o consumo de O2, geram maior demanda de oxigênio pelos tecidos. Isso é possível poque há um aumento do débito cardíaco e da ventilação. O aumento da contratilidade é causada pelo aumento da síntese de receptores adrenérgicos, tornando o coração mais sensível ao estímulo pelo sistema nervoso simpático.Crescimento: atuam sinergicamente com o hormônio do crescimento e com as somatomedinas, promovendo a formação do osso. Juntos, promovem a ossificação e fusão das placas ósseas e a maturação do osso.Sistema nervoso central (SNC): no período perinatal, esse hormônio é essencial para a maturação normal do SNC, por isso é necessário uma triagem e se dectado o hipotireoidismo deve haver reposição hormonal para que haja reversão dos efeitos no SNC. Em adultos, pode causar desatenção, movimentos lentificados, sonolência, memória prejudicada e capacidade mental diminuída. O hipertireoidismo causa hiperexcitabilidade, hiper-reflexia e irritabilidade.Sistema nervoso autônomo: sabe-se que há interação entre os hormônios tireoidianos e o

sistema nervoso simpático. O significado dessa sinergia pode ser pela eficácia de bloqueadores adrenérgicos no tratamento dos sintomas do hipertireoidismo.HipertiroidismoA forma mais comum dessa doença é auto-imune (aumento de anticorpos contra receptores de TSH). Quando isso acontece ocorre uma estimulação intensa da tireóide, resultando na secreção aumentada de seus hormônios e hipertrofia da glândula. Outras causas são neoplasias da tireóide, produção excessiva de TRH ou TSH e administração excessiva de hormônios tireoidianos exógenos. O diagnóstico se baseia nos sintomas e determinação dos níveis aumentados de T3 e T4. Os níveis de TSH e TRH podem estar aumentados ou diminuídos dependendo da causa do hipertiroidismo. Os sintomas são perda de peso acompanhada de aumento de ingestão de alimentar (alta taxa metabólica), excessiva produção de calor e sudorese (aumento do consumo de oxigênio), alta frequência cardíaca (↑ receptores adrenérgicos), respiração ofegante ao esforço, tremor, nervosismo e fraqueza. Pode causar o bócio (atividade aumentada causa o crescimento da tireóide). O tratamento é feito pela administração de fármacos que inbem a síntese dos hormônios, remoção cirúrgica da glândula ou ablação radioativa da tireóide. HipotireoidismoA causa mais comum é a destruição auto-imune da tireóide, remoção cirúrgica da tireóide, insuficiência hipotalâmica ou hipofisária, deficiência de I-, raramente resistência do tecido-alvo. O diagnóstico se baseia nos sintomas e nos níveis diminuídos de T3 e T4. Os sitomas incluem taxa metabólica diminuída, ganho de peso sem ingestão alimentar, diminuição da produção de calor e intolerância ao frio, frequência cardíaca diminuída, lentidão dos movimentos, atividade mental diminuída, sonolência, perda de cabelo e disfunção menstrual. Quando a causa é defeito na tireóide há formação de bócio. Durante o período perinatal pode causar forma irreversível de retardo mental e de crescimento. O tratamento envolve reposição pelos hormônios tireoidianos, semelhante ao hormônio endógeno.